
Molecular Detection and Phylogeny of Tick-Borne Pathogens in Ticks Collected from Dogs in the Republic of Korea

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Distribution of Dog Ticks in the ROK
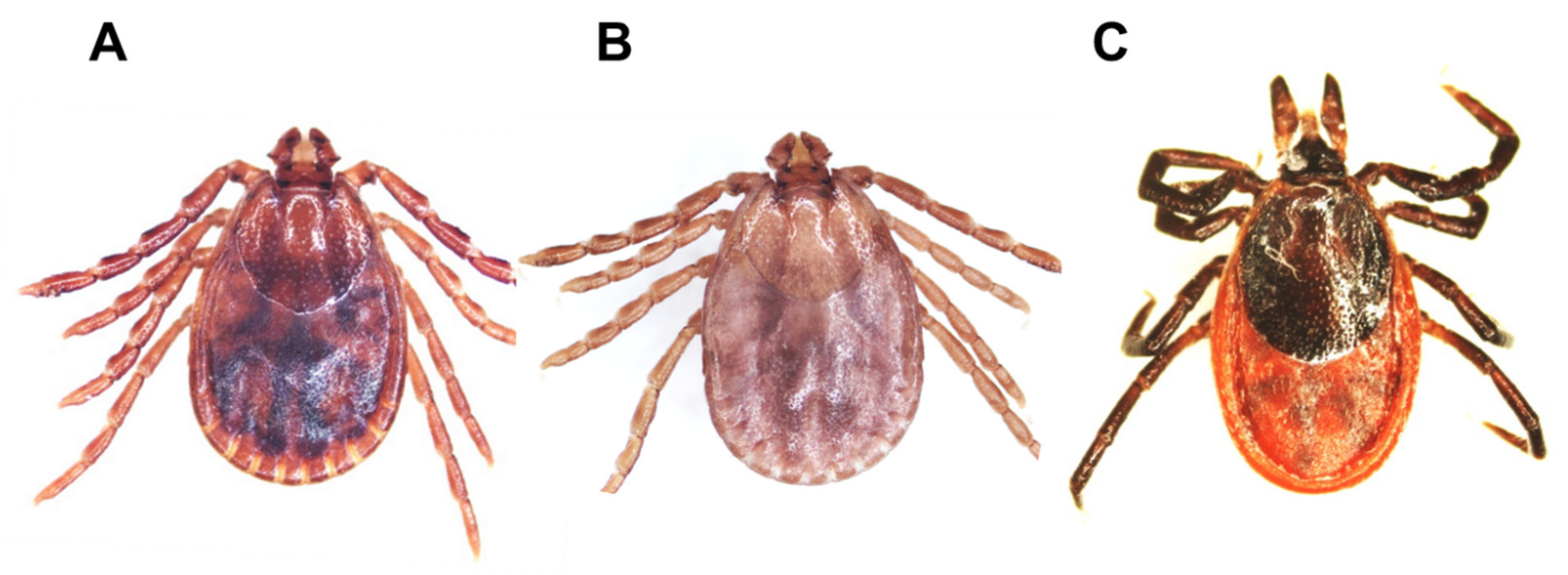
2.2. Identification of Tick Species
2.3. Detection of Tick-Borne Pathogens in Dog Ticks
2.4. Sequencing and Phylogenetic Analysis of Tick-Borne Pathogens
3. Discussion
4. Materials and Methods
4.1. Collection of Ticks
4.2. Identification of Tick Species
4.3. Isolation of Tick Nucleic Acids
4.4. Detection of Tick-Borne Pathogens
4.5. Sequencing and Phylogenetic Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De la Fuente, J.; Estrada-Pena, A.; Venzal, J.M.; Kocan, K.M.; Sonenshine, D.E. Overview: Ticks as vectors of pathogens that cause disease in humans and animals. Front. Biosci. 2008, 13, 6938–6946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogbu, K.I.; Olaolu, O.S.; Ochai, S.O.; Tion, M.T. A review of some tick-borne pathogens of dogs. JASVM 2018, 3, 140–153. [Google Scholar] [CrossRef]
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, 3–14. [Google Scholar] [CrossRef]
- Cao, W.; Zhan, L.; He, J.; Foley, J.E.; de Vlas, S.J.; Wu, X.; Yang, H.; Richardus, J.H.; Habbema, J.D.F. Natural Anaplasma phagocytophilum infection of ticks and rodents from a forest area of Jilin province, China. Am. J. Trop. Med. Hyg. 2006, 75, 664–668. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, N.; Inayoshi, M.; Kitamura, K.; Kawamori, F.; Kawaguchi, D.; Nishimura, Y.; Naitou, H.; Hiroi, M.; Masuzawa, T. Anaplasma phagocytophilum-infected ticks, Japan. Emerg. Infect. Dis. 2005, 11, 1780–1783. [Google Scholar] [CrossRef]
- Shaw, S.E.; Day, M.J.; Birtles, R.J.; Breitschwerdt, E.B. Tick-borne infectious diseases of dogs. Trends Parasitol. 2001, 17, 74–80. [Google Scholar] [CrossRef]
- Biggs, H.M.; Behravesh, C.B.; Bradley, K.K.; Dahlgren, F.S.; Drexler, N.A.; Dumler, J.S.; Folk, S.M.; Kato, C.Y.; Lash, R.R.; Levin, M.L.; et al. Diagnosis and Management of Tickborne Rickettsial Diseases: Rocky Mountain Spotted Fever and Other Spotted Fever Group Rickettsioses, Ehrlichioses, and Anaplasmosis-United States. MMWR Recomm. Rep. 2016, 65, 1–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdullah, S.; Helps, C.; Tasker, S.; Newbury, H.; Wall, R. Ticks infesting domestic dogs in the UK: A large-scale surveillance programme. Parasites Vectors 2016, 9, 391. [Google Scholar] [CrossRef] [Green Version]
- Beck, S.; Schreiber, C.; Schein, E.; Krücken, J.; Baldermann, C.; Pachnicke, S.; Samson-Himmelstjerna, G.V.; Kohn, B. Tick infestation and prophylaxis of dogs in northeastern Germany: A prospective study. Ticks Tick Borne Dis. 2014, 5, 336–342. [Google Scholar] [CrossRef]
- Földvári, G.; Farkas, R. Ixodid tick species attaching to dogs in Hungary. Vet. Parasitol. 2005, 129, 125–131. [Google Scholar] [CrossRef]
- Abdullah, S.; Helps, C.; Tasker, S.; Newbury, H.; Wall, R. Prevalence and distribution of Borrelia and Babesia species in ticks feeding on dogs in the UK. Med. Vet. Entomol. 2018, 32, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Duncan, A.W.; Correa, M.T.; Levine, J.F.; Breitschwerdt, E.B. The dog as a sentinel for human infection: Prevalence of Borrelia burgdorferi C6 antibodies in dogs from southeastern and mid-Atlantic states. Vector Borne Zoonotic Dis. 2004, 4, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Irwin, P.J.; Robertson, I.D.; Westman, M.E.; Perkins, M.; Straubinger, R.K. Searching for Lyme borreliosis in Australia: Results of a canine sentinel study. Parasites Vectors 2017, 10, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, F.D.; Ballantyne, R.; Morgan, E.R.; Wall, R. Estimating Lyme disease risk using pet dogs as sentinels. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Namina, A.; Capligina, V.; Seleznova, M.; Krumins, R.; Aleinikova, D.; Kivrane, A.; Akopjana, S.; Lazovska, M.; Berzina, I.; Ranka, R. Tick-borne pathogens in ticks collected from dogs, Latvia, 2011–2016. BMC Vet. Res. 2019, 15, 398. [Google Scholar] [CrossRef]
- Choi, W.Y. Diagnosis and epidemiology of toxoplasmosis in Korea. Korean J. Parasitol. 1990, 28, 41–44. [Google Scholar] [CrossRef]
- Im, J.H.; Baek, J.; Durey, A.; Kwon, H.Y.; Chung, M.H.; Lee, J.S. Current status of tick-borne diseases in South Korea. Vector Borne Zoonotic Dis. 2019, 19, 4. [Google Scholar] [CrossRef] [PubMed]
- Korea Centers for Disease Control and Prevention (KCDC). Infectious Diseases Surveillance Yearbook 2018; Korea Centers for Disease Control and Prevention (KCDC): Chungbuk, Korea, 2019. [Google Scholar]
- Seo, M.G.; Kwon, O.D.; Kwak, D. Molecular detection and phylogenetic analysis of canine tick-borne pathogens from Korea. Ticks Tick Borne Dis. 2020, 11, 101357. [Google Scholar] [CrossRef]
- Suh, G.H.; Ahn, K.S.; Ahn, J.H.; Kim, H.J.; Leutenegger, C.; Shin, S. Serological and molecular prevalence of canine vector-borne diseases (CVBDs) in Korea. Parasites Vectors 2017, 10, 146. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Goo, Y.K.; Geraldino, P.J.L.; Kwon, O.D.; Kwak, D. Molecular detection and characterization of Borrelia garinii (Spirochaetales: Borreliaceae) in Ixodes nipponensis (Ixodida: Ixodidae) parasitizing a dog in Korea. Pathogens 2019, 8, 289. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.L.; Kim, H.C.; Coburn, J.M.; Chong, S.T.; Chang, N.W.; Robbins, R.G.; Klein, T.A. Tick surveillance in two southeastern provinces, including three metropolitan areas, of the Republic of Korea during 2014. Syst. Appl. Acarol. 2017, 22, 271–288. [Google Scholar] [CrossRef]
- Kim-Jeon, M.D.; Jegal, S.; Jun, H.; Jung, H.; Park, S.H.; Ahn, S.K.; Lee, J.; Gong, Y.W.; Joo, K.; Kwon, M.J.; et al. Four year surveillance of the vector hard ticks for SFTS, Ganghwa-do, Republic of Korea. Korean J. Parasitol. 2019, 57, 691–698. [Google Scholar] [CrossRef]
- Park, J.W.; Lee, S.H.; Lee, G.S.; Seo, J.J.; Chung, J.K. Epidemiological characteristics of field tick-borne pathogens in Gwang-ju metropolitan area, South Korea, from 2014 to 2018. Osong Public Health Res. Perspect. 2020, 11, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Yi, J.Y.; Oh, W.S.; Kim, N.H.; Choi, S.J.; Choe, N.J.; Lee, J.K.; Oh, M.D. Human granuylocytic anaplasmosis, South Korea, 2013. Emerg. Infect. Dis. 2014, 20, 1708–1711. [Google Scholar] [CrossRef]
- Wu, T.J.; Sun, H.J.; Wu, Y.C.; Huang, H.P. Prevalence and risk factors of canine ticks and tick-borne diseases in Taipei, Taiwan. J. Vet. Clin. Sci. 2009, 2, 75–78. [Google Scholar]
- Kamani, J.; Baneth, G.; Mumcuoglu, K.Y.; Waziri, N.E.; Eyal, O.; Guthmann, Y.; Harrus, S. Molecular detection and characterization of tick-borne pathogens in dogs and ticks from Nigeria. PLoS Negl. Trop. Dis. 2013, 7, e2108. [Google Scholar] [CrossRef]
- Livanova, N.N.; Fomenko, N.V.; Akimov, I.A.; Ivanov, M.J.; Tikunova, N.V.; Armstrong, R.; Konyaev, S.V. Dog survey in Russian veterinary hospitals: Tick identification and molecular detection of tick-borne pathogens. Parasites Vectors 2018, 11, 591. [Google Scholar] [CrossRef] [Green Version]
- Palomar, A.M.; Portillo, A.; Santibáñez, P.; Mazuelas, D.; Roncero, L.; García-Álvarez, L.; Santibáñez, S.; Gutiérrez, Ó.; Oteo, J.A. Detection of tick-borne Anaplasma bovis, Anaplasma phagocytophilum and Anaplasma centrale in Spain. Med. Vet. Entomol. 2015, 29, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Polin, H.; Hufnagl, P.; Haunschmid, R.; Gruber, F.; Ladurner, G. Molecular evidence of Anaplasma phagocytophilum in Ixodes ricinus ticks and wild animals in Austria. J. Clin. Microbiol. 2004, 42, 2285–2286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuen, S. Anaplasma Phagocytophilum-the most widespread tick-borne infection in animals in Europe. Vet. Res. Commun. 2007, 31, 79–84. [Google Scholar] [CrossRef]
- Kim, C.M.; Kim, M.S.; Park, M.S.; Park, J.H.; Chae, J.S. Identification of Ehrlichia chaffeensis, Anaplasma phagocytophilum, and A. bovis in Haemaphysalis longicornis and Ixodes persulcatus ticks from Korea. Vector Borne Zoonotic Dis. 2003, 3, 17–26. [Google Scholar] [CrossRef]
- Fraenkel, C.J.; Garpmo, U.; Berglund, J. Determination of novel Borrelia genospecies in Swedish Ixodes ricinus ticks. J. Clin. Microbiol. 2002, 40, 3308–3312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurokawa, C.; Lynn, G.E.; Pedra, J.H.F.; Pal, U.; Narasimhan, S.; Fikrig, E. Interactions between Borrelia burgdorferi and ticks. Nat. Rev. Microbiol. 2020, 18, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, M.; Okada, K.; Nakao, M.; Konishi, T.; Sato, Y. Phylogenetic analysis of Borrelia species based on flagellin gene sequences and its application for molecular typing of Lyme disease borreliae. Int. J. Syst. Bacteriol. 1996, 46, 898–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoogstraal, H.; Roberts, F.H.; Kohls, G.M.; Tipton, V.J. Review of Haemaphysalis (kaiserinana) longicornis Neumann (resurrected) of Australia, New Zealand, New Caledonia, Fiji, Japan, Korea, and Norteastern China and USSR, and its parthenogenetic and bisexual populations (Ixodoidea, Ixodidae). J. Parasitol. 1968, 54, 1197–1213. [Google Scholar] [CrossRef] [Green Version]
- Hoogstraal, H.; Wassef, H.Y. The Haemaphysalis ticks (Ixodoidea: Ixodidae) of birds. 3. H. (ornithophysalis) subgen. n.: Definition, species, hosts, and distribution in the Oriental, Palearctic, Malagasy, and Ethiopian faunal regions. J. Parasitol. 1973, 59, 1099–1117. [Google Scholar]
- Yamaguti, N.; Tipton, V.J.; Keegan, H.L.; Toshioka, S. Ticks of Japan, Korea, and the Ryukyu Islands. Brigham Young Univ. Sci. Bull. Biol. Ser. 1971, 15, 1–226. [Google Scholar]
- Hancock, S.I.; Breitschwerdt, E.B.; Pitulle, C. Differentiation of Ehrlichia platys and E. equi infections in dogs by using 16S ribosomal DNA-based PCR. J. Clin. Microbiol. 2001, 39, 4577–4578. [Google Scholar] [CrossRef] [Green Version]
- Mathew, J.S.; Ewing, S.A.; Murphy, G.L.; Kocan, K.M.; Corstvet, R.E.; Fox, J.C. Characterization of a new isolate of Ehrlichia platys (Order Rickettsiales) using electron microscopy and polymerase chain reaction. Vet. Parasitol. 1997, 68, 1–10. [Google Scholar] [CrossRef]
- Anderson, B.E.; Sumner, J.W.; Dawson, J.E.; Tzianabos, T.; Greene, C.R.; Olson, J.G.; Fishbein, D.B.; Olsen-Rasmussen, M.; Holloway, B.P.; George, E.H. Detection of the etiologic agent of human ehrlichiosis by polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 775–780. [Google Scholar] [CrossRef] [Green Version]
- Murphy, G.L.; Ewing, S.A.; Whitworth, L.C.; Fox, J.C.; Kocan, A.A. A molecular and serologic survey of Ehrlichia canis, E. chaffeensis, and E. ewingii in dogs and ticks from Oklahoma. Vet. Parasitol. 1998, 79, 325–339. [Google Scholar] [CrossRef]
- Kang, J.G.; Ko, S.; Smith, W.B.; Kim, H.C.; Lee, I.Y.; Chae, J.S. Prevalence of Anaplasma, Bartonella and Borrelia species in Haemaphysalis longicornis collected from goats in North Korea. J. Vet. Sci. 2016, 17, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Ano, H.; Makimura, S.; Harasawa, R. Detection of Babesia species from infected dog blood by polymerase chain reaction. J. Vet. Med. Sci. 2001, 63, 111–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkin, M.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Stage | Tick Pool (Number of Ticks) | Total | |||
|---|---|---|---|---|---|---|
| Pet Dogs | Stray Dogs | Dog Shelters 1 | Military Working Dogs | |||
| Haemaphysalis longicornis | Larvae | 4 (49) | 26 (126) | 4 (166) | 0 | 34 (341) |
| Nymph | 73 (606) | 51 (149) | 9 (93) | 15 (110) | 148 (958) | |
| Adult | 111 (129) | 282 (295) | 29 (29) | 478 (509) | 900 (962) | |
| Sub total | 188 (784) | 359 (570) | 42 (288) | 493 (619) | 1082 (2261) | |
| H. flava | Larvae | 0 | 0 | 0 | 0 | 0 |
| Nymph | 1 (3) | 0 | 1 (1) | 1 (3) | 3 (7) | |
| Adult | 2 (2) | 0 | 0 | 1 (1) | 3 (3) | |
| Sub total | 3 (5) | 0 (0) | 1 (1) | 2 (4) | 6 (10) | |
| Ixodes nipponensis | Larvae | 0 | 0 | 0 | 0 | 0 |
| Nymph | 0 | 0 | 0 | 0 | 0 | |
| Adult | 18 (18) | 2 (2) | 1 (1) | 1 (1) | 22 (22) | |
| Sub total | 18 (18) | 2 (2) | 1 (1) | 1 (1) | 22 (22) | |
| Total | 209 (807) | 361 (572) | 44 (290) | 496 (624) | 1110 (2293) | |
| Region | Site | Species | Total (%) | ||
|---|---|---|---|---|---|
| Haemaphysalis longicornis | H. flava | Ixodes nipponensis | |||
| Northern | Pet dogs | 552 | 4 | 12 | 568 |
| Stray dogs | 189 | 0 | 1 | 190 | |
| Dog shelters 1 | 100 | 0 | 0 | 100 | |
| Military working dogs | 599 | 3 | 1 | 603 | |
| Subtotal | 1440 (62.80) | 7 (0.31) | 14 (0.61) | 1461 (63.72) | |
| Central | Pet dogs | 228 | 1 | 6 | 235 |
| Stray dogs | 321 | 0 | 1 | 322 | |
| Dog shelters | 188 | 1 | 1 | 190 | |
| Military working dogs | 20 | 0 | 0 | 20 | |
| Subtotal | 757 (33.01) | 2 (0.09) | 8 (0.35) | 767 (33.45) | |
| Southern | Pet dogs | 4 | 0 | 0 | 4 |
| Stray dogs | 60 | 0 | 0 | 60 | |
| Dog shelters | 0 | 0 | 0 | 0 | |
| Military working dogs | 0 | 1 | 0 | 1 | |
| Subtotal | 64 (2.79) | 1 (0.04) | 0 | 65 (2.83) | |
| Total (%) | 2261 (98.60) | 10 (0.44) | 22 (0.96) | 2293 (100) | |
| Scheme | Stage | Number of Tick Pools | Species | |||||
|---|---|---|---|---|---|---|---|---|
| Anaplasma spp. | A. platys | E. canis | E. chaffeensis | Borrelia spp. | B. gibsoni | |||
| Haemaphysalis longicornis | Larvae | 34 | 3 | - | - | - | - | - |
| Nymph | 148 | 5 | - | - | - | - | - | |
| Adult | 900 | 14 | - | - | - | 1 | - | |
| Subtotal | 1082 | 22 (2.03) | - | - | - | 1 (0.09) | - | |
| H. flava | Larvae | 0 | - | - | - | - | - | - |
| Nymph | 3 | - | - | - | - | - | - | |
| Adult | 3 | - | - | - | - | - | - | |
| Subtotal | 6 | - | - | - | - | - | - | |
| Ixodes nipponensis | Larvae | 0 | - | - | - | - | - | - |
| Nymph | 0 | - | - | - | - | - | - | |
| Adult | 22 | 2 | - | - | - | 3 | - | |
| Subtotal | 22 | 2 (9.09) | - | - | - | 3 (13.64) | - | |
| Total | 1110 | 24 (2.16) | 0 | 0 | 0 | 4 (0.36) | 0 | |
| Region | Site | Number of Tick Pools | Species | |||||
|---|---|---|---|---|---|---|---|---|
| Anaplasma spp. | A. platys | E. canis | E. chaffeensis | Borrelia spp. | B. gibsoni | |||
| Northern | Pet dogs | 84 | 1 (1.19) | - | - | - | 1 (1.19) | - |
| Stray dogs | 169 | - | - | - | - | 1 (0.59) | - | |
| Dog shelters 1 | 2 | - | - | - | - | - | - | |
| Military working dogs | 482 | 11 (2.28) | - | - | - | - | - | |
| Subtotal | 737 | 12 (1.63) | - | - | - | 2 (0.27) | - | |
| Central | Pet dogs | 121 | 3 (2.48) | - | - | - | 2 (1.65) | - |
| Stray dogs | 169 | 5 (2.96) | - | - | - | - | - | |
| Dog shelters | 42 | - | - | - | - | - | - | |
| Military working dogs | 13 | - | - | - | - | - | - | |
| Subtotal | 345 | 8 (2.32) | - | - | - | 2 (0.58) | - | |
| Southern | Pet dogs | 4 | - | - | - | - | - | - |
| Stray dogs | 23 | 4 (17.39) | - | - | - | - | - | |
| Dog shelters | 0 | - | - | - | - | - | - | |
| Military working dogs | 1 | - | - | - | - | - | - | |
| Subtotal | 28 | 4 (14.29) | - | - | - | 0 | - | |
| Total (%) | 1110 | 24 (2.16) | 0 | 0 | 0 | 4 (0.36) | 0 | |
| Pathogens | Primers | Sequences (5′-3′) | Target Gene (bp) | PCR Conditions | References |
|---|---|---|---|---|---|
| Anaplasma spp. | PITA-F | GTCGAACGGATTATTCTTTA | 16S rRNA (511) | 95 °C (5 min); 37 cycles of 95 °C (30 s), 50 °C (30 s), and 72 °C (40 s); 72 °C (7 min) | [39] |
| PITA-R | TTCACCTTTAACTTACCGAA | ||||
| A. platys | EPLAT5 | TTTGTCGTAGCTTGCTATGAT | 16S rRNA (359) | 95 °C (5 min); 37 cycles of 95 °C (30 s), 53 °C (30 s), and 72 °C (30 s); 72 °C (7 min) | [40] |
| EPLAT3 | CTTCTGTGGGTACCGTC | ||||
| Ehrlichia canis | ECAN5 | GCAAATTATTTATAGCCTCTGGCTATAG | 16S rRNA (365) | 95 °C (5 min); 37 cycles of 95 °C (30 s), 56 °C (30 s), and 72 °C (30 s); 72 °C (7 min) | [41,42] |
| HE3 | TTATAGGTACCGTCATTATCTTCCCTA | ||||
| E. chaffeensis | HE1 | ACAATATTGCTTATAACCTTTTGGTTATA | 16S rRNA (390) | 95 °C (5 min); 37 cycles of 95 °C (30 s), 56 °C (30 s), and 72 °C (30 s); 72 °C (7 min) | |
| HE3 | TTATAGGTACCGTCATTATCTTCCCTA | ||||
| Borrelia spp. | B3 | GCAGCTAAGAATCTTCCGCA | 16S rRNA (714) | 95 °C (5 min); 37 cycles of 95 °C (30 s), 58 °C (30 s), and 72 °C (1 min); 72 °C (7 min) | [43] |
| B6 | CAACCATGCAGCACCTGTATAT | ||||
| Babesia gibsoni | PIRO-F | AGTCATATGCTTGTCTTA | 18S rRNA (500) | 95 °C (5 min); 37 cycles of 95 °C (30 s), 47 °C (30 s), and 72 °C (40 s); 72 °C (7 min) | [44] |
| PIRO-R | CCATCATTCCAATTACAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truong, A.-T.; Noh, J.; Park, Y.; Seo, H.-J.; Kim, K.-H.; Min, S.; Lim, J.; Yoo, M.-S.; Kim, H.-C.; Klein, T.A.; et al. Molecular Detection and Phylogeny of Tick-Borne Pathogens in Ticks Collected from Dogs in the Republic of Korea. Pathogens 2021, 10, 613. https://doi.org/10.3390/pathogens10050613
Truong A-T, Noh J, Park Y, Seo H-J, Kim K-H, Min S, Lim J, Yoo M-S, Kim H-C, Klein TA, et al. Molecular Detection and Phylogeny of Tick-Borne Pathogens in Ticks Collected from Dogs in the Republic of Korea. Pathogens. 2021; 10(5):613. https://doi.org/10.3390/pathogens10050613
Chicago/Turabian StyleTruong, A-Tai, Jinhyeong Noh, Yeojin Park, Hyun-Ji Seo, Keun-Ho Kim, Subin Min, Jiyeon Lim, Mi-Sun Yoo, Heung-Chul Kim, Terry A. Klein, and et al. 2021. "Molecular Detection and Phylogeny of Tick-Borne Pathogens in Ticks Collected from Dogs in the Republic of Korea" Pathogens 10, no. 5: 613. https://doi.org/10.3390/pathogens10050613