
Culture of Mycobacterium smegmatis in Different Carbon Sources to Induce In Vitro Cholesterol Consumption Leads to Alterations in the Host Cells after Infection: A Macrophage Proteomics Analysis


 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mycobacterial Strain and Culture Conditions
2.2. Macrophage Culture and Infection with Mycobacterium Smegmatis
2.3. Cell Lysis and Sample Preparation
2.4. NanoLC-ESI MS/MS Analysis
2.5. Data Processing and Database Searches
2.6. Functional Correlation Analysis
3. Results and Discussion
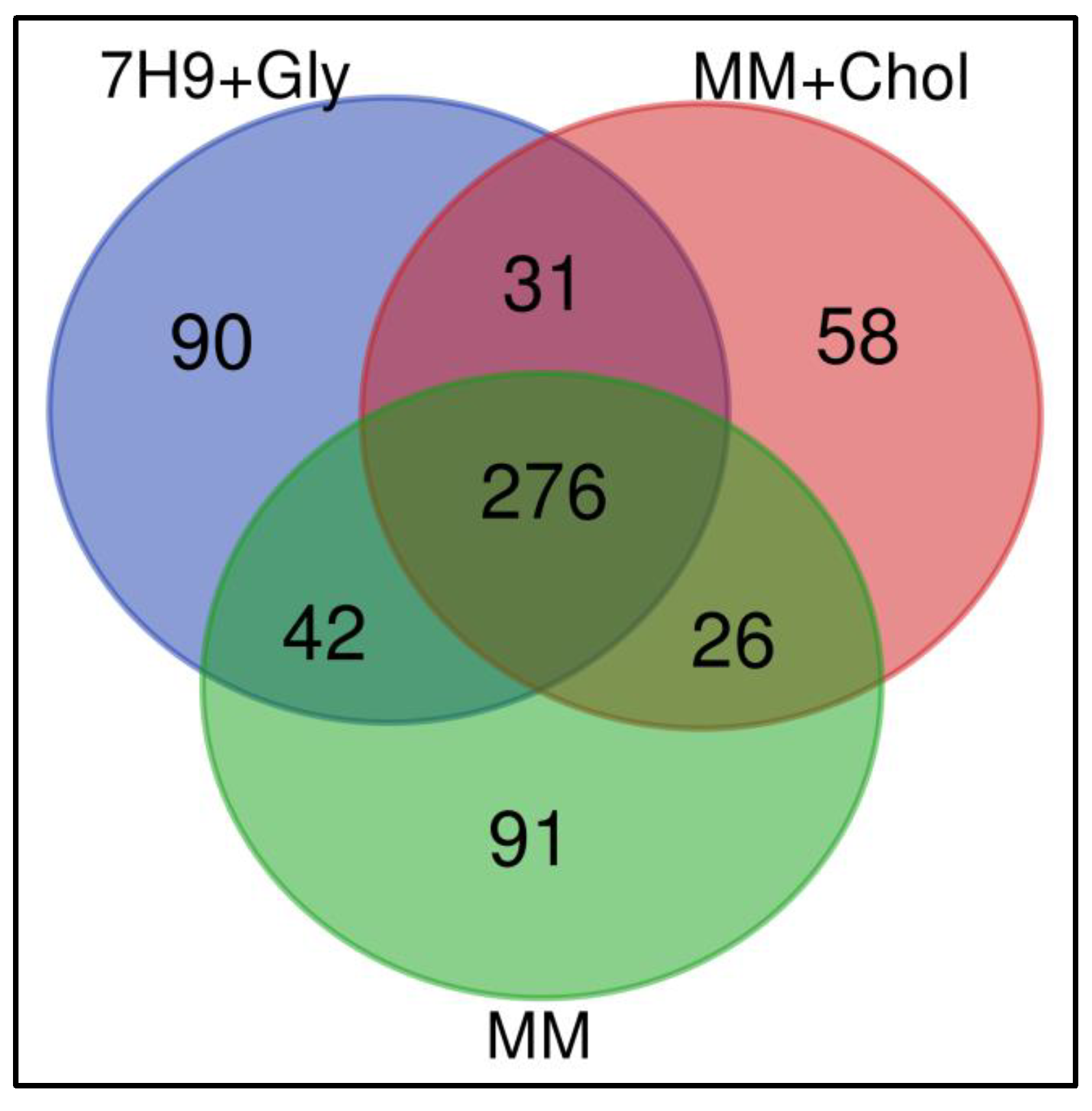
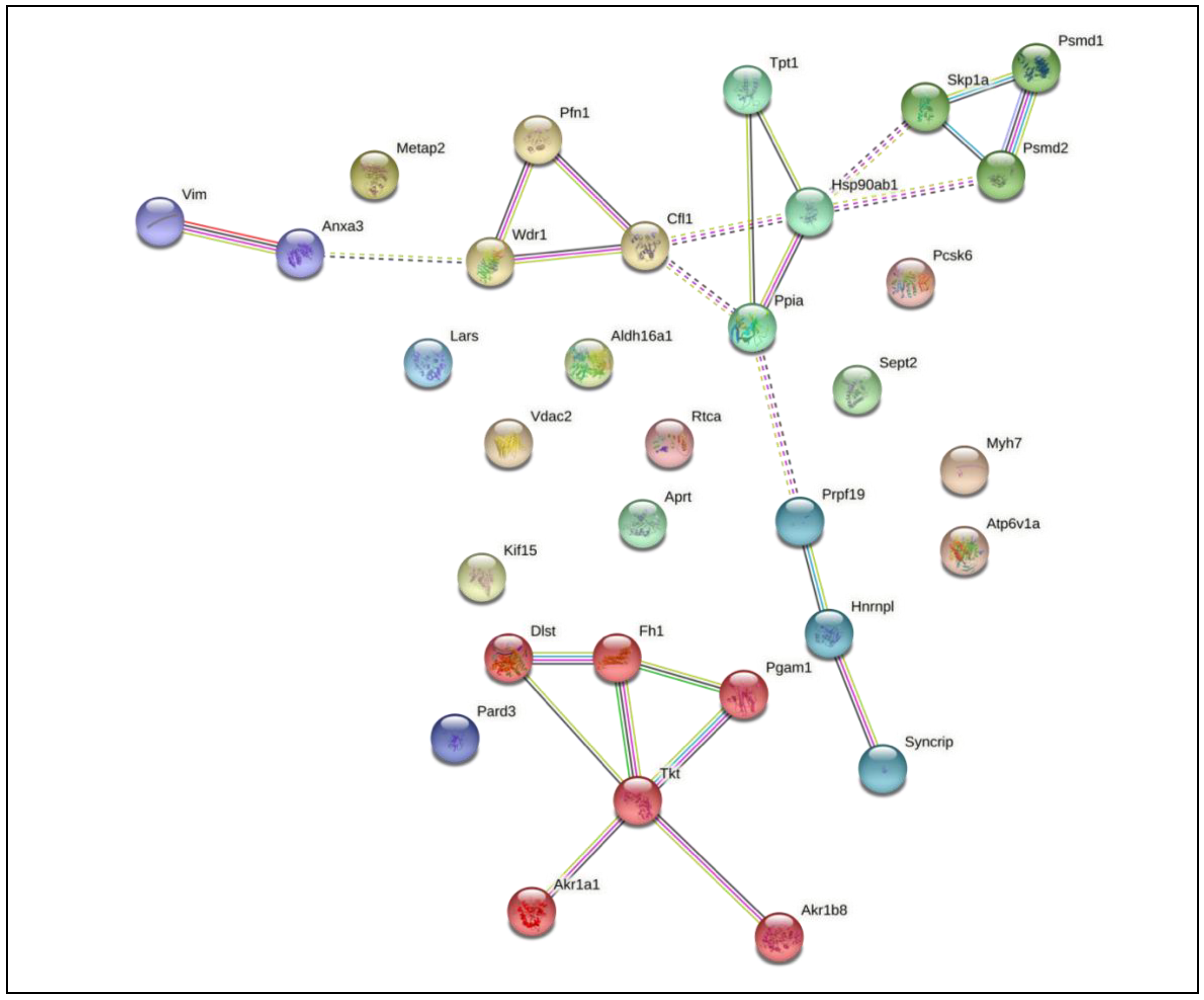
3.1. Proteomics Revealed Different Expression Profiles When Macrophages Are Infected with M. smegmatis after Cholesterol Consumption
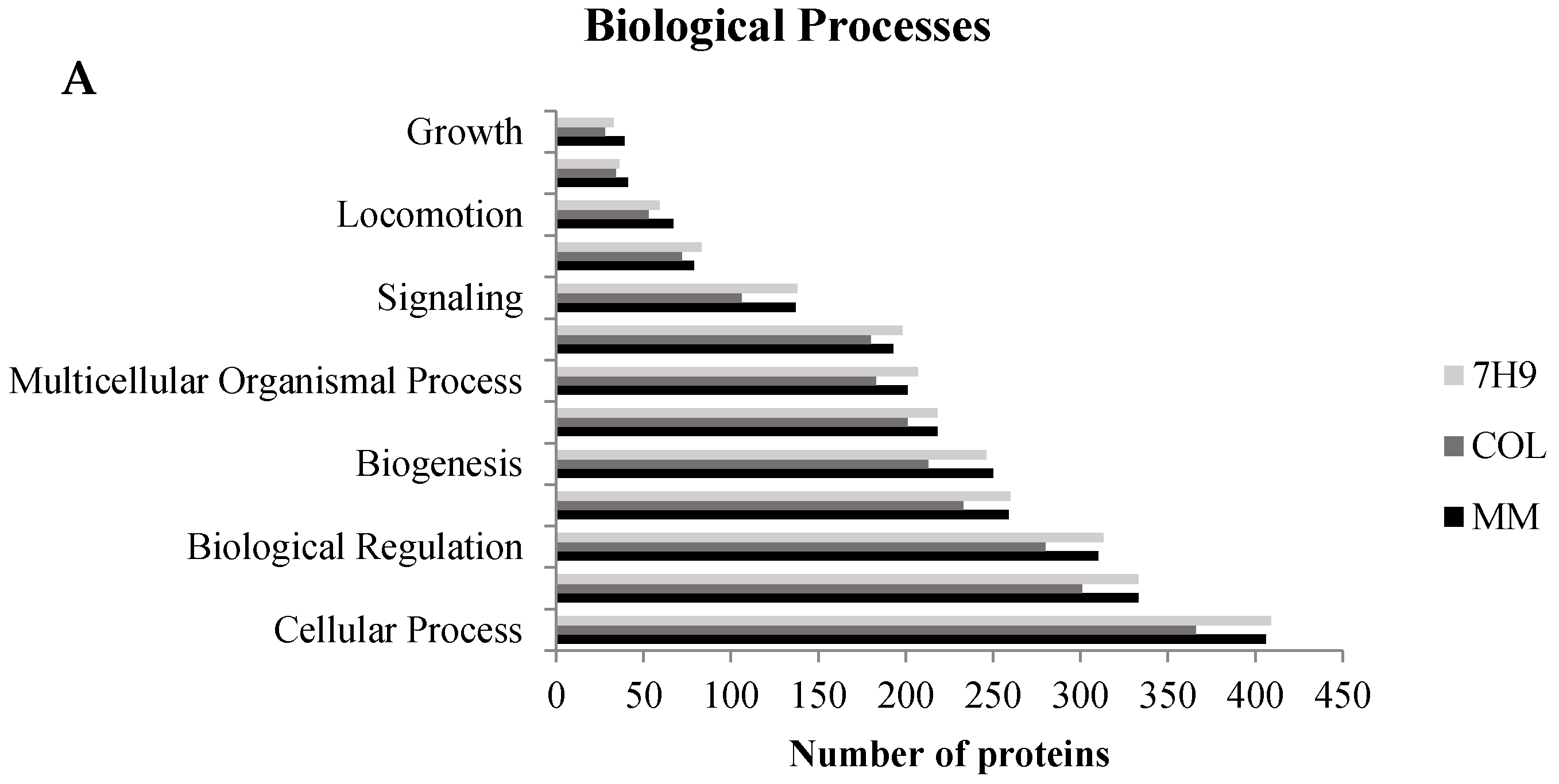
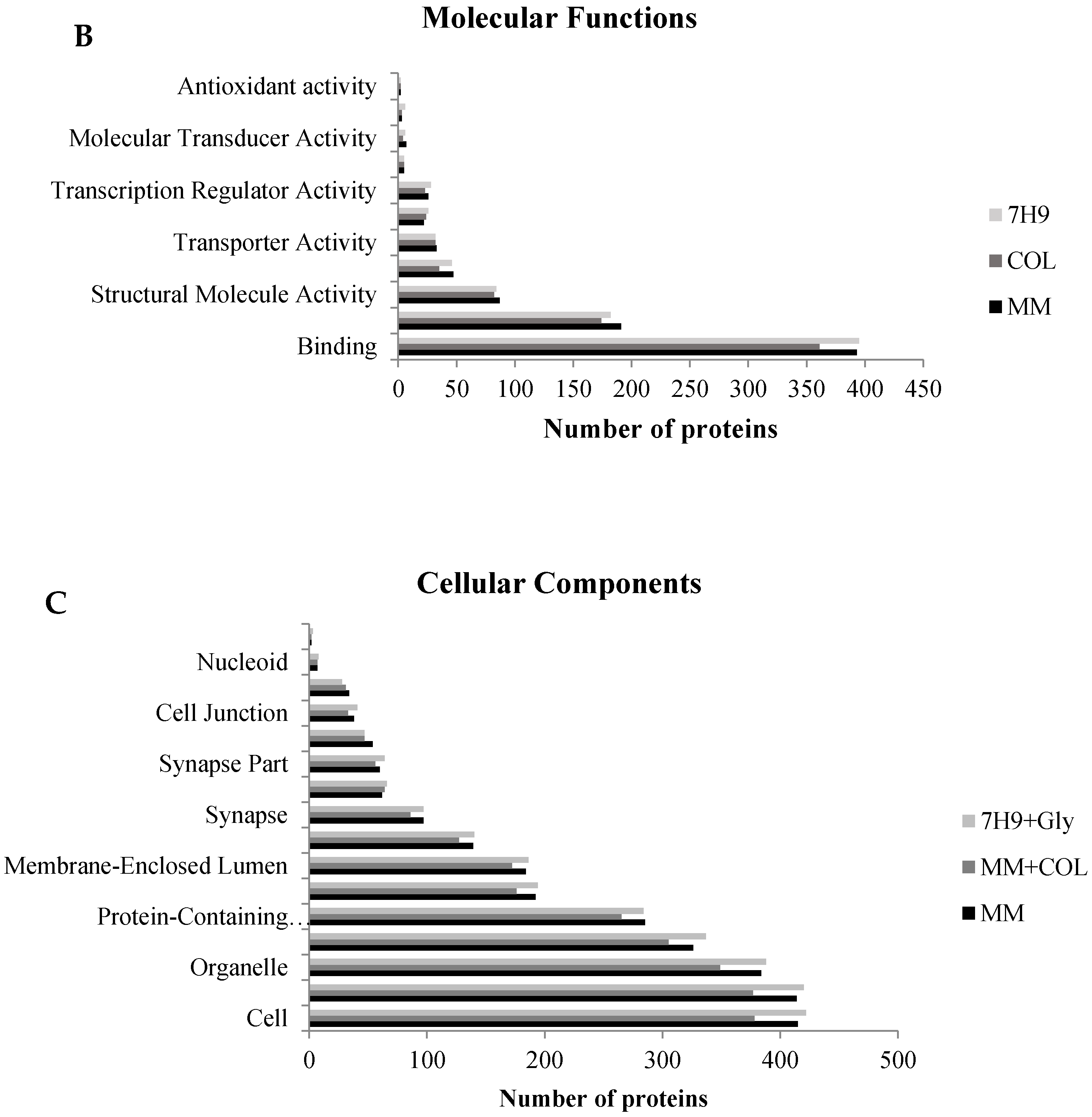
3.2. Common Effects—Immune System and Cytoskeleton
3.2.1. Cytoskeleton
3.2.2. Immune System
3.3. Exclusive Events—Specific Differences in Host Protein Regulation Induced by M. smegmatis Infection
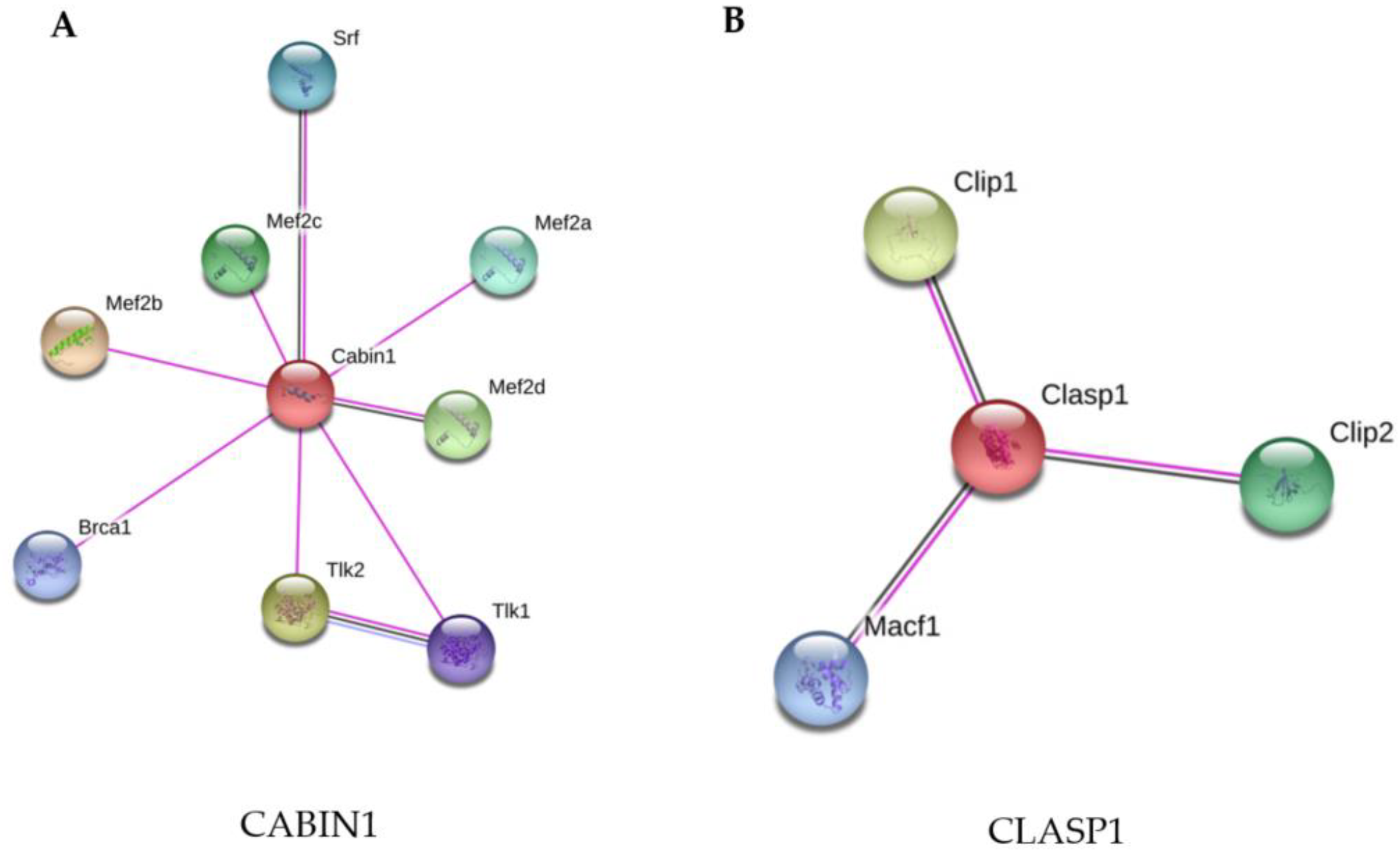
3.3.1. Infection of Macrophages by M. Smegmatis Grown in 7H9 + Gly up-Regulates CABIN1, a Negative Regulator of Calcineurin
3.3.2. Infection of Macrophages with M. Smegmatis Grown in MM Induces up-Regulation of CLASP 1, a Cytoskeleton Protein
3.3.3. Infection of Macrophages with M. smegmatis Grown in MM + Chol Induces Differential Expression of Proteins of the Immune System and Immunometabolism
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Tuberculosis Report 2020; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- VanderVen, B.C.; Huang, L.; Rohde, K.H.; Russell, D.G. The Minimal Unit of Infection: Mycobacterium tuberculosis in the Macrophage. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Hmama, Z.; Peña-Díaz, S.; Joseph, S.; Av-Gay, Y. Immunoevasion and immunosuppression of the macrophage by Mycobacterium tuberculosis. Immunol. Rev. 2015, 264, 220–232. [Google Scholar] [CrossRef]
- Russell, D.G.; Huang, L.; VanderVen, B.C. Immunometabolism at the interface between macrophages and pathogens. Nat. Rev. Immunol. 2019, 19, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.A.; Hart, P.D. Phagosome-lysosome interactions in cultured macrophages infected with virulent tubercle bacilli. Reversal of the usual nonfusion pattern and observations on bacterial survival. J. Exp. Med. 1975, 142, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Sturgill-Koszycki, S.S.; Chakraborty, P.; Haddix, P.L.; Collins, H.L.; Russell, D.G. Lack of Acidification in Mycobacterium Phagosomes Produced by Exclusion of the Vesicular Proton-ATPase. Science 1994, 263, 4. [Google Scholar] [CrossRef]
- Li, P.; Wang, R.; Dong, W.; Hu, L.; Zong, B.; Zhang, Y.; Wang, X.; Guo, A.; Zhang, A.; Xiang, Y.; et al. Comparative Proteomics Analysis of Human Macrophages Infected with Virulent Mycobacterium bovis. Front. Cell. Infect. Microbiol. 2017, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Haglund, C.M.; Welch, M.D. Pathogens and polymers: Microbe–host interactions illuminate the cytoskeleton. J. Cell Biol. 2011, 195, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Mishra, M.; Dadhich, R.; Mogha, P.; Kapoor, S. Mycobacterium Lipids Modulate Host Cell Membrane Mechanics, Lipid Diffusivity, and Cytoskeleton in a Virulence-Selective Manner. ACS Infect. Dis. 2020, 6, 2386–2399. [Google Scholar] [CrossRef]
- He, Y.; Li, W.; Liao, G.; Xie, J. Mycobacterium tuberculosis -Specific Phagosome Proteome and Underlying Signaling Pathways. J. Proteome Res. 2012, 11, 2635–2643. [Google Scholar] [CrossRef]
- Anes, E.; Peyron, P.; Staali, L.; Jordao, L.; Gutierrez, M.G.; Kress, H.; Hagedorn, M.; Maridonneau-Parini, I.; Skinner, M.A.; Wildeman, A.G.; et al. Dynamic life and death interactions between Mycobacterium smegmatis and J774 macrophages. Cell. Microbiol. 2006, 8, 939–960. [Google Scholar] [CrossRef]
- Jordao, L.; Bleck, C.K.E.; Mayorga, L.; Griffiths, G.; Anes, E. On the killing of mycobacteria by macrophages. Cell. Microbiol. 2007, 10, 529–548. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Sassetti, C.M. Mycobacterial persistence requires the utilization of host cholesterol. Proc. Natl. Acad. Sci. USA 2008, 105, 4376–4380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, J.E.; Pandey, A.K.; Gilmore, S.A.; Mizrahi, V.; McKinney, J.D.; Bertozzi, C.R.; Sassetti, C.M. Cholesterol Catabolism by Mycobacterium tuberculosis Requires Transcriptional and Metabolic Adaptations. Chem. Biol. 2012, 19, 218–227. [Google Scholar] [CrossRef] [Green Version]
- Lovewell, R.R.; Sassetti, C.M.; VanderVen, B.C. Chewing the fat: Lipid metabolism and homeostasis during M. tuberculosis infection. Curr. Opin. Microbiol. 2016, 29, 30–36. [Google Scholar] [CrossRef]
- Dos Santos, A.C.D.; Marinho, V.H.D.S.; Silva, P.H.D.A.; Macchi, B.D.M.; Arruda, M.S.P.; Da Silva, E.O.; Nascimento, J.L.M.D.; De Sena, C.B.C. Microenvironment of Mycobacterium smegmatis Culture to Induce Cholesterol Consumption Does Cell Wall Remodeling and Enables the Formation of Granuloma-Like Structures. BioMed Res. Int. 2019, 2019, 1–13. [Google Scholar] [CrossRef]
- Brzostek, A.; Pawelczyk, J.; Rumijowska-Galewicz, A.; Dziadek, B.; Dziadek, J. Mycobacterium tuberculosis Is Able To Accumulate and Utilize Cholesterol. J. Bacteriol. 2009, 191, 6584–6591. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wei, S.; Fang, Y.; Li, M.; Li, X.; Li, Z.; Zhang, J.; Zhu, G.; Li, C.; Bi, L.; et al. Quantitative proteomic analysis of host responses triggered by Mycobacterium tuberculosis infection in human macrophage cells. Acta Biochim. Biophys. Sin. 2017, 49, 835–844. [Google Scholar] [CrossRef] [Green Version]
- Shui, W.; Gilmore, S.A.; Sheu, L.; Liu, J.; Keasling, J.D.; Bertozzi, C.R. Quantitative Proteomic Profiling of Host−Pathogen Interactions: The Macrophage Response to Mycobacterium tuberculosis Lipids. J. Proteome Res. 2009, 8, 282–289. [Google Scholar] [CrossRef] [Green Version]
- Shui, W.; Petzold, C.J.; Redding, A.; Liu, J.; Pitcher, A.; Sheu, L.; Hsieh, T.-Y.; Keasling, J.D.; Bertozzi, C.R. Organelle Membrane Proteomics Reveals Differential Influence of Mycobacterial Lipoglycans on Macrophage Phagosome Maturation and Autophagosome Accumulation. J. Proteome Res. 2011, 10, 339–348. [Google Scholar] [CrossRef]
- Diaz, G.; Wolfe, L.M.; Kruh-Garcia, N.A.; Dobos, K.M. Changes in the Membrane-Associated Proteins of Exosomes Released from Human Macrophages after Mycobacterium tuberculosis Infection. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Menon, D.; Singh, K.; Pinto, S.M.; Nandy, A.; Jaisinghani, N.; Kutum, R.; Dash, D.; Prasad, T.S.K.; Gandotra, S. Quantitative Lipid Droplet Proteomics Reveals Mycobacterium tuberculosis Induced Alterations in Macrophage Response to Infection. ACS Infect. Dis. 2019, 5, 559–569. [Google Scholar] [CrossRef] [Green Version]
- Sena, C.B.C.; Fukuda, T.; Miyanagi, K.; Matsumoto, S.; Kobayashi, K.; Murakami, Y.; Maeda, Y.; Kinoshita, T.; Morita, Y.S. Controlled expression of branch-forming mannosyltransferase is critical for mycobacterial lipoarabinomannan biosynthesis. J. Biol. Chem. 2010, 285, 13326–13336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassoli, J.S.; Martins-De-Souza, D. Comprehensive Shotgun Proteomic Analyses of Oligodendrocytes Using Ion Mobility and Data-Independent Acquisition; Springer Nature Experiments, 2017; Volume 127, Available online: https://experiments.springernature.com/articles/10.1007/978-1-4939-7119-0_5 (accessed on 20 July 2020).
- Nascimento, S.V.D.; Magalhães, M.M.; Cunha, R.L.; Costa, P.H.D.O.; Alves, R.C.D.O.; De Oliveira, G.C.; Valadares, R.B.D.S. Differential accumulation of proteins in oil palms affected by fatal yellowing disease. PLoS ONE 2018, 13, e0195538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.-Z.; Vissers, J.P.C.; Silva, J.C.; Golick, D.; Gorenstein, M.V.; Geromanos, S.J. Database searching and accounting of multiplexed precursor and product ion spectra from the data independent analysis of simple and complex peptide mixtures. Proteomics 2009, 9, 1696–1719. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, E.W.; Bandeira, N.; Sharma, V.; Perez-Riverol, Y.; Carver, J.J.; Kundu, D.J.; García-Seisdedos, D.; Jarnuczak, A.F.; Hewapathirana, S.; Pullman, B.S.; et al. The ProteomeXchange consortium in 2020: Enabling ‘big data’ approaches in proteomics. Nucleic Acids Res. 2020, 48, 1145–1152. [Google Scholar] [CrossRef] [Green Version]
- Perez-Riverol, Y.; Xu, Q.-W.; Wang, R.; Uszkoreit, J.; Griss, J.; Sanchez, A.; Reisinger, F.; Csordas, A.; Ternent, T.; Del-Toro, N.; et al. PRIDE Inspector Toolsuite: Moving Toward a Universal Visualization Tool for Proteomics Data Standard Formats and Quality Assessment of ProteomeXchange Datasets. Mol. Cell. Proteom. 2016, 15, 305–317. [Google Scholar] [CrossRef] [Green Version]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]
- Rosentreter, A.; Hofmann, A.; Xavier, C.-P.; Stumpf, M.; Noegel, A.A.; Clemen, C.S. Coronin 3 involvement in F-actin-dependent processes at the cell cortex. Exp. Cell Res. 2007, 313, 878–895. [Google Scholar] [CrossRef] [Green Version]
- Rybakin, V.; Clemen, C.S. Coronin proteins as multifunctional regulators of the cytoskeleton and membrane trafficking. Bioessays 2005, 27, 625–632. [Google Scholar] [CrossRef]
- Hoyer, M.J.; Chitwood, P.J.; Ebmeier, C.C.; Striepen, J.F.; Qi, R.Z.; Old, W.M.; Voeltz, G.K. A Novel Class of ER Membrane Proteins Regulates ER-Associated Endosome Fission. Cell 2018, 175, 254–265. [Google Scholar] [CrossRef] [Green Version]
- Behrens, J.; Solga, R.; Ziemann, A.; Rastetter, R.H.; Berwanger, C.; Herrmann, H.; Noegel, A.A.; Clemen, C.S. Coronin 1C-free primary mouse fibroblasts exhibit robust rearrangements in the orientation of actin filaments, microtubules and intermediate filaments. Eur. J. Cell Biol. 2016, 95, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Pieters, J.; Müller, P.; Jayachandran, R. On guard: Coronin proteins in innate and adaptive immunity. Nat. Rev. Immunol. 2013, 13, 1–9. [Google Scholar] [CrossRef]
- Mori, M.; Mode, R.; Pieters, J. From Phagocytes to Immune Defense: Roles for Coronin Proteins in Dictyostelium and Mammalian Immunity. Front. Cell. Infect. Microbiol. 2018, 8, 77. [Google Scholar] [CrossRef]
- Reymann, A.-C.; Boujemaa-Paterski, R.; Martiel, J.-L.; Guérin, C.; Cao, W.; Chin, H.F.; De La Cruz, E.M.; Théry, M.; Blanchoin, L. Actin Network Architecture Can Determine Myosin Motor Activity. Science 2012, 336, 1310–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luxenburg, C.; Heller, E.; Pasolli, H.A.; Chai, S.; Nikolova, M.; Stokes, N.; Fuchs, E. Wdr1-mediated cell shape dynamics and cortical tension are essential for epidermal planar cell polarity. Nat. Cell Biol. 2015, 17, 592–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rius, C.; Aller, P. Vimentin expression as a late event in the in vitro differentiation of human promonocytic cells. J. Cell Sci. 1992, 101, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Beneš, P.; Macečková, V.; Zdráhal, Z.; Konečná, H.; Zahradníčková, E.; Mužík, J.; Šmarda, J. Role of vimentin in regulation of monocyte/macrophage differentiation. Differentiation 2006, 74, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Mor-Vaknin, N.; Punturieri, A.; Sitwala, K.; Markovitz, D.M. Vimentin is secreted by activated macrophages. Nat. Cell Biol. 2003, 5, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Mahesh, P.P.; Retnakumar, R.J.; Mundayoor, S. Downregulation of vimentin in macrophages infected with live Mycobacterium tuberculosis is mediated by Reactive Oxygen Species. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hare, N.J.; Chan, B.; Chan, E.; Kaufman, K.L.; Britton, W.J.; Saunders, B.M. Microparticles released from Mycobacterium tuberculosis-infected human macrophages contain increased levels of the type I interferon inducible proteins including ISG15. Proteomics 2015, 15, 3020–3029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonilla, D.L.; Bhattacharya, A.; Sha, Y.; Xu, Y.; Xiang, Q.; Kan, A.; Jagannath, C.; Komatsu, M.; Eissa, N.T. Autophagy Regulates Phagocytosis by Modulating the Expression of Scavenger Receptors. Immunity 2013, 39, 537–547. [Google Scholar] [CrossRef] [Green Version]
- Bjørkøy, G.; Lamark, T.; Johansen, T. p62/SQSTM1: A Missing Link between Protein Aggregates and the Autophagy Machinery. Autophagy 2006, 2, 138–139. [Google Scholar] [CrossRef] [Green Version]
- Wee, Y.S.; Roundy, K.M.; Weis, J.J.; Weis, J.H. Interferon-inducible transmembrane proteins of the innate immune response act as membrane organizers by influencing clathrin and v-ATPase localization and function. Innate Immun. 2012, 18, 834–845. [Google Scholar] [CrossRef]
- Ranjbar, S.; Haridas, V.; Jasenosky, L.D.; Falvo, J.V.; Goldfeld, A.E. A Role for IFITM Proteins in Restriction of Mycobacterium tuberculosis Infection. Cell Rep. 2015, 13, 874–883. [Google Scholar] [CrossRef] [Green Version]
- Wong, D.; Bach, H.; Sun, J.; Hmama, Z.; Av-Gay, Y. Mycobacterium tuberculosis protein tyrosine phosphatase (PtpA) excludes host vacuolar-H+-ATPase to inhibit phagosome acidification. Proc. Natl. Acad. Sci. USA 2011, 108, 19371–19376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, D.; Li, W.; Chao, J.D.; Zhou, P.; Narula, G.; Tsui, C.; Ko, M.; Xie, J.; Martinez-Frailes, C.; Av-Gay, Y. Protein tyrosine kinase, PtkA, is required for Mycobacterium tuberculosis growth in macrophages. Sci. Rep. 2018, 8, 155. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Meena, L.S. Potential of Ca2+ in Mycobacterium tuberculosis H37Rv Pathogenesis and Survival. Appl. Biochem. Biotechnol. 2016, 181, 762–771. [Google Scholar] [CrossRef] [PubMed]
- Meena, L.S. Interrelation of Ca2+ and PE_PGRS proteins during Mycobacterium tuberculosis pathogenesis. J. Biosci. 2019, 44, 24. [Google Scholar] [CrossRef]
- Zhao, X.; Kumar, P.; Shah-Simpson, S.; CaraDonna, K.L.; Galjart, N.; Teygong, C.; Blader, I.; Wittmann, T.; Burleigh, B.A. Host microtubule plus-end binding protein CLASP1 influences sequential steps in the Trypanosoma cruzi infection process: Microtubule plus-end binding proteins facilitate intracellular trypanosome infection. Cell. Microbiol. 2013, 15, 571–584. [Google Scholar] [CrossRef] [Green Version]
- Patel, P.C.; Fisher, K.H.; Yang, E.C.C.; Harrison, R.E. Proteomic Analysis of Microtubule-associated Proteins during Macrophage Activation. Mol. Cell. Proteom. 2009, 8, 2500–2514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shekhawat, S.D.; Purohit, H.J.; Taori, G.M.; Daginawala, H.F.; Kashyap, R.S. Evaluation of heat shock proteins for discriminating between latent tuberculosis infection and active tuberculosis: A preliminary report. J. Infect. Public Health 2016, 9, 143–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Singh, P.; Kolloli, A.; Shi, L.; Bushkin, Y.; Tyagi, S.; Subbian, S. Immunometabolism of Phagocytes During Mycobacterium tuberculosis Infection. Front. Mol. Biosci. 2019, 6, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Eugenin, E.A.; Subbian, S. Immunometabolism in Tuberculosis. Front. Immunol. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Jiang, Q.; Bushkin, Y.; Subbian, S.; Tyagi, S. Biphasic Dynamics of Macrophage Immunometabolism during Mycobacterium tuberculosis Infection. mBio 2019, 10, e02550-18. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Salamon, H.; Eugenin, E.A.; Pine, R.; Cooper, A.; Gennaro, M.L. Infection with Mycobacterium tuberculosis induces the Warburg effect in mouse lungs. Sci. Rep. 2016, 5, 18176. [Google Scholar] [CrossRef] [PubMed]
- Cumming, B.M.; Addicott, K.W.; Adamson, J.H.; Steyn, A.J. Mycobacterium tuberculosis induces decelerated bioenergetic metabolism in human macrophages. eLife 2018, 7, 1–18. [Google Scholar] [CrossRef]
- Radulovic, M.; Godovac-Zimmermann, J. Proteomic approaches to understanding the role of the cytoskeleton in host-defense mechanisms. Expert Rev. Proteom. 2011, 8, 117–126. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Groups | Identified Proteins | Quantified Proteins | Exclusive Proteins | Regulated Proteins | |
|---|---|---|---|---|---|
| Up- | Down- | ||||
| 7H9 + Gly | 439 | 119 | 90 | 5 | 2 |
| MM + Chol | 391 | 139 | 58 | 7 | 26 |
| MM | 435 | 90 | 91 | 4 | 0 |
| Total | 1265 | 44 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Lima, J.B.; da Silva Fonseca, L.P.; Xavier, L.P.; de Matos Macchi, B.; Cassoli, J.S.; da Silva, E.O.; da Silva Valadares, R.B.; do Nascimento, J.L.M.; Santos, A.V.; de Sena, C.B.C. Culture of Mycobacterium smegmatis in Different Carbon Sources to Induce In Vitro Cholesterol Consumption Leads to Alterations in the Host Cells after Infection: A Macrophage Proteomics Analysis. Pathogens 2021, 10, 662. https://doi.org/10.3390/pathogens10060662
de Lima JB, da Silva Fonseca LP, Xavier LP, de Matos Macchi B, Cassoli JS, da Silva EO, da Silva Valadares RB, do Nascimento JLM, Santos AV, de Sena CBC. Culture of Mycobacterium smegmatis in Different Carbon Sources to Induce In Vitro Cholesterol Consumption Leads to Alterations in the Host Cells after Infection: A Macrophage Proteomics Analysis. Pathogens. 2021; 10(6):662. https://doi.org/10.3390/pathogens10060662
Chicago/Turabian Stylede Lima, Jaqueline Batista, Lana Patricia da Silva Fonseca, Luciana Pereira Xavier, Barbarella de Matos Macchi, Juliana Silva Cassoli, Edilene Oliveira da Silva, Rafael Borges da Silva Valadares, José Luiz Martins do Nascimento, Agenor Valadares Santos, and Chubert Bernardo Castro de Sena. 2021. "Culture of Mycobacterium smegmatis in Different Carbon Sources to Induce In Vitro Cholesterol Consumption Leads to Alterations in the Host Cells after Infection: A Macrophage Proteomics Analysis" Pathogens 10, no. 6: 662. https://doi.org/10.3390/pathogens10060662
APA Stylede Lima, J. B., da Silva Fonseca, L. P., Xavier, L. P., de Matos Macchi, B., Cassoli, J. S., da Silva, E. O., da Silva Valadares, R. B., do Nascimento, J. L. M., Santos, A. V., & de Sena, C. B. C. (2021). Culture of Mycobacterium smegmatis in Different Carbon Sources to Induce In Vitro Cholesterol Consumption Leads to Alterations in the Host Cells after Infection: A Macrophage Proteomics Analysis. Pathogens, 10(6), 662. https://doi.org/10.3390/pathogens10060662