1. Introduction
Verticillium wilt of olive (VWO), caused by
Verticillium dahliae Kleb., is one of the major constraints for olive growing throughout the Mediterranean basin [
1,
2]. Particularly, the disease is severe in the Guadalquivir Valley in southern Spain (Andalusia), the world’s leading area for the production of olive oil and table olives (more than 1.5 million ha of olive orchards). In this area, the pathogen causes severe economic losses and kills thousands of trees annually. Thus, recent surveys on Verticillium wilt affected orchards in this region revealed mean disease incidence of 12%, 22%, and 24% in the three main olive producing provinces, Sevilla, Córdoba, and Jaén, respectively [
3].
Some of the major causes of the VWO prevalence are the use of infested soils due to the previous crop of susceptible hosts of the pathogen and/or the infestation of healthy soils with inoculum coming from several outside inoculum sources. There were reported a number of means of dispersal of the pathogen in olive growing areas, mainly related to the movement of infective and survival structures (microsclerotia) that
V. dahliae releases into the soil at the end of the disease cycle. Therefore, microsclerotia can be spread within or between olive orchards by the movement of infested soil or infected plant debris due to wind and rainfall or by the use of contaminated machinery [
4,
5,
6,
7,
8]. In addition, dispersal by cultural practices, such as the use of infested animal manure as amendments in olive plantations [
9], leaves and dried inflorescences from wilted trees [
10], and irrigation water [
11,
12] were demonstrated. Besides these dispersal means, also the use of infected planting material is considered as a major factor in pathogen dispersal [
13].
In soil, microsclerotia germinate and infect the roots of olive trees, initiating tissue colonization that leads the pathogen to the xylem vessels. There, the fungus produces mycelia and conidia that colonize the xylem. The infection process is favored by the moderate temperatures (20–25 °C) present in the conditions of southern Spain during spring and autumn months. If the density of the
V. dahliae in the soil and its virulence are high, the olive cultivar presents high susceptibility to the fungus, and the favorable period for the development of infections is long; an extensive and consistent colonization of the vascular tissues is achieved during these weeks. Consequently, the tracheomycosis of the plant occurs, which interrupts water transport, subsequently causing defoliating, chlorosis, wilting of stems and branches of the tree, and, eventually, tree death [
14,
15].
Phloeotribus scarabaeoides (Bernard), the olive bark beetle, is a small xylophagous insect (Coleoptera: Scolytidae) widely spread in the Mediterranean basin and considered a secondary pest in olive groves [
16]. The olive bark beetle attacks young olive branches in diseased, damaged, or weakened trees still attached to the stem or detached pruned branches as debris that are left in the field or collected and piled by farmers for fireplaces. In southern Spain, wintering adults have their refugee in early spring and look for olive branch debris from tree pruning or from declining olive trees. They perforate the bark and excavate a place for mating, which consists of a number of sub-cortical galleries where females deposit the eggs (mating galleries). After an incubation period from 8 to 13 days, hatched larvae excavate new galleries that are perpendicular to the maternal gallery. From April to the middle of May, depending on the weather, new adults come out from mating galleries, causing numerous and recognizable exit holes. These adults attack new plants producing feeding galleries, easily recognizable by the sawdust exuded from galleries and deposited around entrance holes. They remain in the feeding galleries after the end of summer (end of September or beginning of October), then adults leave them and excavate new small holes in olive wood or in the axils of the vegetative buds in which they overwinter, originating the attack the following spring [
16].
Several plant pathogenic fungal species can be efficiently transmitted by xylophagous insects [
17]. Nevertheless, only few cases are directly associated with vascular diseases. The most representative example of a vascular disease caused by a beetle-transmitted fungus is Dutch elm disease caused by
Ophiostoma ulmi (Buisman) Nannf., which is transmitted by
Scolytus spp. [
18]. Adults of
Scolytus spp. transport conidia of the pathogen stuck to their bodies when they leave the mating galleries drilled in diseased elm trees and go to healthy trees to make feeding galleries. This fungus–insect association is not a simple transmission but a mutualistic relationship between the beetle and the pathogen, which does not appear to occur in the case of
V. dahliae.
In the case of
Verticillium spp., there is only little information about its transmission by insects [
19,
20,
21,
22,
23,
24]. One of the examples is alfalfa, in which certain insects, such as pea aphid, alfalfa weevil, migratory grasshopper, two-striped grasshopper, and alfalfa leafcutting bee, can serve as vectors for effective transmission of the relative pathogen
V. albo-atrum [
2]. Interestingly, [
19,
21] demonstrated the transmission of
V. dahliae by xylophagous insects
Scolytus intricatus (Ratz.) and
Xyleborus dispar (F.) when studying the causes of oak decline in Italy. Oak, however, is a rare host of
V. dahliae. Moreover, recent studies conducted by [
24] demonstrate that
Eucryptorrhynchus brandti Harold (Coleoptera: Curculionidae) has the ability to carry and transfer
V. nonalfalfae to
Ailanthus altissima (tree of heaven) in a laboratory setting, initiating plant seedling infection after external contact with the fungus.
In this context, the vascular nature of infections of V. dahliae in olive and the life cycle of P. scarabaeoides could suggest an interaction between the two organisms regarding the spread of V. dahliae inoculum from Verticillium wilt affected olive trees to healthy trees. This hypothesis was supported by previous observations in fields doubly damaged by the olive bark beetle and VWO in several locations in Andalusia (southern Spain).
Therefore, the objective of the present study was to assess the possible role of P. scarabaeoides as a vector for transmission of V. dahliae in olive orchards. If it is so, it would explain why it is common to find old olive orchards (more than 60 years old) affected by VWO, when these orchards were established in pathogen free soil and apparently out of the influence of external contribution of V. dahliae inoculum by anyone of the dispersal means indicated above.
3. Discussion
This is the first study evaluating the role of
P. scarabaeoides, the olive bark beetle, as a potential vector of
V. dahliae in olive orchards. The initial hypothesis was based on the activities related to the biological cycle of this beetle (Coleoptera, Scolytidae) and the cycle of infection of
V. dahliae. The main means of dispersal of
V. dahliae take place through the movement of its survival structures, called microsclerotia, during the phase of the cycle in which the pathogen is not in contact with the plant. Nevertheless, once the microsclerotia infect the root of the plant, the pathogen causes the infection of the vascular tissues through the production of mycelium and conidia that progressively colonize the xylem, where it remains viable. It constitutes the basis of the initial hypothesis of this work according to which the adults of the olive bark beetles could acquire and later transmit the pathogen to other healthy plants during the biological coincidence of both (production of feeding or mating galleries of
P. scarabaeoides close or in the invaded xylem vessels by
V. dahliae) [
16]. We were unable to find any evidence that pointed to olive bark beetles as vectors of
V. dahliae following sampling of adult beetles from infected olive trees in several olive orchards affected by the two agents: the fungal pathogen and the olive bark beetle. This was despite samples of shoots and branches colonized by the beetles confirmed as having high disease incidence. The validity of this negative result was supported by the fact that sampling was conducted at the most favorable time (April and May) for disease development under our conditions [
25].
Despite this outcome, experiments were conducted over a long period (March 2009 to May 2012) in order to explore in several years all the possibilities of inoculation by the insect when its biological cycle coincided with the most favorable period for infection and VWO symptom development. These periods were mainly spring, early summer, and fall.
The experimental design, using mosquito nets, allowed the isolation of the treatments, where the movement of adult olive bark beetles between healthy and artificially wilted olive trees was guaranteed. Indeed, the quantification of entry holes caused by the insect in the shoots of the healthy trees included in the nets was a main parameter for demonstrating that the inoculation of
V. dahliae might have occurred if the insect was able to transmit the pathogen (
Table 1,
Table 2,
Table 3 and
Table 4). Moreover, the role of the olive bark beetle as a vector of
V. dahliae was assessed by evaluating disease symptoms of VWO and the presence of the pathogen in tissues of olive trees invaded by adult insects. Finally, adult beetles were also extracted from mating and feeding galleries excavated in shoots of olive trees artificially or naturally infected by
V. dahliae that showed severe VWO symptoms. Thereafter, the presence of the pathogen in superficial and internal tissues of these beetles was investigated.
The presence of feeding galleries in the shoots sampled during the period from September 2009 to March 2010 (Experiment II) shows that the insect was active and could move from one tree to another. However, the absence of disease symptoms and infections of trees or contaminated beetle adults showed that the inoculation did not occur.
A third experiment was performed in which the olive trees used in treatments T3 and T4 were inoculated twice by stem injection with
V. dahliae. The aim was to uniformly infect the tissues in which
P. scarabaeoides was actively producing mating and feeding galleries and, thereafter, introduce in the nets new, healthy olive trees. The first inoculation did not produce consistent symptoms of VWO. This was likely due to high temperatures registered at the beginning of summer, which impeded an extensive colonization of tree tissues by the pathogen. Furthermore, the stem injection inoculation of olive plants generally results in colonization not as intense as when using root inoculation in controlled conditions [
26,
27]. Nevertheless, the second inoculation, conducted in October 2010, caused typical VWO symptoms in the olive trees, but the isolation on PDA plates from affected tissues confirmed the presence of the pathogen in only 4 out of the 13 symptomatic trees. This slow rate of isolations is sometimes common after artificial inoculations of woody host of
V. dahliae such as olive tree, since its slow growth makes it tough to isolate if any other fungi are present.
In this experiment, the number of feeding and mating galleries caused by beetles (
Table 3 and
Table 4) showed a similar trend in Experiment II regardless of the releasing insects. Although the number of feeding galleries was variable in the three treatments, we think that this number was high enough to transmit the pathogen if the olive bark beetle was an effective vector of the pathogen.
Healthy olive trees exposed to olive bark beetles in the described experimental conditions showed Verticillium wilt disease symptoms. Similarly, the pathogen could not be isolated from any of the tissues of sampled shoots where the insects made galleries or from any of the tissues of the beetle adults collected from the galleries.
Ascomycetous fungi associated with bark beetles include the genera
Ophiostoma,
Ceratocystis, and
Ceratocystiopsis and the anamorphic genera
Graphium and
Leptographium [
28,
29,
30]. Several fungal species causing different diseases in tree hosts can be efficiently transmitted by xylophagous insects [
17]. Nevertheless, only a few cases are directly associated with vascular diseases. The most representative example of a vascular disease caused by a beetle-transmitted fungus is Dutch elm disease caused by
Ophiostoma ulmi, which is transmitted by
Scolytus spp. [
18]. Diseases caused by
Ceratocystis include vascular wilts,
Ceratocystis fimbriata, Ellis & Halst., one of the most virulent and economically important vascular pathogens in many agricultural crops and forest trees [
31], which is frequently associated with insects on
Populus [
32] and
Prunus [
33].
We did not find scientific evidence in literature that expressly indicates that Verticillium dahliae is capable of surviving in the adults or the larval stages of the olive bark beetle. In fact, the starting hypothesis did not consider such survival per se but considered that the adults of the insect could constitute a support for the transfer of the conidia from one tree to another. The hypothesis was based on a possible parallelism with the mutualistic association that occurs in the pathosystem Ophiostoma novo ulmi/Bark beetles of Scolytus spp. Geoffroy, S. scolytus (F.) and S. multistriatus (Marsham), the main vectors of the pathogen which causes the Dutch elm disease.
Adaptations for insect dispersal (ascomata and conidiomata with sticky spore drops) arose frequently in the evolution of Ascomycetes [
34]. In this case, O. novo-ulmi presents during its life cycle two asexual forms that produce asexual spores (conidia). First, in infected trees (dying or recently dead), the mycelium of the fungus produces sticky conidia (Graphium-type spores) in the galleries created by beetles just under the bark. These spores are produced at the tips of synnemata (coremia), dark brown to black stalks about 1–2 mm in length, each composed of aggregated hyphae [
35,
36,
37].
When the new beetles emerge as adults from infected elms, they acquire the conidia of the fungus on and in their bodies and carry them to new elm trees, where they produce feeding tunnels in direct contact with the xylem vessels of the host. The pathogen invades the vascular tissues where they germinate to produce mycelium and cephalosporium-type spores (yeast phase) that move and colonize the vessels, causing tree wilt [
38,
39]. In the case of
V. dahliae, there is very little information about its transmission by insects. For example, the transmission of
Verticillium dahliae by xylophagous insect vectors
Scolytus intricatus (Ratz.) and
Xyleborus dispar (F.) was demonstrated when studying the causes of oak decline in Italy [
19,
21]. Oak tree, however, is a rare host of
V. dahliae. Nevertheless,
V. dahliae does not produces similar asexual structures such as the mentioned synnemata produced by
Ophiostoma spp that would cause the pathogen to contact the adult beetle bodies when they abandon the galleries looking for a new, healthy tree. The pathogen enters their host plants through the roots, and, following penetration, the fungi colonize the cortical cells from where hyphae migrate intercellularly toward the vascular parenchyma cells and invade the xylem vessels, where Yadeta and Thomma (2013) are mostly restricted. Therefore, the conidial production is limited to the vascular vessels, and the opportunity of beetle adults to get in contact with and to acquire conidia of the pathogen is restricted to this moment.
We can conclude that P. scarabaeoides, the olive bark beetle, is not a vector of V. dahliae, the pathogen causing Verticillium wilt of olive, in the investigated experimental conditions. The causes are probably related to the inefficiency of the beetle to acquire inoculum from diseased trees (attached to its body or inside its digestive tract) and/or transport and deposit viable pathogen conidia in the appropriate site, close to or inside xylem vessels, which would allow pathogens to establish the infection.
4. Materials and Methods
During the period May 2009 to April 2012, three experiments were conducted in commercial olive orchards and in semi-controlled conditions in a net-house at the Campus of Rabanales, University of Córdoba.
4.1. Experiment I
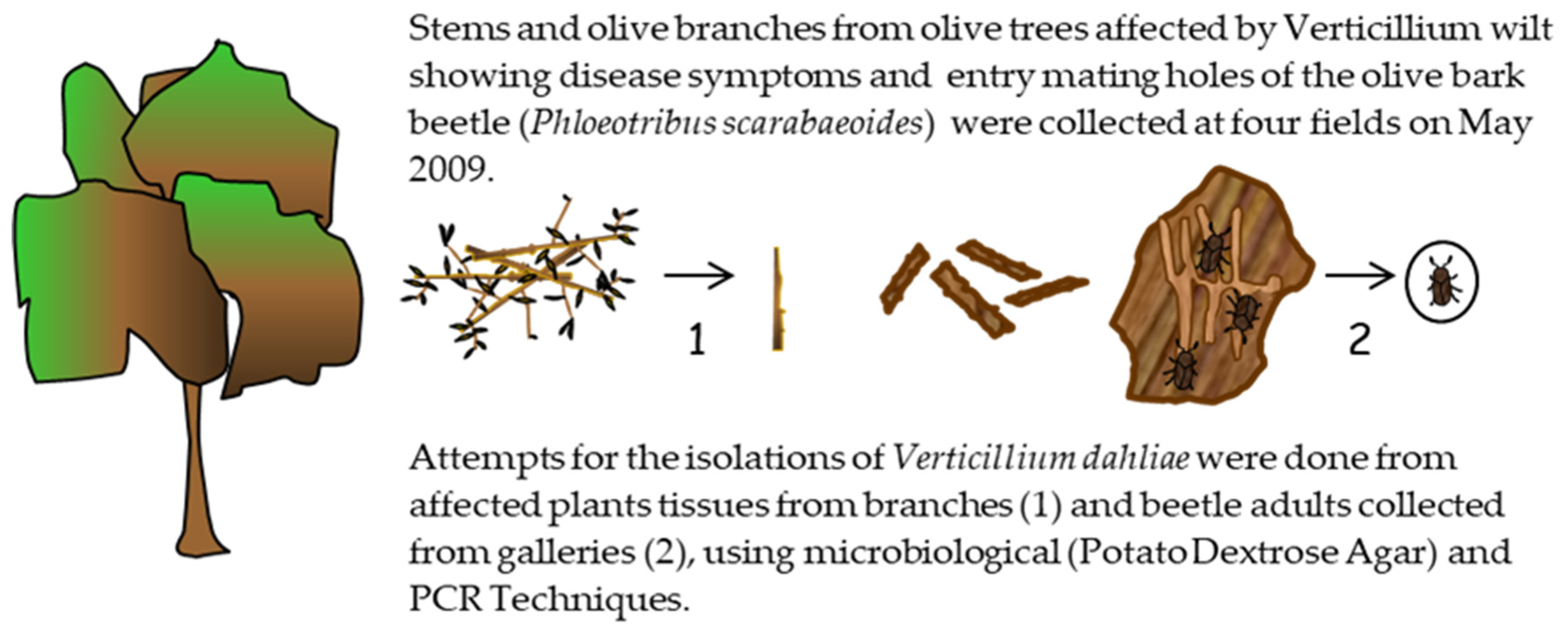
A field survey was conducted to collect olive branches showing wilt symptoms and/or entry holes (mating galleries) of
P. scarabaeoides in four olive orchards severely affected by VWO, located in the provinces of Córdoba (municipalities of Almodóvar, Montilla and Santaella) and Jaén (municipality of Andújar) (
Figure 1). The tree samples were used to check for the presence of the pathogen on the potential vector, and as a source of beetles for Experiments II and III. In addition, 3 to 6 shoots or branches (between 8 and 15 cm in diameter) per olive tree were collected from 12 olive trees from each orchard at two sampling dates during period April and May 2009.The presence of
V. dahliae in sampled branches was assessed by microbiological isolations. For these analyses, samples of affected woody tissue were washed in running tap water for 20 min, bark was removed, and tissue was surface disinfected in 0.5% sodium hypochlorite for 40 s. Wood chips were placed on potato dextrose agar (PDA) plates and incubated at 24 °C in the dark for 5 to 6 days. In addition, the detection of the pathogen in the samples was tested by molecular analyses based on nested-PCR [
34].
For collecting adults of P. scarabaeoides from the plant samples, branches were stored in big plastic boxes (38 cm length, 30 cm width, and 14 cm high) at room temperature in the laboratory. After emergence, adults were collected in small plastic boxes, and microbiological isolations of V. dahliae from inner and superficial tissues of them were done using two different methods.
(1) To check for the superficial presence of the fungal pathogen on the insect, 50 adults were chosen at random from those that emerged in the plastic boxes containing plant material from the different surveyed fields and placed onto plates (five adults per plate) of Malt Agar culture media amended with 1 mL of tetracycline (Sigma Aldrich) (0.05 g/mL), 1 mL of streptomycin (Sigma Aldrich) (0.60 g/mL), 5 mL of Ciclohexamida PB (Panreac) (0.05 g/mL), and 1 mL of dodine pestanal (Sigma Aldrich) (0.1 g/mL) [
35,
36].
(2) Another 50 adults were surface disinfected in 1.5% sodium hypochlorite for 40 s and then dried on sterile filter paper. Then, they were separately immersed in 5 mL of sterile distilled water and individually ground with a sterile mortar and pestle. The resulting mass per insect was diluted (0.1, 0.01, and 0.001), and 300 µL of each suspension was spread onto culture plates of the same medium. Plates were incubated at 22–24 °C for 7 days. Finally, the detection of the pathogen in the ground samples was also attempted using the nested-PCR technique [
34].
4.2. Experiment II
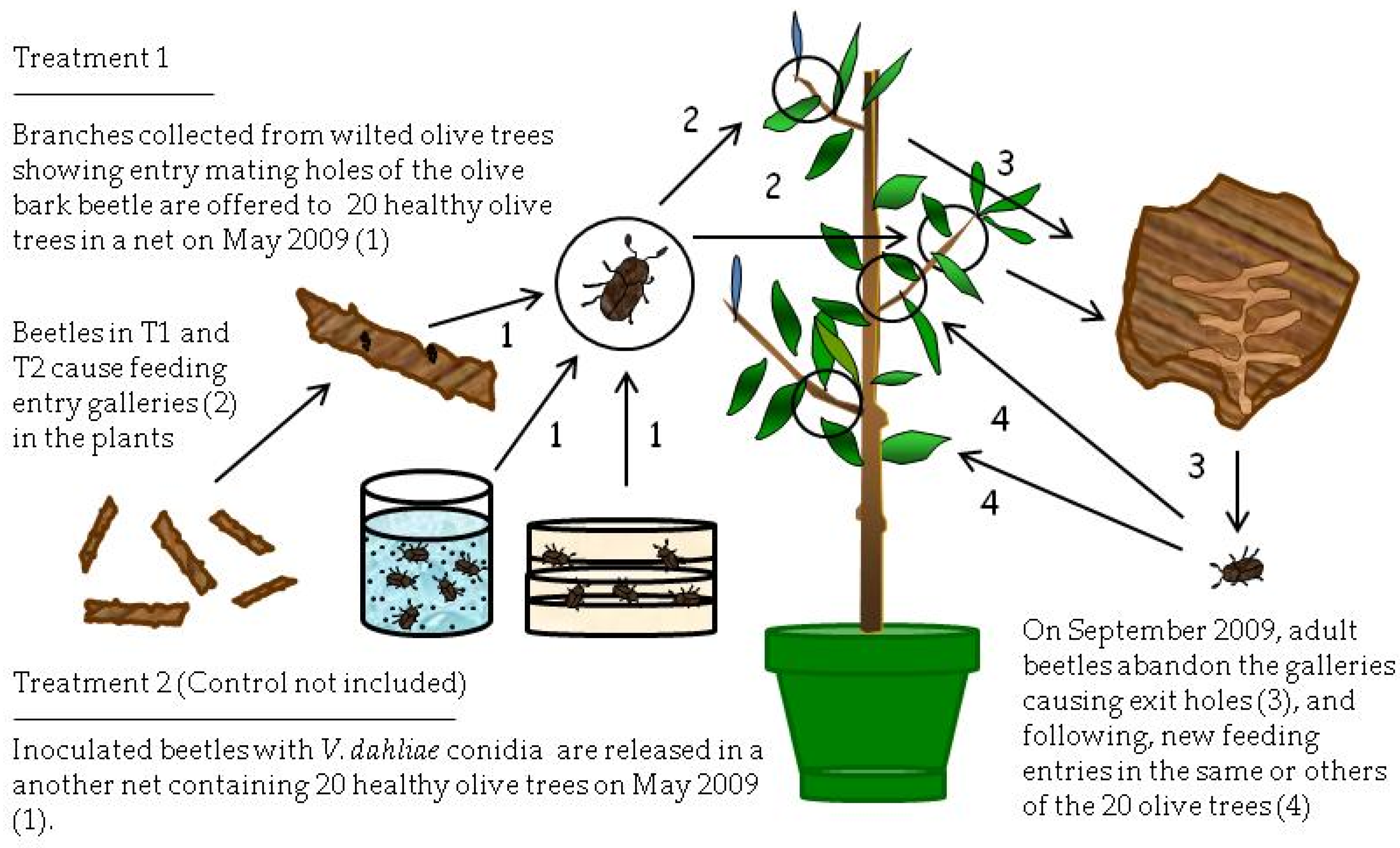
This experiment was carried out from May 2009 to March 2010 (
Figure 2). Healthy, 2.5-year-old olive trees (1.6–2.0 m height) of the Spanish cultivar Picual, susceptible to Verticillium wilt [
26], were used. These trees were acquired from a commercial nursery in Villanueva del Duque (north of Córdoba province, Spain). Nursery trees were propagated by rooting under mist of soft-wood cuttings obtained from certified disease-free olive mother trees. Trees grew in plastic pots (20 L in volume) in a lathhouse.
The experiment consisted of the following treatments. Treatment 1 (T1) was composed of four groups of four VWO-free olive trees. Each group of four trees was individually covered by a carbon fiber mosquito net and exposed early in May to seven branches affected by VWO that showed entry holes (mating galleries) by the bark beetle (
Figure 2). The hypothesis was that adults, developed into mating galleries (supposedly infested by conidia of
V. dahlia) after exiting the mating galleries from May to June, would produce entry holes (feeding galleries) in branches or main trunk and inoculate the healthy trees with
V. dahliae.
Treatment two (T2) also consisted of four groups of four similar olive trees covered by mosquito nets, but, in this case, trees were exposed to bark beetle adults previously infested by the pathogen that were released inside the nets (
Figure 2). The isolate of
V. dahliae coded as V117, a highly virulent cotton defoliating strain from the collection of the Plant Pathology Laboratory of the Agronomy Department at the University of Córdoba [
37], was used to inoculate the bark beetle adults. The pathogen was transferred from PDA slants, where it was maintained at 4 °C, to PDA plates by spreading the mycelium uniformly over the entire surface of each plate. Plates were incubated at 25 °C for 6 days in the dark. Infestation of beetles was achieved by confining adults for 7 h in a sandwich composed of two PDA plates with active cultures of
V. dahliae facing each other and sealed with Parafilm
® (American National Can). In the evening, one plastic cap containing 60 olive bark beetles from the PDA plates was placed on the soil near each tree. This method was successfully used in previous works [
23] that demonstrated that adults of fungus gnats and moth flies externally acquired the conidia of
V. dahliae after exposure to the cultures and subsequently became efficient distributors of the pathogen.
Finally, the control treatment consisted of two groups of two healthy olive trees. The first group (Control 1) was covered by an individual mosquito net and exposed to five V. dahliae-free branches that showed entry holes (mating galleries) of the olive bark beetle. A second group (Control 2) remained uncovered and not exposed to P. scarabaeoides adults or damaged olive branches by the beetles, and it was used for confirming that plants were free from infections of V. dahliae.
Several days after inoculation, the number of entry holes (feeding galleries) produced by beetles was recorded in the healthy trees used for the different treatments. During the following months after beetle inoculations (July to September 2009), trees were periodically inspected to identify Verticillium wilt disease symptoms such as wilt, chlorosis, leaf and shoot necrosis, or defoliation.
In September 2009, the damaged branches that were used as a beetle source for healthy trees were removed. By this time, beetles were exiting the galleries they made on the healthy trees, on which they fed during summer, to look for new trees in which to make feeding galleries. Removing branches forced adults to make new feeding galleries in the same trees.
From September 2009 to March 2010, three shoots per tree were collected, and the number of feeding galleries was recorded. The sampling was only conducted in trees with high numbers of entry holes, rejecting trees slightly damaged. Thus, totals of 6, 42, and 24 shoots were collected from 2, 14, and 8 trees from Control 1, T1, and T2, respectively. Adult beetles were periodically extracted from the feeding galleries of these plants to check if the insects were superficially or internally infested by
V. dahliae and/or if olive shoots were infected by the inoculum presumably transmitted by the beetles. Therefore, molecular analyses and microbiological isolations from plant tissues and from adults were done using the nested-PCR technique [
34].
4.3. Experiment III
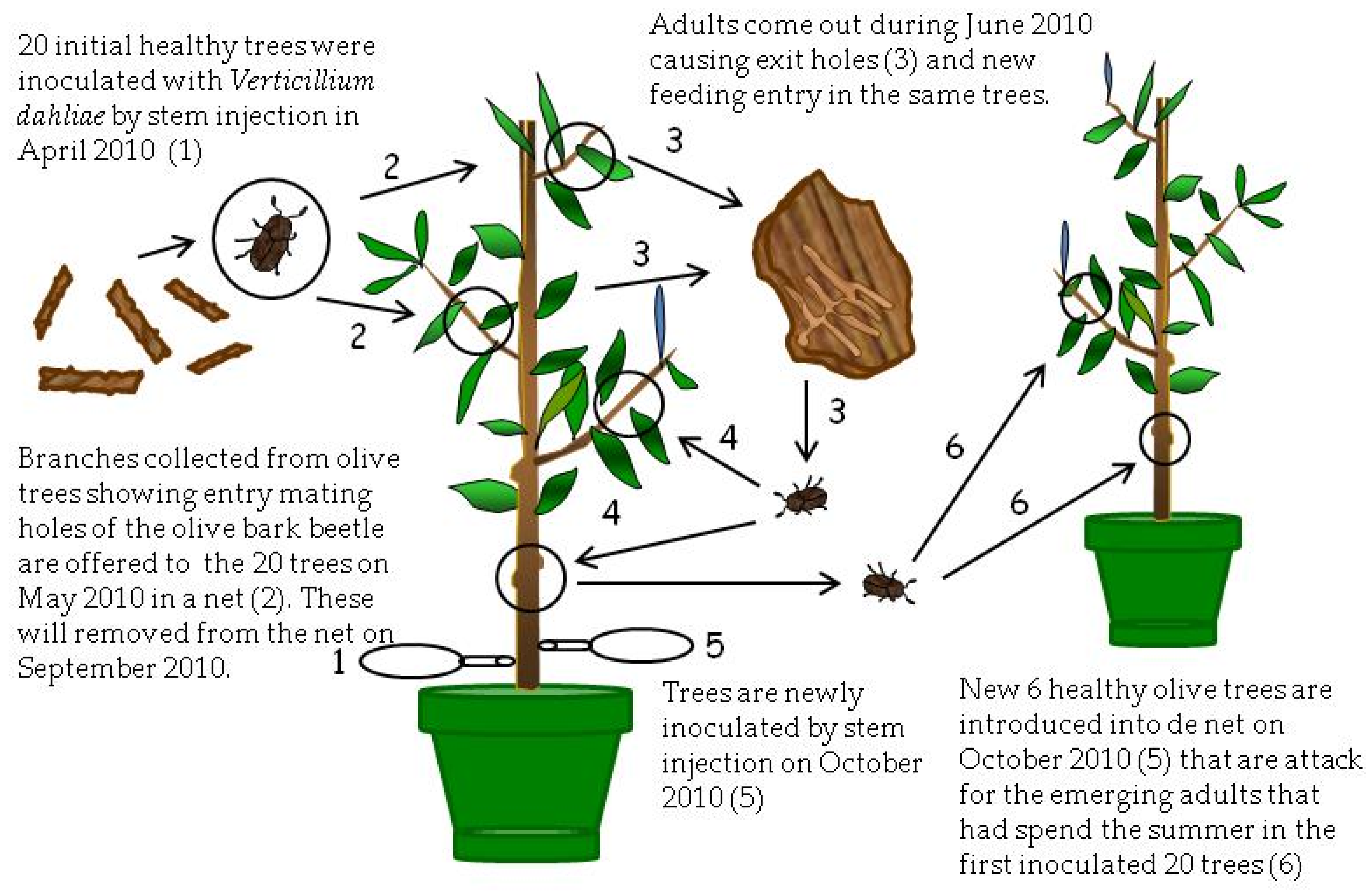
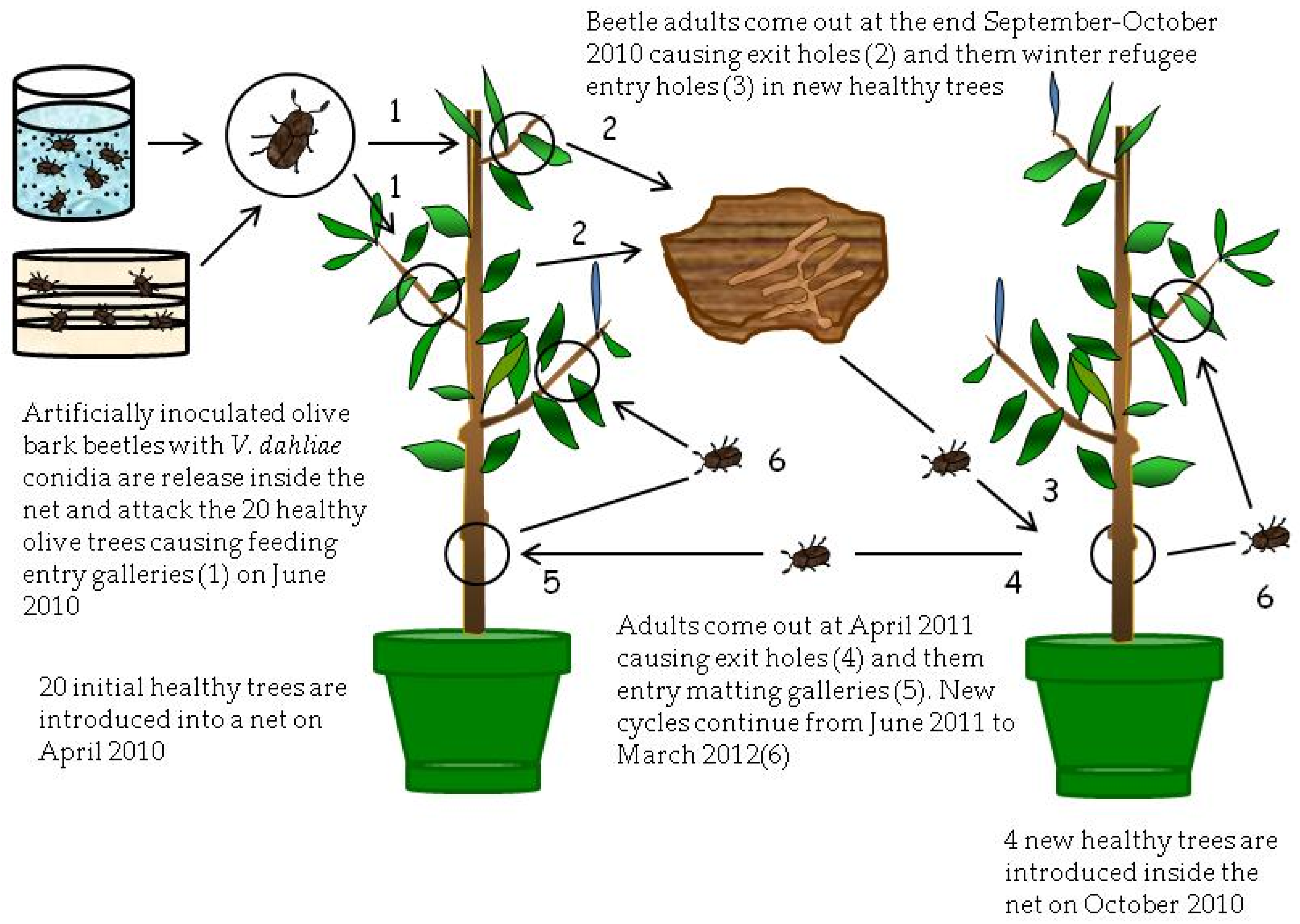
This experiment was carried out from April 2010 to March 2012 in which olive trees from Experiment II were used because Verticillium was not detected in either plants or beetles. The experiment consisted of three treatments (
Figure 3 and
Figure 4).
In the first treatment (T3), twenty trees chosen at random from the treatments T1 and T2 of the Experiment II were placed in April (2010) inside a new mosquito net and were inoculated by stem injection with 50 mL of a conidial suspension of 10
5 conidia/mL of the V117
V. dahliae isolate [
27] (
Figure 3). To produce conidia of
V. dahliae, 15 mL of sterilized distilled water was poured over active colonies of
V. dahliae on the PDA plates and then the surface was gently rubbed with a sterilized glass rod. The mycelial and conidial suspension was passed through a double sterilized cheesecloth yielding a water-suspension of conidia that was finally adjusted to a concentration of 10
7 conidia/mL. Four weeks after inoculation (May 2010), thirty
V. dahliae-free branches collected from the Montilla field with entry holes (mating galleries) of the insect were again introduced in the mosquito net. Adults emerged at the beginning of June 2010 and infested olive trees again, producing feeding galleries (
Figure 3). The aim of these inoculations was to increase pathogen colonization of tissues and the number of beetles in these trees. After inoculations (May 2010), sampling of plant tissue was conducted to confirm the presence of the pathogen in affected shoots by microbiological isolations in PDA. Moreover, during the following months, trees were periodically inspected to assess VWO symptoms and damages caused by the insect. In September 2010, introduced branches on May 2010 were removed from inside the mosquito net. In October 2010, trees of this treatment were again inoculated with
V. dahliae by trunk injection, similar to the previous April. Afterwards, six new, healthy trees with the same characteristics of those used initially in Experiment II and acquired from the same commercial nursery were introduced inside the net. Again, the hypothesis was that adults that spent the summer within inoculated trees and were now abandoning the feeding galleries would be able to transmit the pathogen to the new, healthy trees they were using to build new feeding galleries (
Figure 3). In November 2010, microbiological isolations on PDA plates of symptomatic tissues were conducted using trees of this treatment to confirm the presence of the pathogen in symptomatic shoots.
The second treatment of this experiment (T4) also consisted of another group of twenty trees, chosen at random from treatments T1 and T2 of the Experiment II that were introduced in April 2010 in a new mosquito net (
Figure 4). Trees were exposed during June 2010 to 420 adults of
P. scarabaeoides. These adults were extracted from damaged olive branches collected in the Montilla field and infested with
V. dahliae by two different ways. Half of the adults were infested as described for Experiment II using a sandwich of
V. dahliae PDA culture plates. The other half was infested by immersion for 40 s in a conidial suspension of 10
8 conidia/mL of the V117 isolate. Infested beetles were released into the mosquito net to produce new entry holes (feeding galleries) in the twenty olive trees during the summer feeding. Finally, during October 2010, four healthy trees were introduced inside the net of the treatment T4 to expose them to attack by the beetles from the initial twenty olive trees (
Figure 4).
A third treatment (T5) (not illustrated by a figure) consisted of eight healthy trees that were exposed to thirty olive branches damaged by P. scarabaeoides that showed clear VWO symptoms in April 2011. These branches were collected from an olive orchard in the municipality of Almodóvar near Cordoba.
The control treatment in this experiment consisted of four healthy trees not exposed to the bark beetle.
In June and July 2011, the new generation of beetles came out from trees inside the corresponding net, and the number of entry holes (feeding and mating) produced in tree shoots by beetles was recorded. In addition, samplings of affected shoots from the healthy trees of the three treatments were conducted in November 2011 and March 2012 using the same methods explained above for Experiment II. In these samples, the number of holes (feeding and mating galleries) was recorded. Moreover, infections by V. dahliae of collected plant material were assessed using isolations on PDA and nested-PCR analyses. The same analyses were made for adults of the olive bark beetle recovered from galleries of collected shoots.
4.4. Statistical Analysis of Data
Analysis of variance (ANOVA) of evaluated parameters was performed using Statistix 9.0 program (Analytical Software, Tallahassee, FL, USA). Mean values were compared by the Fisher’s protected LSD test at p = 0.05. Prior to analysis, data were checked in regard to the three main assumptions (homogeneity of variances, normality, and distribution of residuals).




 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}