
Prevalence of blaKPC-2, blaKPC-3 and blaKPC-30—Carrying Plasmids in Klebsiella pneumoniae Isolated in a Brazilian Hospital

 , ,
, ,  and
and
Abstract
:1. Introduction
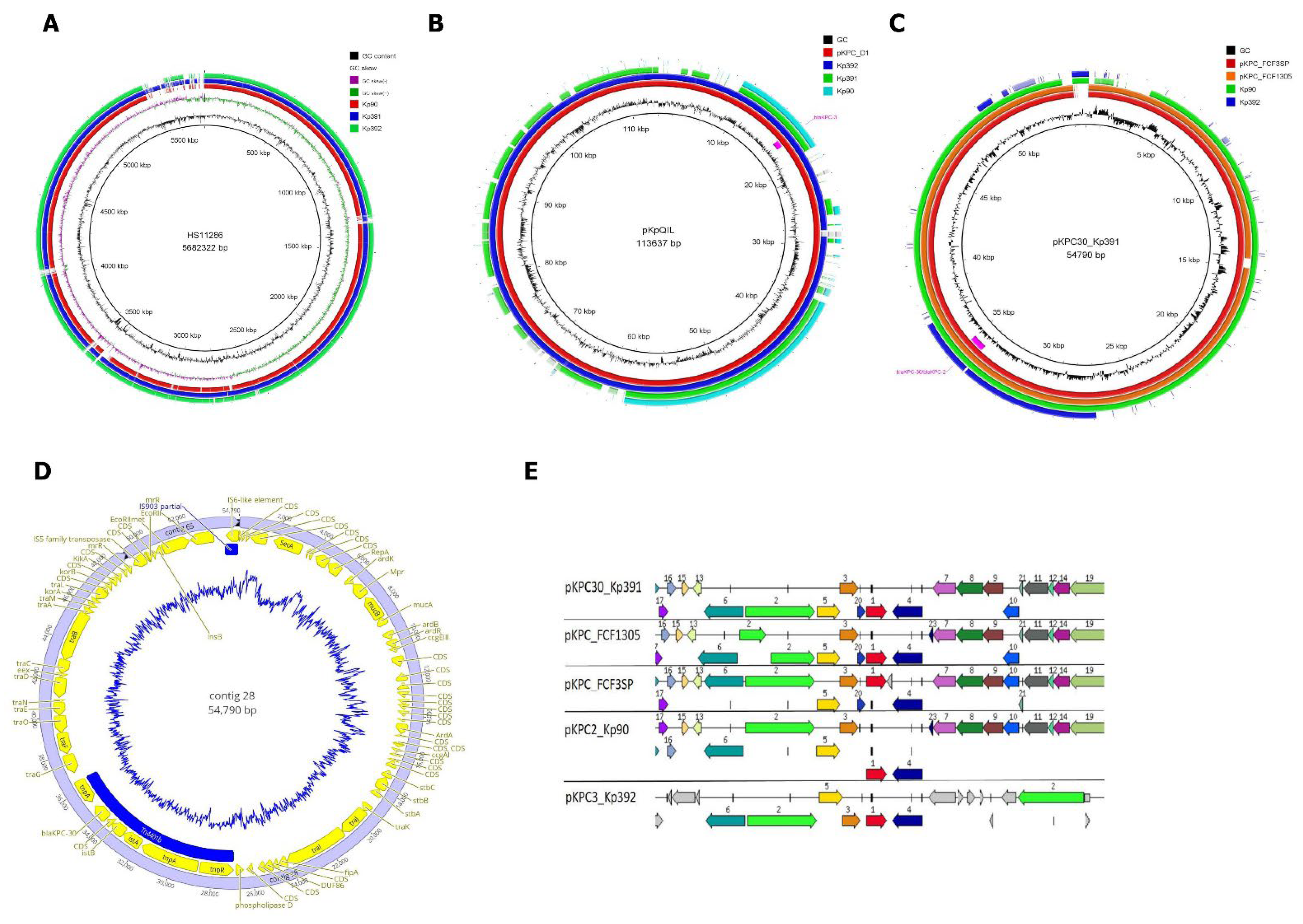
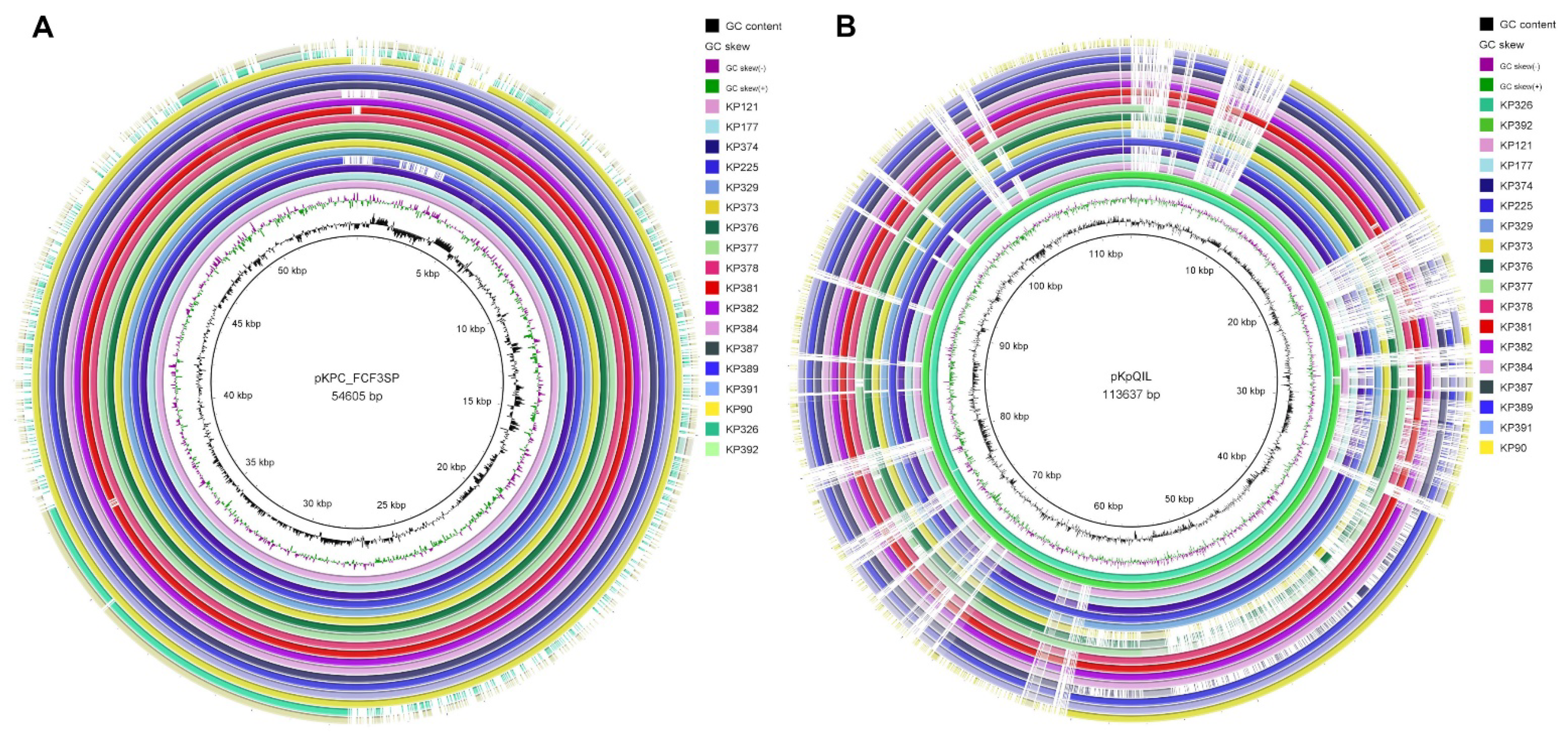
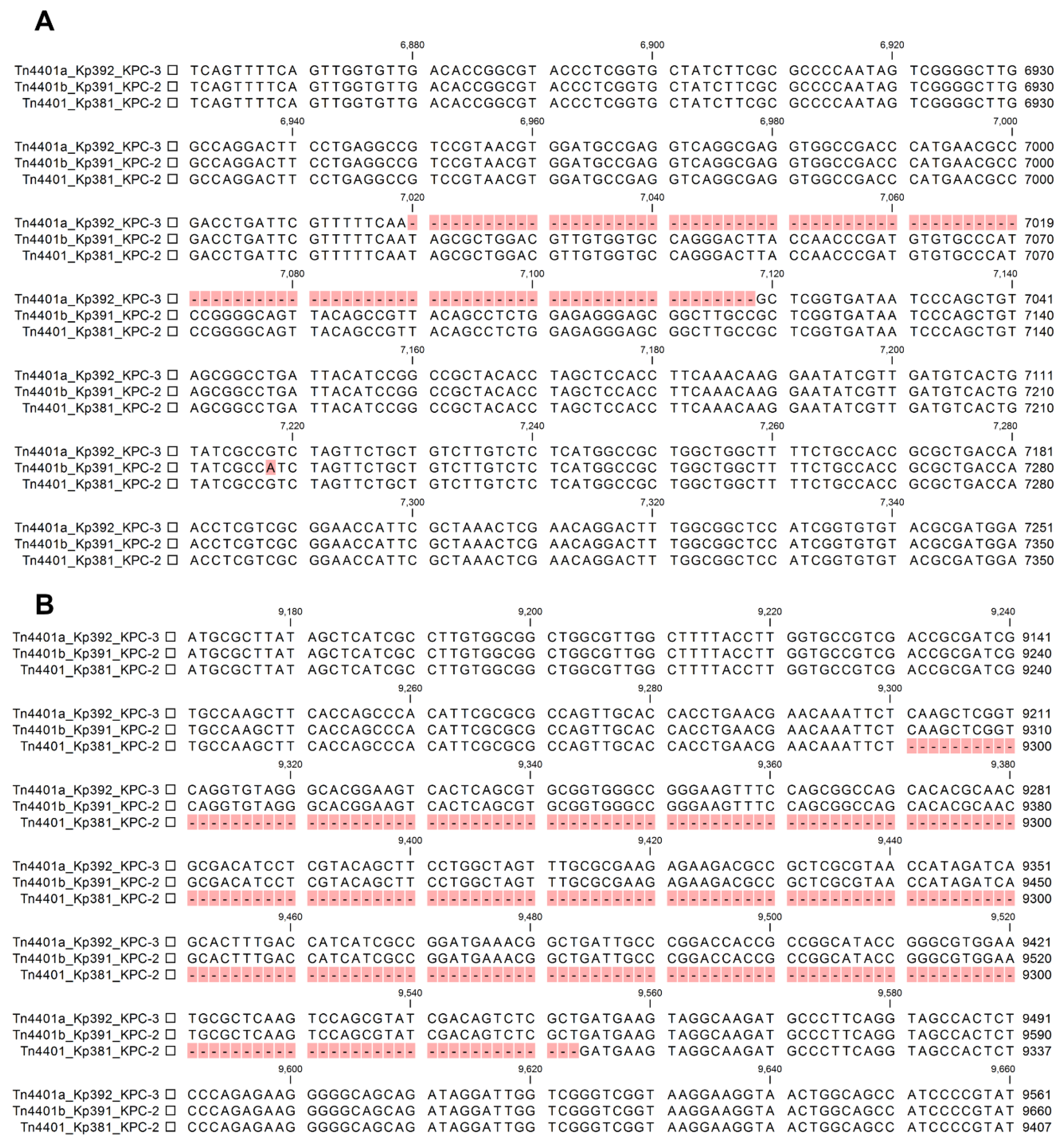
2. Results
3. Discussion
4. Materials and Methods
4.1. Clinical Isolates
4.2. Antibiotic Susceptibility Testing
4.3. Genome Sequencing
4.4. MLST and Capsule Synthesis Loci (K-loci) Analysis
4.5. NCBI Database Data Extraction for K. pneumoniae KPC-Harboring Plasmids
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Durante-Mangoni, E.; Andini, R.; Zampino, R. Management of carbapenem-resistant Enterobacteriaceae infections. Clin. Microbiol. Infect. 2019, 25, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Tooke, C.L.; Hinchliffe, P.; Bragginton, E.C.; Colenso, C.K.; Hirvonen, V.H.A.; Takebayashi, Y.; Spencer, J. beta-Lactamases and beta-Lactamase Inhibitors in the 21st Century. J. Mol. Biol. 2019, 431, 3472–3500. [Google Scholar] [CrossRef]
- Ambler, R.P.; Coulson, A.F.; Frère, J.M.; Ghuysen, J.M.; Joris, B.; Forsman, M.; Levesque, R.C.; Tiraby, G.; Waley, S.G. A standard numbering scheme for the class A beta-lactamases. Biochem. J. 1991, 276 Pt 1, 269–270. [Google Scholar] [CrossRef]
- Queenan, A.M.; Bush, K. Carbapenemases: The Versatile β-Lactamases. Clin. Microbiol. Rev. 2007, 20, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordmann, P.; Poirel, L. The difficult-to-control spread of carbapenemase producers among Enterobacteriaceae worldwide. Clin. Microbiol. Infect. 2014, 20, 821–830. [Google Scholar] [CrossRef] [Green Version]
- Logan, L.K.; Weinstein, R.A. The Epidemiology of Carbapenem-Resistant Enterobacteriaceae: The Impact and Evolution of a Global Menace. J. Infect. Dis. 2017, 215, S28–S36. [Google Scholar] [CrossRef] [Green Version]
- Maya, J.J.; Ruiz, S.J.; Blanco, V.M.; Gotuzzo, E.; Guzman-Blanco, M.; Labarca, J.; Salles, M.; Quinn, J.P.; Villegas, M.V. Current status of carbapenemases in Latin America. Expert Rev. Anti-Infect. Ther. 2013, 11, 657–667. [Google Scholar] [CrossRef]
- Monteiro, J.; Santos, A.F.; Asensi, M.D.; Peirano, G.; Gales, A.C. First report of KPC-2-producing Klebsiella pneumoniae strains in Brazil. Antimicrob. Agents Chemother. 2009, 53, 333–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seki, L.M.; Pereira, P.S.; de Souza Mda, P.; Conceição Mde, S.; Marques, E.A.; Porto, C.O.; Colnago, E.M.; Alves Cde, F.; Gomes, D.; Assef, A.P.; et al. Molecular epidemiology of KPC-2- producing Klebsiella pneumoniae isolates in Brazil: The predominance of sequence type 437. Diagn. Microbiol. Infect. Dis. 2011, 70, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, J.L.M.; Gales, A.C. Antimicrobial resistance in Enterobacteriaceae in Brazil: Focus on ²-lactams and polymyxins. Braz. J. Microbiol. 2016, 47, 31–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, P.C.S.; Monteiro, A.S.; Marques, S.G.; Monteiro, S.G.; Monteiro-Neto, V.; Coqueiro, M.M.M.; Marques, A.C.G.; de Jesus Gomes Turri, R.; Santos, S.G.; Bomfim, M.R.Q. Phenotypic and molecular detection of the blaKPC gene in clinical isolates from inpatients at hospitals in São Luis, MA, Brazil. BMC Infect. Dis. 2016, 16, 737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodford, N.; Tierno, P.M., Jr.; Young, K.; Tysall, L.; Palepou, M.-F.I.; Ward, E.; Painter, R.E.; Suber, D.F.; Shungu, D.; Silver, L.L.; et al. Outbreak of Klebsiella pneumoniae producing a new carbapenem-hydrolyzing class A beta-lactamase, KPC-3, in a New York Medical Center. Antimicrob. Agents Chemother. 2004, 48, 4793–4799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitchel, B.; Rasheed, J.K.; Patel, J.B.; Srinivasan, A.; Navon-Venezia, S.; Carmeli, Y.; Brolund, A.; Giske, C.G. Molecular epidemiology of KPC-producing Klebsiella pneumoniae isolates in the United States: Clonal expansion of multilocus sequence type 258. Antimicrob. Agents Chemother. 2009, 53, 3365–3370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, S.C.; Rice, K.; Palzkill, T. Natural Variants of the KPC-2 Carbapenemase have Evolved Increased Catalytic Efficiency for Ceftazidime Hydrolysis at the Cost of Enzyme Stability. PLoS Pathog. 2015, 11, e1004949. [Google Scholar] [CrossRef] [PubMed]
- Manageiro, V.; Ferreira, E.; Almeida, J.; Barbosa, S.; Simões, C.; Bonomo, R.A.; Caniça, M. Predominance of KPC-3 in a survey for carbapenemase-producing Enterobacteriaceae in Portugal. Antimicrob. Agents Chemother. 2015, 59, 3588–3592. [Google Scholar] [CrossRef] [Green Version]
- Campos, A.C.; Albiero, J.; Ecker, A.B.; Kuroda, C.M.; Meirelles, L.E.; Polato, A.; Tognim, M.C.; Wingeter, M.A.; Teixeira, J.J. Outbreak of Klebsiella pneumoniae carbapenemase-producing K pneumoniae: A systematic review. Am. J. Infect. Control 2016, 44, 1374–1380. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Antibiotic Resistance Threats in the United States. 2019. Available online: www.cdc.gov/drugresistance/pdf/threats-report/2019-ar-threats-report-508.pdf (accessed on 20 November 2020).
- Tzouvelekis, L.S.; Markogiannakis, A.; Psichogiou, M.; Tassios, P.T.; Daikos, G.L. Carbapenemases in Klebsiella pneumoniae and other Enterobacteriaceae: An evolving crisis of global dimensions. Clin. Microbiol. Rev. 2012, 25, 682–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, C.; Viehweger, A.; Singh, A.; Pletz, M.W.; Wibberg, D.; Kalinowski, J.; Lerch, S.; Müller, B.; Makarewicz, O. Assessing genetic diversity and similarity of 435 KPC-carrying plasmids. Sci. Rep. 2019, 9, 11223. [Google Scholar] [CrossRef] [PubMed]
- Villegas, M.V.; Blanco, M.G.; Sifuentes-Osornio, J.; Rossi, F. Increasing prevalence of extended-spectrum-beta-lactamase among Gram-negative bacilli in Latin America–2008 update from the Study for Monitoring Antimicrobial Resistance Trends (SMART). Braz. J. Infect. Dis. 2011, 15, 34–39. [Google Scholar] [PubMed]
- Beirão, E.M.; Rodrigues, S.D.S.; Andrade, T.K.; Serra, F.B.; Paula, M.D.N.; Polis, T.J.B.; Gales, A.C. Activity of ceftolozane-tazobactam and comparators against gram-negative bacilli: Results from the study for monitoring antimicrobial resistance trends (SMART–Brazil; 2016–2017). Braz. J. Infect. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Gorrie, C.; Edwards, D.J.; Wertheim, H.F.L.; Hsu, L.Y.; Van Kinh, N.; Zadoks, R.; Baker, S.; Holt, K.E. Extensive Capsule Locus Variation and Large-Scale Genomic Recombination within the Klebsiella pneumoniae Clonal Group 258. Genome Biol. Evol. 2015, 7, 1267–1279. [Google Scholar] [CrossRef] [Green Version]
- Qu, D.; Shen, Y.; Hu, L.; Jiang, X.; Yin, Z.; Gao, B.; Zhao, Y.; Yang, W.; Yang, H.; Han, J.; et al. Comparative analysis of KPC-2-encoding chimera plasmids with multi-replicon IncR:Inc(pA1763-KPC):IncN1 or IncFII(pHN7A8):Inc(pA1763-KPC):IncN1. Infect. Drug Resist. 2019, 12, 285–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, C.; Bavlovič, J.; Machado, E.; Amorim, J.; Peixe, L.; Novais, Â. KPC-3-Producing Klebsiella pneumoniae in Portugal Linked to Previously Circulating Non-CG258 Lineages and Uncommon Genetic Platforms (Tn4401d-IncFIA and Tn4401d-IncN). Front. Microbiol. 2016, 7, 1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leavitt, A.; Chmelnitsky, I.; Carmeli, Y.; Navon-Venezia, S. Complete nucleotide sequence of KPC-3-encoding plasmid pKpQIL in the epidemic Klebsiella pneumoniae sequence type 258. Antimicrob. Agents Chemother. 2010, 54, 4493–4496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jousset, A.B.; Bonnin, R.A.; Takissian, J.; Girlich, D.; Mihaila, L.; Cabanel, N.; Dortet, L.; Glaser, P.; Naas, T. Concomitant carriage of KPC-producing and non-KPC-producing Klebsiella pneumoniae ST512 within a single patient. J. Antimicrob. Chemother. 2020, 75, 2087–2092. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Hu, L.; Zhong, Q.; Hang, Y.; Liu, Y.; Hu, X.; Ding, H.; Chen, Y.; Xu, X.; Fang, X.; et al. Dissemination of Klebsiella pneumoniae ST11 isolates with carbapenem resistance in integrated and emergency intensive care units in a Chinese tertiary hospital. J. Med. Microbiol. 2019, 68, 882–889. [Google Scholar] [CrossRef]
- Chen, C.M.; Guo, M.K.; Ke, S.C.; Lin, Y.P.; Li, C.R.; Vy Nguyen, H.T.; Wu, L.T. Emergence and nosocomial spread of ST11 carbapenem-resistant Klebsiella pneumoniae co-producing OXA-48 and KPC-2 in a regional hospital in Taiwan. J. Med. Microbiol. 2018, 67, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yu, J.; Chen, F.; Yu, J.; Simner, P.; Tamma, P.; Liu, Y.; Shen, L. Emergence and establishment of KPC-2-producing ST11 Klebsiella pneumoniae in a general hospital in Shanghai, China. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 293–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Zhang, X.; Torres, V.V.L.; Liu, H.; Rocker, A.; Zhang, Y.; Wang, J.; Chen, L.; Bi, W.; Lin, J.; et al. An Outbreak of Carbapenem-Resistant and Hypervirulent Klebsiella pneumoniae in an Intensive Care Unit of a Major Teaching Hospital in Wenzhou, China. Front. Public Health 2019, 7, 229. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Chaparro, P.J.; Cerdeira, L.T.; Queiroz, M.G.; de Lima, C.P.S.; Levy, C.E.; Pavez, M.; Lincopan, N.; Gonçalves, E.C.; Mamizuka, E.M.; Sampaio, J.L.M.; et al. Complete nucleotide sequences of two blaKPC-2-bearing IncN Plasmids isolated from sequence type 442 Klebsiella pneumoniae clinical strains four years apart. Antimicrob. Agents Chemother. 2014, 58, 2958–2960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrey, D.O.; Pereira Dantas, P.; Martins, W.B.S.; Marques De Carvalho, F.; Almeida, L.G.P.; Sands, K.; Portal, E.; Sauser, J.; Cayô, R.; Nicolas, M.F.; et al. An Emerging Clone, Klebsiellapneumoniae Carbapenemase 2-Producing K. pneumoniae Sequence Type 16, Associated With High Mortality Rates in a CC258-Endemic Setting. Clin. Infect. Dis. 2020, 71, e141–e150. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Hu, X.; Zhao, Y.; Shi, Y.; Ding, H.; Wu, R.; Zhao, Z.; Ji, J. Comparative Analysis of bla (KPC) Expression in Tn4401 Transposons and the Tn3-Tn4401 Chimera. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [Green Version]
- Gootz, T.D.; Lescoe, M.K.; Dib-Hajj, F.; Dougherty, B.A.; He, W.; Della-Latta, P.; Huard, R.C. Genetic organization of transposase regions surrounding blaKPC carbapenemase genes on plasmids from Klebsiella strains isolated in a New York City hospital. Antimicrob. Agents Chemother. 2009, 53, 1998–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, W.W.; Stone, G.G.; Newell, P.; Broadhurst, H.; Wardman, A.; MacPherson, M.; Yates, K.; Riccobene, T.; Critchley, I.A.; Das, S. Ceftazidime-Avibactam Susceptibility Breakpoints against Enterobacteriaceae and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- The European Committee on Antimicrobial Susceptibility Testing (EUCAST). Breakpoint Tables for Interpretation of MICs and Zone Diameters. 2020. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_10.0_Breakpoint_Tables.pdf (accessed on 30 April 2020).
- Walsh, F. The multiple roles of antibiotics and antibiotic resistance in nature. Front. Microbiol. 2013, 4, 255. [Google Scholar] [CrossRef] [Green Version]
- Alba, J.; Ishii, Y.; Thomson, K.; Moland, E.S.; Yamaguchi, K. Kinetics study of KPC-3, a plasmid-encoded class A carbapenem-hydrolyzing beta-lactamase. Antimicrob. Agents Chemother. 2005, 49, 4760–4762. [Google Scholar] [CrossRef] [Green Version]
- Stillwell, T.; Green, M.; Barbadora, K.; Ferrelli, J.G.; Roberts, T.L.; Weissman, S.J.; Nowalk, A. Outbreak of KPC-3 Producing Carbapenem-Resistant Klebsiella pneumoniae in a US Pediatric Hospital. J. Pediatric Infect. Dis. Soc. 2015, 4, 330–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shields, R.K.; Clancy, C.J.; Hao, B.; Chen, L.; Press, E.G.; Iovine, N.M.; Kreiswirth, B.N.; Nguyen, M.H. Effects of Klebsiella pneumoniae carbapenemase subtypes, extended-spectrum β-lactamases, and porin mutations on the in vitro activity of ceftazidime-avibactam against carbapenem-resistant K. pneumoniae. Antimicrob. Agents Chemother. 2015, 59, 5793–5797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navon-Venezia, S.; Leavitt, A.; Schwaber, M.J.; Rasheed, J.K.; Srinivasan, A.; Patel, J.B.; Carmeli, Y.; Israeli, K.P.C.K.S.G. First report on a hyperepidemic clone of KPC-3-producing Klebsiella pneumoniae in Israel genetically related to a strain causing outbreaks in the United States. Antimicrob. Agents Chemother. 2009, 53, 818–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leavitt, A.; Chmelnitsky, I.; Ofek, I.; Carmeli, Y.; Navon-Venezia, S. Plasmid pKpQIL encoding KPC-3 and TEM-1 confers carbapenem resistance in an extremely drug-resistant epidemic Klebsiella pneumoniae strain. J. Antimicrob. Chemother. 2010, 65, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Doumith, M.; Findlay, J.; Hirani, H.; Hopkins, K.L.; Livermore, D.M.; Dodgson, A.; Woodford, N. Major role of pKpQIL-like plasmids in the early dissemination of KPC-type carbapenemases in the UK. J. Antimicrob. Chemother. 2017, 72, 2241–2248. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Chavda, K.D.; Melano, R.G.; Jacobs, M.R.; Koll, B.; Hong, T.; Rojtman, A.D.; Levi, M.H.; Bonomo, R.A.; Kreiswirth, B.N. Comparative genomic analysis of KPC-encoding pKpQIL-like plasmids and their distribution in New Jersey and New York Hospitals. Antimicrob. Agents Chemother. 2014, 58, 2871–2877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Dong, N.; Chan, E.W.-C.; Zhang, R.; Chen, S. Carbapenem Resistance-Encoding and Virulence-Encoding Conjugative Plasmids in Klebsiella pneumoniae. Trends Microbiol. 2020. [Google Scholar] [CrossRef]
- Holt, K.E.; Wertheim, H.; Zadoks, R.N.; Baker, S.; Whitehouse, C.A.; Dance, D.; Jenney, A.; Connor, T.R.; Hsu, L.Y.; Severin, J.; et al. Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proc. Natl. Acad. Sci. USA 2015, 112, E3574–E3581. [Google Scholar] [CrossRef] [Green Version]
- O’Hara, J.A.; Hu, F.; Ahn, C.; Nelson, J.; Rivera, J.I.; Pasculle, A.W.; Doi, Y. Molecular epidemiology of KPC-producing Escherichia coli: Occurrence of ST131-fimH30 subclone harboring pKpQIL-like IncFIIk plasmid. Antimicrob. Agents Chemother. 2014, 58, 4234–4237. [Google Scholar] [CrossRef] [Green Version]
- Papagiannitsis, C.C.a. Characterization of KPC-encoding plasmids from two endemic settings, Greece and Italy. J. Antimicrob. Chemother. 2016, 71, 2824–2830. [Google Scholar] [CrossRef] [PubMed]
- Eilertson, B.; Chen, L.; Chavda, K.D.; Kreiswirth, B.N. Genomic Characterization of Two KPC-Producing Klebsiella Isolates Collected in 1997 in New York City. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade, L.N.; Curiao, T.; Ferreira, J.C.; Longo, J.M.; Clímaco, E.C.; Martinez, R.; Bellissimo-Rodrigues, F.; Basile-Filho, A.; Evaristo, M.A.; Del Peloso, P.F.; et al. Dissemination of blaKPC-2 by the spread of Klebsiella pneumoniae clonal complex 258 clones (ST258, ST11, ST437) and plasmids (IncFII, IncN, IncL/M) among Enterobacteriaceae species in Brazil. Antimicrob. Agents Chemother. 2011, 55, 3579–3583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes, J.A.; Melano, R.; Cárdenas, P.A.; Trueba, G. Mobile genetic elements associated with carbapenemase genes in South American Enterobacterales. Braz. J. Infect. Dis. 2020, 24, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Brazilian Committee on Antimicrobial Susceptibility Testing (BrCAST). Breakpoint Tables for Interpretation of MICs and Zone Diameters—Tabelas de Pontos de Corte PARA Interpretação de CIMs e Diametros de Halos; Brazilian Committee on Antimicrobial Susceptibility Testing (BrCAST): São Paulo, Brazil, 2020. [Google Scholar]
- Salvà-Serra, F.; Svensson-Stadler, L.; Busquets, A.; Jaén-Luchoro, D.; Karlsson, R.; Moore, E.R.B.; Gomila, M. A protocol for extraction and purification of high-quality and quantity bacterial DNA applicable for genome sequencing: A modified version of the Marmur procedure. Protoc. Exch. 2018. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- García-Alcalde, F.; Okonechnikov, K.; Carbonell, J.; Cruz, L.M.; Götz, S.; Tarazona, S.; Dopazo, J.; Meyer, T.F.; Conesa, A. Qualimap: Evaluating next-generation sequencing alignment data. Bioinformatics 2012, 28, 2678–2679. [Google Scholar] [CrossRef] [PubMed]
- Alikhan, N.-F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wick, R.R.; Heinz, E.; Holt, K.E.; Wyres, K.L. Kaptive Web: User-Friendly Capsule and Lipopolysaccharide Serotype Prediction for Klebsiella Genomes. J. Clin. Microbiol. 2018, 56, e00197-00118. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Isolate | Year of Isolation | Origin of Specimen | bla-KPC Variant | Mlst Type (ST) | Clonal Group (CG) | CPS Locus (KL) | Genome Size (kb) | G+C (%) | Coverage (Fold) | Contigs (n 1) | CDS (n) | Genbank Accession |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Kp90 | 2015 | Blood | bla-KPC-2 | ST437 | 258 | 36 | 5429 | 57.4 | 77 | 76 | 5496 | JACBOR000000000 |
| Kp391 | 2016 | Abdominal abscess | bla-KPC-30 | ST11 | 258 | 64 | 5871 | 56.9 | 77 | 67 | 5956 | JACBOQ000000000 |
| Kp392 | 2017 | Urine | bla-KPC-3 | ST512 | 258 | 107 | 5681 | 57.1 | 103 | 81 | 5787 | JACBOP000000000 |
| Kp121 | 2014 | Blood | bla-KPC-2 | ST437 | 258 | 36 | 5554 | 57.3 | 84 | 55 | 5601 | JAEVGO000000000 |
| Kp177 | 2013 | Blood | bla-KPC-2 | ST437 | 258 | 36 | 5588 | 57.3 | 83 | 79 | 5679 | JAEVGN000000000 |
| Kp225 | 2011 | Blood | bla-KPC-2 | ST437 | 258 | 36 | 5777 | 57.1 | 64 | 81 | 5847 | JAEVGM000000000 |
| Kp326 | 2016 | Blood | bla-KPC-2 | ST16 | - | 51 | 5568 | 57.2 | 91 | 73 | 5648 | JAEVGL000000000 |
| Kp329 | 2015 | Bronchoalveolar lavage | bla-KPC-2 | ST11 | 258 | 64 | 5887 | 57.1 | 66 | 127 | 6049 | JAEVGK000000000 |
| Kp384 | 2014 | Blood | bla-KPC-2 | ST437 | 258 | 36 | 5569 | 57.3 | 66 | 79 | 5672 | JAEVGJ000000000 |
| Kp387 | 2013 | Ascitic fluid | bla-KPC-2 | ST11 | 258 | 27 | 5852 | 57.0 | 51 | 105 | 5967 | JAEVGI000000000 |
| Kp389 | 2013 | Bronchoalveolar lavage | bla-KPC-2 | ST437 | 258 | 36 | 5473 | 57.3 | 88 | 75 | 5524 | JAEVGH000000000 |
| Kp373 | 2013 | Rectal surveillance swabs | bla-KPC-2 | ST11 | 258 | 64 | 5900 | 57.0 | 81 | 68 | 6021 | JAEVGG000000000 |
| Kp374 | 2013 | Rectal surveillance swabs | bla-KPC-2 | ST437 | 258 | 36 | 5604 | 57.3 | 84 | 78 | 5702 | JAEVGF000000000 |
| Kp376 | 2015 | Rectal surveillance swabs | bla-KPC-2 | ST11 | 258 | 27 | 6162 | 56.6 | 82 | 135 | 6385 | JAEVGE000000000 |
| Kp377 | 2015 | Rectal surveillance swabs | bla-KPC-2 | ST11 | 258 | 64 | 5808 | 56.8 | 91 | 115 | 5953 | JAEVGD000000000 |
| Kp378 | 2016 | Rectal surveillance swabs | bla-KPC-2 | ST11 | 258 | 64 | 5872 | 56.9 | 87 | 62 | 5965 | JAEVGC000000000 |
| Kp381 | 2017 | Rectal surveillance swabs | bla-KPC-2 | ST437 | 258 | 36 | 6095 | 56.5 | 78 | 128 | 6344 | JAEVGB000000000 |
| Kp382 | 2017 | Rectal surveillance swabs | bla-KPC-2 | ST11 | 258 | 64 | 5877 | 56.9 | 94 | 97 | 6023 | JAEVGA000000000 |
| Kp90 | Kp391 | Kp392 | ||||
|---|---|---|---|---|---|---|
| Antimicrobial | MIC µg/mL | Profile | MIC µg/mL | Profile | MIC µg/mL | Profile |
| Ampicillin/Sulbactam | ≥32 | R | ≥32 | R | ≥32 | R |
| Piperacilin/Tazobactam | ≥128 | R | ≥128 | R | ≥128 | R |
| Cefuroxime | ≥64 | R | ≥64 | R | ≥64 | R |
| Cefoxitin | ≥64 | R | ≥64 | R | ≥64 | R |
| Ceftazidime | 16 | R | ≥64 | R | ≥64 | R |
| Ceftriaxone | ≥64 | R | ≥64 | R | ≥64 | R |
| Cefepime | ≥64 | R | ≥64 | R | ≥64 | R |
| Ertapenem | ≥8 | R | ≥8 | R | ≥8 | R |
| Imipenem | ≥16 | R | ≥16 | R | ≥16 | R |
| Meropenem | ≥16 | R | ≥16 | R | ≥16 | R |
| Amikacin | ≤2 | S | ≤2 | S | ≥64 | R |
| Gentamicin | ≤1 | S | ≤1 | S | 8 | R |
| Ciprofloxacin | ≥4 | R | ≥4 | R | ≥4 | R |
| Tigecycline | ≥8 | - | ≤0.5 | - | 2 | - |
| Imipenem | >32 | R | >32 | R | 32 | R |
| Meropenem | >32 | R | >32 | R | >32 | R |
| Ceftazidime/avibactam | 2 | S | 2 | S | 6 | S |
| Colistin | ≥16 | R | ≥16 | R | ≥16 | R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Migliorini, L.B.; de Sales, R.O.; Koga, P.C.M.; Doi, A.M.; Poehlein, A.; Toniolo, A.R.; Menezes, F.G.; Martino, M.D.V.; Gales, A.C.; Brüggemann, H.; et al. Prevalence of blaKPC-2, blaKPC-3 and blaKPC-30—Carrying Plasmids in Klebsiella pneumoniae Isolated in a Brazilian Hospital. Pathogens 2021, 10, 332. https://doi.org/10.3390/pathogens10030332
Migliorini LB, de Sales RO, Koga PCM, Doi AM, Poehlein A, Toniolo AR, Menezes FG, Martino MDV, Gales AC, Brüggemann H, et al. Prevalence of blaKPC-2, blaKPC-3 and blaKPC-30—Carrying Plasmids in Klebsiella pneumoniae Isolated in a Brazilian Hospital. Pathogens. 2021; 10(3):332. https://doi.org/10.3390/pathogens10030332
Chicago/Turabian StyleMigliorini, Letícia B., Romário O. de Sales, Paula C. M. Koga, Andre M. Doi, Anja Poehlein, Alexandra R. Toniolo, Fernando G. Menezes, Marines D. V. Martino, Ana C. Gales, Holger Brüggemann, and et al. 2021. "Prevalence of blaKPC-2, blaKPC-3 and blaKPC-30—Carrying Plasmids in Klebsiella pneumoniae Isolated in a Brazilian Hospital" Pathogens 10, no. 3: 332. https://doi.org/10.3390/pathogens10030332
APA StyleMigliorini, L. B., de Sales, R. O., Koga, P. C. M., Doi, A. M., Poehlein, A., Toniolo, A. R., Menezes, F. G., Martino, M. D. V., Gales, A. C., Brüggemann, H., & Severino, P. (2021). Prevalence of blaKPC-2, blaKPC-3 and blaKPC-30—Carrying Plasmids in Klebsiella pneumoniae Isolated in a Brazilian Hospital. Pathogens, 10(3), 332. https://doi.org/10.3390/pathogens10030332