
Prevalence and Genetic Diversity of Avian Haemosporidian Parasites in Southern Iran


Abstract
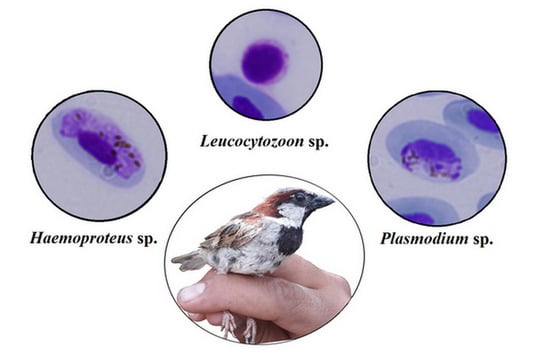
:
1. Introduction
2. Results
2.1. Parasite Prevalence
2.2. Lineage Diversity
2.3. Co-Infection
3. Discussion
4. Materials and Methods
4.1. Collection of the Samples
4.2. DNA Extraction, PCR and Sequencing
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Valkiūnas, G. Avian Malaria Parasites and other Haemosporidia; Apple Academic Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Atkinson, C.T.; Thomas, N.J.; Hunter, D.B. Parasitic Diseases of Wild Birds; John Wiley & Sons: Ames, IA, USA, 2008; pp. 3–107. [Google Scholar]
- Telford, S.R., Jr. Haemoparasites of the Reptilia, Color Atlas and Text; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Clark, N.J.; Clegg, S.M.; Lima, M.R. A review of global diversity in avian haemosporidians (Plasmodium and Haemoproteus: Haemosporida): New insights from molecular data. Int. J. Parasitol. 2014, 44, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Marzal, A.; Bensch, S.; Reviriego, M.I.; Balbontin, J.; De Lope, F. Effects of malaria double infection in birds: One plus one is not two. J. Evol. Biol. 2008, 21, 979–987. [Google Scholar] [CrossRef]
- Dimitrov, D.; Zehtindjiev, P.; Bensch, S. Genetic diversity of avian blood parasites in SE Europe: Cytochrome b lineages of the genera Plasmodium and Haemoproteus (Haemosporida) from Bulgaria. Acta Parasitol. 2010, 55, 201–209. [Google Scholar] [CrossRef]
- Lapointe, D.A.; Atkinson, C.T.; Samuel, M.D. Ecology and conservation biology of avian malaria. Ann. N.Y. Acad. Sci. 2012, 1249, 211–226. [Google Scholar] [CrossRef]
- Graham, A.L.; Lamb, T.J.; Read, A.F.; Allen, J.E. Malaria-Filaria Coinfection in Mice Makes Malarial Disease More Severe unless Filarial Infection Achieves Patency. J. Infect. Dis. 2005, 191, 410–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidar, P.; Morton, E.S. Are Multiple Infections More Severe for Purple Martins (Progne Subis) Than Single Infections? Auk 2006, 123, 141–147. [Google Scholar] [CrossRef]
- Atkinson, C.T.; Van Riper, C., III. Pathogenicity and epizootiology of avian haematozoa: Plasmodium, Leucocytozoon, and Haemoproteus. In Bird-Parasite Interactions: Ecology, Evolution, and Behaviour, 2rd ed.; Oxford University Press: Cary, NC, USA; New York, NY, USA, 1991; pp. 19–48. [Google Scholar]
- Bensch, S.; Stjernman, M.; Hasselquist, D.; Örjan, Ö.; Hansson, B.; Westerdahl, H.; Pinheiro, R.T. Host specificity in avian blood parasites: A study of Plasmodium and Haemoproteus mitochondrial DNA amplified from birds. Proc. R. Soc. B Boil. Sci. 2000, 267, 1583–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bensch, S.; Hellgren, O.; Pérez-Tris, J. MalAvi: A public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Mol. Ecol. Resour. 2009, 9, 1353–1358. [Google Scholar] [CrossRef]
- Okwa, O. Malaria Parasites; InTech: Rijeka, Croatia, 2012. [Google Scholar]
- MalAvi. A Database for Avian Haemosporidian Parasites. Table: Host and Sites. Available online: http:// mbio-serv2.mbioekol.lu.se/ Malavi/ (accessed on 23 April 2021).
- Jarvi, S.I.; Schultz, J.J.; Atkinson, C.T. PCR diagnostics underestimate the prevalence of avian malaria (Plasmodium relictum) in experimentally-infected passerines. J. Parasitol 2002, 88, 153–158. [Google Scholar] [CrossRef]
- Pellegrino, I.; Ilahiane, L.; Boano, G.; Cucco, M.; Pavia, M.; Prestridge, H.; Voelker, G. Avian Haemosporidian Diversity on Sardinia: A First General Assessment for the Insular Mediterranean. Diversity 2021, 13, 75. [Google Scholar] [CrossRef]
- Van Hemert, C.; Meixell, B.W.; Smith, M.M.; Handel, C.M. Prevalence and diversity of avian blood parasites in a resident northern passerine. Parasit Vectors 2019, 12, 292. [Google Scholar] [CrossRef]
- Khaleghizadeh, A.; Roselaar, K.; Scott, D.A.; Tohidifar, M.; Mlíkovský, J.; Blair, M.; Kvartalnov, P. Birds of Iran: Annotated Checklist of the Species and Subspecies; Iranian Research Institute of Plant Protection: Tehran, Iran, 2017. [Google Scholar]
- Madjnoonian, H.; Kiabi, B.H.; Danesh, M. Readings in Zoogeography of Iran: Part I; Department of Environment: Tehran, Iran, 2005. [Google Scholar]
- Evans, M.I. Important Bird Areas in the Middle East; Birdlife International: Cambridge, UK, 1994. [Google Scholar]
- Nazifi, S.; Razavi, S.M.; Yavari, F.; Rajaifar, M.; Bazyar, E.; Esmailnejad, Z. Evaluation of hematological values in indigenous chickens infected with Plasmodium gallinaceum and Aegyptianella pullorum. Comp. Haematol. Int. 2008, 17, 145–148. [Google Scholar] [CrossRef]
- Radfar, M.H.; Fathi, S.; Asl, E.N.; Dehaghi, M.M.; Seghinsara, H.R. A survey of parasites of domestic pigeons (Columba livia domestica) in South Khorasan, Iran. Vet. Res. 2011, 4, 18–23. [Google Scholar] [CrossRef]
- Gorji, S.F.; Shemshadi, B.; Habibi, H.; Jalali, S.; Davary, M.; Sepehri, M. Survey on parasites in sparrows of Amol (Mazandaran Province, Iran). J. Life Sci. 2012, 6, 783–785. [Google Scholar]
- Dehghani Samani, A.; Pirali Kheirabadi, K. Prevalence and Rate of Parasitemia of Haemoproteus columbae in Columba Iivia domestica Southwest of Iran. Iran J. Parasitol. 2013, 8, 641–644. [Google Scholar] [PubMed]
- Dezfoulian, O.; Zibaei, M.; Nayebzadeh, H.; Haghgoo, M.; Emami-Razavi, A.; Kiani, K. Leucocytozoonosis in Domestic Birds in Southwestern Iran: An Ultrastructural Study. Iran J. Parasitol. 2013, 8, 171–176. [Google Scholar] [PubMed]
- Doosti, A.; Mohammadalipour, Z.; Zohoor, A. Detection of Haemoproteus Columbae in Iranian Pigeons Using PCR. Int. Conf. Biol. Civ. Environ. Eng. (BCEE-2014) 2014, 36, 8. [Google Scholar] [CrossRef]
- Shemshadi, B.; Ranjbar-Bahadori, S.; Faghihzadeh-Gorgi, S. Occurrence of parasitic protozoa in wild waterfowl in southern coastal Caspian Sea lagoons. Iran J. Vet. Med. 2015, 8, 261–267. [Google Scholar] [CrossRef]
- Tavassoli, M.; Esmaeilnejad, B.; Malekifard, F.; Mardani, K. PCR-RFLP detection of Haemoproteus spp. (Haemosporida: Haemoproteidae) in pigeon blood samples from Iran. Bulg. J. Veter-Med. 2018, 21, 429–435. [Google Scholar] [CrossRef]
- Mohaghegh, M.A.; Namdar, F.; Azami, M.; Ghomashlooyan, M.; Kalani, H.; Hejazi, S.H. The first report of blood parasites in the birds in Fasa district, Southern Iran. Comp. Haematol. Int. 2018, 27, 289–293. [Google Scholar] [CrossRef]
- Hashemi, S.H.; Sargazi, D.; Anvari, D. The study of Lico contamination by Haemoproteus spp in Sistan region. Biol. Forum 2015, 7, 757–758. [Google Scholar]
- Nourani, L.; Aliabadian, M.; Mirshamsi, O.; Djadid, N.D. Correction: Molecular detection and genetic diversity of avian haemosporidian parasites in Iran. PLoS ONE 2019, 14, e0212453. [Google Scholar] [CrossRef] [PubMed]
- Cracraft, J. Deep-history biogeography: Retrieving the historical pattern of evolving continental biotas. Syst. Biol. 1988, 37, 221–236. [Google Scholar] [CrossRef]
- Dickinson, E.C.; Christidis, L. The Howard and Moore Complete Checklist of the Birds of the World: Passerines, 4th ed.; Aves Press: Eastbourne, UK, 2014. [Google Scholar]
- Cornuault, J.; Khimoun, A.; Harrigan, R.J.; Bourgeois, Y.X.C.; Mila, B.; Thébaud, C.; Heeb, P. The role of ecology in the geographical separation of blood parasites infecting an insular bird. J. Biogeogr. 2013, 40, 1313–1323. [Google Scholar] [CrossRef]
- Hernández-Lara, C.; Gonzalez-Garcia, F.; Santiago-Alarcon, D. Spatial and seasonal variation of avian malaria infections in five different land use types within a Neotropical montane forest matrix. Landsc. Urban Plan. 2017, 157, 151–160. [Google Scholar] [CrossRef]
- Martinsen, E.S.; Paperna, I.; Schall, J.J. Morphological versus molecular identification of avian Haemosporidia: An exploration of three species concepts. Parasitology 2006, 133, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Marzal, A.; Ricklefs, R.E.; Valkiūnas, G.; Albayrak, T.; Arriero, E.; Bonneaud, C.; Czirják, G.Á.; Ewen, J.; Hellgren, O.; Hořáková, D.; et al. Diversity, Loss, and Gain of Malaria Parasites in a Globally Invasive Bird. PLoS ONE 2011, 6, e21905. [Google Scholar] [CrossRef] [Green Version]
- Martinsen, E.S.; Perkins, S.L.; Schall, J.J. A three-genome phylogeny of malaria parasites (Plasmodium and closely related genera): Evolution of life-history traits and host switches. Mol. Phylogenetics Evol. 2008, 47, 261–273. [Google Scholar] [CrossRef]
- Loiseau, C.; Zoorob, R.; Robert, A.; Chastel, O.; Julliard, R.; Sorci, G. Plasmodium relictum infection and MHC diversity in the house sparrow (Passer domesticus). Proc. R. Soc. B Boil. Sci. 2010, 278, 1264–1272. [Google Scholar] [CrossRef] [Green Version]
- López, G.; Muñoz, J.; Soriguer, R.; Figuerola, J. Increased Endoparasite Infection in Late-Arriving Individuals of a Trans-Saharan Passerine Migrant Bird. PLoS ONE 2013, 8, e61236. [Google Scholar] [CrossRef] [Green Version]
- Mata, V.A.; Da Silva, L.P.; Lopes, R.J.; Drovetski, S.V. The Strait of Gibraltar poses an effective barrier to host-specialised but not to host-generalised lineages of avian Haemosporidia. Int. J. Parasitol. 2015, 45, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Calero-Riestra, M.; García, J.T. Sex-dependent differences in avian malaria prevalence and consequences of infections on nestling growth and adult condition in the Tawny pipit, Anthus campestris. Malar. J. 2016, 15, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-López, R.; La Puente, J.M.-D.; Gangoso, L.; Yan, J.; Soriguer, R.C.; Figuerola, J. Do mosquitoes transmit the avian malaria-like parasite Haemoproteus? An experimental test of vector competence using mosquito saliva. Parasites Vectors 2016, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Ayadi, T.; Selmi, S.; Hammouda, A.; Reis, S.; Boulinier, T.; Loiseau, C. Diversity, prevalence and host specificity of avian parasites in southern Tunisian oases. Parasitology 2017, 145, 971–978. [Google Scholar] [CrossRef] [Green Version]
- Emmenegger, T.; Bauer, S.; Dimitrov, D.; Marin, J.O.; Zehtindjiev, P.; Hahn, S. Host migration strategy and blood parasite infections of three sparrow species sympatrically breeding in Southeast Europe. Parasitol. Res. 2018, 117, 3733–3741. [Google Scholar] [CrossRef]
- Garcia-Longoria, L.; Marzal, A.; De Lope, F.; Garamszegi, L. Host-parasite interaction explains variation in the prevalence of avian haemosporidians at the community level. PLoS ONE 2019, 14, e0205624. [Google Scholar] [CrossRef] [Green Version]
- Waldenström, J.; Bensch, S.; Kiboi, S.; Hasselquist, D.; Ottosson, U. Cross-species infection of blood parasites between resident and migratory songbirds in Africa. Mol. Ecol. 2002, 11, 1545–1554. [Google Scholar] [CrossRef] [Green Version]
- Drovetski, S.V.; Aghayan, S.A.; Mata, V.; Lopes, R.J.; Mode, N.A.; Harvey, J.A.; Voelker, G. Does the niche breadth or trade-off hypothesis explain the abundance-occupancy relationship in avian Haemosporidia? Mol. Ecol. 2014, 23, 3322–3329. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Alarcon, D.; MacGregor-Fors, I.; Kühnert, K.; Segelbacher, G.; Schaefer, H.M. Avian haemosporidian parasites in an urban forest and their relationship to bird size and abundance. Urban Ecosyst. 2016, 19, 331–346. [Google Scholar] [CrossRef]
- Peev, S.; Zehtindjiev, P.; Ilieva, M.; Träff, J.; Briedis, M.; Adamík, P. Haemosporidian blood parasite diversity and prevalence in the semi-collared flycatcher (Ficedula semitorquata) from the eastern Balkans. Parasitol. Int. 2016, 65, 613–617. [Google Scholar] [CrossRef]
- Stanković, D.; Jönsson, J.; Raković, M. Diversity of avian blood parasites in wild passerines in Serbia with special reference to two new lineages. J. Ornithol. 2019, 160, 545–555. [Google Scholar] [CrossRef]
- Hellgren, O.; Waldenström, J.; Pérez-Tris, J.; Szöll, E.; Si, Ö.; Hasselquist, D.; Križanauskienė, A.; Ottosson, U.; Bensch, S. Detecting shifts of transmission areas in avian blood parasites—A phylogenetic approach. Mol. Ecol. 2007, 16, 1281–1290. [Google Scholar] [CrossRef]
- Zehtindjiev, P.; Ilieva, M.; Križanauskienė, A.; Oparina, O.; Oparin, M.; Bensch, S. Occurrence of haemosporidian parasites in the paddyfield warbler, Acrocephalus agricola (Passeriformes, Sylviidae). Acta Parasitol. 2009, 54, 295–300. [Google Scholar] [CrossRef]
- Spurgin, L.G.; Illera, J.C.; Padilla, D.P.; Richardson, D.S. Biogeographical patterns and co-occurrence of pathogenic infection across island populations of Berthelot’s pipit (Anthus berthelotii). Oecologia 2012, 168, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Rojo, M.A.; Campos, F.; Santamaría, T.; Hernández, M.A. Haemosporidians in Iberian Bluethroats Luscinia svecica. Ardeola 2014, 61, 135–143. [Google Scholar] [CrossRef]
- Ivanova, K.; Zehtindjiev, P.; Mariaux, J.; Dimitrov, D.; Georgiev, B.B. Avian haemosporidians from rain forests in Madagascar: Molecular and morphological data of the genera Plasmodium, Haemoproteus and Leucocytozoon. Infect. Genet. Evol. 2018, 58, 115–124. [Google Scholar] [CrossRef]
- Ortego, J.; Calabuig, G.; Cordero, P.J.; Aparicio, J.M. Genetic characterization of avian malaria (Protozoa) in the endangered lesser kestrel, Falco naumanni. Parasitol. Res. 2007, 101, 1153–1156. [Google Scholar] [CrossRef] [PubMed]
- Illera, J.C.; Emerson, B.C.; Richardson, D.S. Genetic characterization, distribution and prevalence of avian pox and avian malaria in the Berthelot’s pipit (Anthus berthelotii) in Macaronesia. Parasitol. Res. 2008, 103, 1435–1443. [Google Scholar] [CrossRef]
- Synek, P.; Albrecht, T.; Vinkler, M.; Schnitzer, J.; Votýpka, J.; Munclinger, P. Haemosporidian parasites of a European passerine wintering in South Asia: Diversity, mixed infections and effect on host condition. Parasitol. Res. 2013, 112, 1667–1677. [Google Scholar] [CrossRef] [PubMed]
- Van Riper, C.; Van Riper, S.G.; Goff, M.L.; Laird, M. The Epizootiology and Ecological Significance of Malaria in Hawaiian Land Birds. Ecol. Monogr. 1986, 56, 327–344. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Ilgūnas, M.; Bukauskaitė, D.; Fragner, K.; Weissenböck, H.; Atkinson, C.T.; Iezhova, T.A. Characterization of Plasmodium relictum, a cosmopolitan agent of avian malaria. Malar. J. 2018, 17, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Rooyen, J.; Lalubin, F.; Glaizot, O.; Christe, P. Avian haemosporidian persistence and co-infection in great tits at the individual level. Malar. J. 2013, 12, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valkiūnas, G.; Iezhova, T.A.; Shapoval, A.P. High prevalence of blood parasites in hawfinch Coccothraustes coccothraustes. J. Nat. Hist. 2003, 37, 2647–2652. [Google Scholar] [CrossRef]
- Scaglione, F.E.; Pregel, P.; Cannizzo, F.T.; Pérez-Rodríguez, A.D.; Ferroglio, E.; Bollo, E. Prevalence of new and known species of haemoparasites in feral pigeons in northwest Italy. Malar. J. 2015, 14, 99. [Google Scholar] [CrossRef] [Green Version]
- Meixell, B.W.; Arnold, T.W.; Lindberg, M.S.; Smith, M.M.; Runstadler, J.A.; Ramey, A.M. Detection, prevalence, and transmission of avian haematozoa in waterfowl at the Arctic/sub-Arctic interface: co-infections, viral interactions, and sources of variation. Parasites Vectors 2016, 9, 390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernotienė, R.; Palinauskas, V.; Iezhova, T.; Murauskaitė, D.; Valkiūnas, G. Avian haemosporidian parasites (Haemosporida): A comparative analysis of different polymerase chain reaction assays in detection of mixed infections. Exp. Parasitol. 2016, 163, 31–37. [Google Scholar] [CrossRef]
- Pacheco, M.A.; Cepeda, A.S.; Bernotienė, R.; Lotta, I.A.; Matta, N.E.; Valkiūnas, G.; Escalante, A.A. Primers targeting mitochondrial genes of avian haemosporidians: PCR detection and differential DNA amplification of parasites belonging to different genera. Int. J. Parasitol. 2018, 48, 657–670. [Google Scholar] [CrossRef]
- Ciloglu, A.; Ellis, V.A.; Bernotienė, R.; Valkiūnas, G.; Bensch, S. A new one-step multiplex PCR assay for simultaneous detection and identification of avian haemosporidian parasites. Parasitol. Res. 2019, 118, 191–201. [Google Scholar] [CrossRef]
- Pérez-Tris, J.; Bensch, S. Diagnosing genetically diverse avian malarial infections using mixed-sequence analysis and TA-cloning. Parasitology 2005, 131, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Zehtindjiev, P.; Križanauskienė, A.; Bensch, S.; Palinauskas, V.; Asghar, M.; Dimitrov, D.; Scebba, S.; Valkiūnas, G. A New Morphologically Distinct Avian Malaria Parasite That Fails Detection by Established Polymerase Chain Reaction–Based Protocols for Amplification of the Cytochrome B Gene. J. Parasitol. 2012, 98, 657–665. [Google Scholar] [CrossRef]
- Richard, F.A.; Sehgal, R.N.; Jones, H.I.; Smith, T.B. A comparative analysis of PCR-based detection methods for avian malaria. J. Parasitol. 2002, 88, 819–822. [Google Scholar] [CrossRef]
- Beadell, J.S.; Fleischer, R.C. A Restriction Enzyme–Based Assay to Distinguish Between Avian Haemosporidians. J. Parasitol. 2005, 91, 683–685. [Google Scholar] [CrossRef]
- Martinez, J.; de la Puente, J.M.; Herrero, J.; Del Cerro, S.; Lobato, E.; Aguilar, J.R.-D.; Vásquez, R.A.; Merino, S. A restriction site to differentiate Plasmodium and Haemoproteus infections in birds: On the inefficiency of general primers for detection of mixed infections. Parasitology 2009, 136, 713–722. [Google Scholar] [CrossRef] [Green Version]
- Dimitrov, D.; Zehtindjiev, P.; Bensch, S.; Ilieva, M.; Iezhova, T.; Valkiūnas, G. Two new species of Haemoproteus Kruse, 1890 (Haemosporida, Haemoproteidae) from European birds, with emphasis on DNA barcoding for detection of haemosporidians in wildlife. Syst. Parasitol. 2014, 87, 135–151. [Google Scholar] [CrossRef]
- Dimitrov, D.; Palinauskas, V.; Iezhova, T.A.; Bernotienė, R.; Ilgūnas, M.; Bukauskaitė, D.; Zehtindjiev, P.; Ilieva, M.; Shapoval, A.P.; Bolshakov, C.V.; et al. Plasmodium spp.: An experimental study on vertebrate host susceptibility to avian malaria. Exp. Parasitol. 2015, 148, 1–16. [Google Scholar] [CrossRef]
- Loiseau, C.; Zoorob, R.; Garnier, S.; Birard, J.; Federici, P.; Julliard, R.; Sorci, G. Antagonistic effects of a MHC class I allele on malaria-infected house sparrows. Ecol. Lett. 2008, 11, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Hasselquist, D.; Östman, Ö.; Waldenström, J.; Bensch, S. Temporal Patterns of Occurrence and Transmission of the Blood Parasite Haemoproteus Payevskyi in the Great Reed Warbler Acrocephalus Arundinaceus. J. Ornithol. 2007, 148, 401–409. [Google Scholar] [CrossRef]
- Palinauskas, V.; Valkiūnas, G.; Bolshakov, C.V.; Bensch, S. Plasmodium relictum (lineage SGS1) and Plasmodium ashfordi (lineage GRW2): The effects of the co-infection on experimentally infected passerine birds. Exp. Parasitol. 2011, 127, 527–533. [Google Scholar] [CrossRef]
- Pigeault, R.; Cozzarolo, C.-S.; Choquet, R.; Strehler, M.; Jenkins, T.; Delhaye, J.; Bovet, L.; Wassef, J.; Glaizot, O.; Christe, P. Haemosporidian infection and co-infection affect host survival and reproduction in wild populations of great tits. Int. J. Parasitol. 2018, 48, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Lapointe, D.A.; Goff, M.L.; Atkinson, C.T. Thermal Constraints to the Sporogonic Development and Altitudinal Distribution of Avian Malaria Plasmodium relictum in Hawai’i. J. Parasitol. 2010, 96, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Alarcon, D.; Palinauskas, V.; Schaefer, H.M. Diptera vectors of avian Haemosporidian parasites: untangling parasite life cycles and their taxonomy. Biol. Rev. 2012, 87, 928–964. [Google Scholar] [CrossRef]
- Zehtindjiev, P.; Ilieva, M.; Westerdahl, H.; Hansson, B.; Valkiūnas, G.; Bensch, S. Dynamics of parasitemia of malaria parasites in a naturally and experimentally infected migratory songbird, the great reed warbler Acrocephalus arundinaceus. Exp. Parasitol. 2008, 119, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Schoener, E.R.; Tompkins, D.M.; Parker, K.A.; Howe, L.; Castro, I. Presence and diversity of mixed avian Plasmodium spp. infections in introduced birds whose distribution overlapped with threatened New Zealand endemic birds. N. Z. Vet. J. 2019, 68, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Evans, M.; Otter, A. Fatal combined infection with Haemoproteus noctuae and Leucocytozoon ziemanni in juvenile snowy owls (Nyctea scandiaca). Veter-Rec. 1998, 143, 72–76. [Google Scholar] [CrossRef] [PubMed]
- De Roode, J.C.; Helinski, M.E.H.; Anwar, M.A.; Read, A.F. Dynamics of Multiple Infection and Within-Host Competition in Genetically Diverse Malaria Infections. Am. Nat. 2005, 166, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Svensson, L.; Mullarney, K.; Zetterström, D. Collins Bird Guide, 2nd ed.; British Birds; HarperCollins: London, UK, 2010. [Google Scholar]
- Bruford, M.W.; Hanotte, O.; Brookfield, J.F.Y.; Burke, T. Single-locus and multilocus DNA fingerprinting. In Molecular Genetic Analysis of Populations: A Practical Approach; Hoelzel, A.R., Ed.; IRL Press: Oxford, UK, 1992; pp. 225–269. [Google Scholar]
- Hellgren, O.; Waldenström, J.; Bensch, S. A new PCR assay for simultaneous studies of leucocytozoon, plasmodium, and haemoproteus from avian blood. J. Parasitol. 2004, 90, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]


{kind=link}
{kind=link}
| Host Family and Species | Study Site | Migration Status | Na | Parasite Species | Lineage (GenBank Accession) | Molecular Prevalence of Lineage (%) b |
|---|---|---|---|---|---|---|
| Acrocephalidae | ||||||
| Acrocephalus arundinaceusc | A, B | P | 6 | Leucocytozoon sp. | AFR214 (MT925903) | 1 (16.7) |
| Haemoproteus sp. | SW4 (MT925861) | 1 (16.7) | ||||
| Haemoproteus nucleocondensus | GRW01 (MT925867) | 1 (16.7) | ||||
| Plasmodium sp. | SYBOR10 (MT925889) | 1 (16.7) | ||||
| Acrocephalus dumetorum | G | B P | 1 | 0 | 0 | 0 |
| Acrocephalus palustris | B | B P | 4 | Haemoproteus belopolskyi | ARW1 (MT925863) | 1 (25.0) |
| Haemoproteus sp. | MW3 (MT925871) | 1 (25.0) | ||||
| Acrocephalus scirpaceus | B | B | 7 | Haemoproteus belopolskyi | ARW1 (MT925863) | 1 (14.3) |
| Haemoproteus belopolskyi | MW1 (MT925870) | 1 (14.3) | ||||
| Iduna pallida | A, B | B | 4 | Haemoproteus sp. | HIP2 (MT925849) | 2 (50.0) |
| Alaudidae | ||||||
| Calandrellabrachydactyla | G | B | 2 | Leucocytozoon sp. | WW6 (MT925894) | 1 (50.0) |
| Galerida cristata | H | R | 1 | 0 | 0 | 0 |
| Emberizidae | ||||||
| Emberiza cineracea | B | B | 1 | 0 | 0 | 0 |
| Emberiza melanocephala | A | B P | 1 | 0 | 0 | 0 |
| Estrildidae | ||||||
| Lonchura malabarica | C | R | 1 | 0 | 0 | 0 |
| Fringillidae | ||||||
| Carduelis carduelisc | H | R | 2 | Plasmodium sp. | LK05 (MT925884) | 1 (50.0) |
| Hypocoliidae | ||||||
| Hypocolius ampelinusc | A | W B | 1 | Haemoproteus passeris | PADOM05 (MT925798) | 1 (100) |
| Laniidae | ||||||
| Lanius isabellinusc | F | W B P | 1 | Haemoproteus lanii | RB1 (MT925874) | 1 (100) |
| Leiothrichidae | ||||||
| Argya caudata | A, E | R | 3 | Haemoproteus sp. | ARGCAU1 (MT925919) | 2 (66.7) |
| Locustellidae | ||||||
| Locustella luscinioides | B | P | 1 | 0 | 0 | 0 |
| Motacillidae | ||||||
| Anthus similies | C | W B | 1 | 0 | 0 | 0 |
| Motacilla alba | H | R | 1 | 0 | 0 | 0 |
| Muscicapidae | ||||||
| Cercotrichas galactotes | A | B | 2 | Plasmodium relictum | SGS1 (MT925890) | 1 (50.0) |
| Irania gutturalis | F | B | 1 | 0 | 0 | 0 |
| Luscinia megarhynchosc | C, F, G | B | 8 | Haemoproteus attenuatus | ROBIN1 (MT925844) | 2 (25.0) |
| Haemoproteus passeris | PADOM05 (MT925798) | 1 (12.5) | ||||
| Plasmodium sp. | LUME4 (MT925922) | 1 (12.5) | ||||
| Plasmodium sp. | SYBOR02 (MT925888) | 1 (12.5) | ||||
| Muscicapa striata | G | B P | 1 | Haemoproteus balmorali | SFC1 (MT925875) | 1 (100) |
| Oenanthe picatac | H | W B P | 1 | Plasmodium sp. | LK05 (MT925884) | 1 (100) |
| Phoenicurus ochrurosc | A | W B | 1 | Haemoproteus passeris | PADOM05 (MT925798) | 1 (100) |
| Nectariniidae | ||||||
| Cinnyris asiaticusc | D | R | 4 | Haemoproteus sp. | PAHIS2 (MT925825) | 1 (25.0) |
| Haemoproteus sp. | CCF2 (MT925853) | 1 (25.0) | ||||
| Oriolidae | ||||||
| Oriolus oriolus | H | P | 1 | 0 | 0 | 0 |
| Passeridae | ||||||
| Gymnoris xanthocollisc | A | B | 12 | Haemoproteus sp. | GYMXAN1 (MT925915) | 2 (16.7) |
| Haemoproteus sp. | GYMXAN2 (MT925917) | 2 (16.7) | ||||
| Haemoproteus passeris | PADOM05 (MT925798) | 4 (33.3) | ||||
| Leucocytozoon sp. | RS4 (MT925895) | 1 (8.3) | ||||
| Passer domesticusc | A,B,D,E,H | R | 80 | Haemoproteus sp. | PAHIS1 (MT925822) | 2 (2.5) |
| Haemoproteus passeris | PADOM05 (MT925798) | 15 (18.7) | ||||
| Haemoproteus sp. | PAHIS2 (MT925825) | 12 (15.0) | ||||
| Haemoproteus sp. | HIP2 (MT925849) | 2 (2.5) | ||||
| Haemoproteus sp. | PADOM34 (MT925921) | 1 (1.2) | ||||
| Haemoproteus sp. | PADOM03 (MT925868) | 2 (2.5) | ||||
| Haemoproteus sp. | CCF2 (MT925853) | 1 (1.2) | ||||
| Plasmodium relictum | SGS1 (MT925890) | 1 (1.2) | ||||
| Plasmodium relictum | GRW04 (MT925877) | 2 (2.5) | ||||
| Plasmodium sp. | PADOM02 (MT925882) | 1 (1.2) | ||||
| Leucocytozoon sp. | PADOM35 (MT925930) | 1 (1.2) | ||||
| Leucocytozoon sp. | RS4 (MT925895) | 4 (5.0) | ||||
| Passer hispaniolensis | A | W P | 1 | Haemoproteus passeris | PADOM05 (MT925798) | 1 (100) |
| Passer moabiticusc | A, B | R | 7 | Haemoproteus sp. | SW4 (MT925861) | 1 (14.3) |
| Haemoproteus sp. | PAHIS1 (MT925822) | 1 (14.3) | ||||
| Plasmodium sp. | PADOM02 (MT925882) | 1 (14.3) | ||||
| Plasmodium sp. | PBPIP1 (MT925881) | 1 (14.3) | ||||
| Passer montanus | H | R | 4 | 0 | 0 | 0 |
| Phylloscopidae | ||||||
| Phylloscopus collybita | C | W P | 2 | 0 | 0 | 0 |
| Phylloscopus trochilusc | A, B | P | 5 | Haemoproteus sp. | PAHIS2 (MT925825) | 1 (20.0) |
| Haemoproteus passeris | PADOM05 (MT925798) | 1 (20.0) | ||||
| Haemoproteus palloris | WW1 (MT925860) | 1 (20.0) | ||||
| Pycnonotidae | ||||||
| Pycnonotus leucotisc | A,D,E,F | R | 24 | Haemoproteus sp. | PYCLEU2 (MT925913) | 1 (4.2) |
| Haemoproteus sp. | CCF2 (MT925853) | 1 (4.2) | ||||
| Haemoproteus sanguinis | BUL1 (MT925876) | 1 (4.2) | ||||
| Plasmodium sp. | LUME4 (MT925922) | 2 (8.3) | ||||
| Plasmodium relictum | GRW04 (MT925877) | 1 (4.2) | ||||
| Leucocytozoon sp. | PYCLEU1 (MT925926) | 1 (4.2) | ||||
| Leucocytozoon sp. | PYCLEU3 (MT925925) | 1 (4.2) | ||||
| Leucocytozoon sp. | PYCLEU4 (MT925929) | 1 (4.2) | ||||
| Leucocytozoon sp. | PYCLEU5 (MT925931) | 1 (4.2) | ||||
| Scotocercidae | ||||||
| Scotocerca inquietac | H | R | 1 | Plasmodium sp. | LK05 (MT925884) | 1 (100) |
| Sittidae | ||||||
| Sitta tephronatac | F,G,H | R | 5 | Haemoproteus sp. | SITTEP02 (MT925909) | 1 (20.0) |
| Haemoproteus sp. | SITTEP03 (MT925910) | 2 (40.0) | ||||
| Leucocytozoon sp. | PARUS20 (MT925904) | 1 (20.0) | ||||
| Sturnidae | ||||||
| Sturnus roseus | H | B | 1 | 0 | 0 | 0 |
| Sylviidae | ||||||
| Sylvia althaeac | F,G,H | B | 9 | Haemoproteus sp. | SYHOR01 (MT925840) | 4 (44.4) |
| Haemoproteus sp. | SYLALT01 (MT925912) | 1 (11.1) | ||||
| Haemoproteus sp. | CURCUR01 (MT925872) | 1 (11.1) | ||||
| Haemoproteus sp. | LWT1 (MT925873) | 1 (11.1) | ||||
| Leucocytozoon sp. | SYBOR07 (MT925892) | 1 (11.1) | ||||
| Leucocytozoon sp. | SFC8 (MT925893) | 1 (11.1) | ||||
| Leucocytozoon sp. | SYNIS4 (MT925927) | 1 (11.1) | ||||
| Sylvia atricapillac | A | P | 7 | Haemoproteus sp. | PAHIS2 (MT925825) | 1 (14.3) |
| Haemoproteus parabelopolskyi | SYAT01 (MT925846) | 1 (14.3) | ||||
| Haemoproteus parabelopolskyi | SYAT07 (MT925847) | 2 (28.6) | ||||
| Leucocytozoon sp. | SYAT22 (MT925906) | 3 (42.8) | ||||
| Sylvia communisc | A | P | 12 | Haemoproteus sp. | SYNIS2 (MT925859) | 1 (8.3) |
| Haemoproteus sp. | CWT2 (MT925856) | 2 (16.7) | ||||
| Haemoproteus sp. | CWT3 (MT925865) | 2 (16.7) | ||||
| Haemoproteus sp. | CWT7 (MT925858) | 1 (8.3) | ||||
| Leucocytozoon sp. | RS4 (MT925895) | 3 (25.0) | ||||
| Leucocytozoon sp. | SYCON05 (MT925905) | 1 (8.3) | ||||
| Sylvia currucac | A,B,C | P | 4 | Plasmodium sp. | LK05 (MT925884) | 1 (25.0) |
| Sylvia mystacea | A | P | 1 | Plasmodium relictum | GRW04 (MT925877) | 1 (100) |
| Sylvia nisoria | A,B | P | 4 | Haemoproteus sp. | PYCLEU2 (MT925913) | 1 (25.0) |
| Leucocytozoon sp. | SYNIS4 (MT925927) | 1 (25.0) | ||||
| 20 families, 29 genera, 41 species | 8 sites | 4 migration status birds | 237 | 13 parasite species | 55 lineages | Overall prevalence 121 (51.1) |
| Host Species | Haemoproteus Lineage (GenBank Accession) | Leucocytozoon Lineage (GenBank Accession) | Na | Study Site b |
|---|---|---|---|---|
| Sylvia althaea | SYHOR01 (MT925840) | SYBOR07 (MT925892) | 1 | F |
| S. althaea | LWT1 (MT925873) | SFC8 (MT925893) | 1 | F |
| S. althaea | CURCUR01 (MT925872) | SYNIS4 (MT925927) | 1 | H |
| S. atricapilla | SYAT07 (MT925847) | SYAT22 (MT925906) | 2 | A |
| S. atricapilla | PAHIS2 (MT925825) | SYAT22 (MT925906) | 1 | A |
| S. communis | SYNIS2 (MT925859) | RS4 (MT925895) | 1 | A |
| S. communis | CWT2 (MT925856) | RS4 (MT925895) | 1 | A |
| Passer domesticus | PAHIS2 (MT925825) | RS4 (MT925895) | 1 | A |
| P. domesticus | PADOM05 (MT925798) | RS4 (MT925895) | 2 | E |
| P. domesticus | HIP2 (MT925849) | RS4 (MT925895) | 1 | E |
| Pycnonotus leucotis | BUL1 (MT925876) | PYCLEU5 (MT925931) | 1 | C |
| Total | 10 lineages | 6 lineages | 13 | 5 sites |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghaemitalab, V.; Mirshamsi, O.; Valkiūnas, G.; Aliabadian, M. Prevalence and Genetic Diversity of Avian Haemosporidian Parasites in Southern Iran. Pathogens 2021, 10, 645. https://doi.org/10.3390/pathogens10060645
Ghaemitalab V, Mirshamsi O, Valkiūnas G, Aliabadian M. Prevalence and Genetic Diversity of Avian Haemosporidian Parasites in Southern Iran. Pathogens. 2021; 10(6):645. https://doi.org/10.3390/pathogens10060645
Chicago/Turabian StyleGhaemitalab, Vajiheh, Omid Mirshamsi, Gediminas Valkiūnas, and Mansour Aliabadian. 2021. "Prevalence and Genetic Diversity of Avian Haemosporidian Parasites in Southern Iran" Pathogens 10, no. 6: 645. https://doi.org/10.3390/pathogens10060645