1. Introduction
In December 2019, the first cases of atypical pneumonia associated with infection by a novel coronavirus (CoV) were reported in China [
1]. Sequence analysis of the new CoV revealed a close phylogenetic relationship to SARS-CoV (Severe Acute Respiratory Syndrome Coronavirus)-like bat CoV strains [
2,
3]. Subsequently, the new CoV was officially named SARS-CoV-2 [
4].
In the past two decades, two other coronaviruses have emerged as novel human pathogens. The first SARS-CoV emerged in 2002 and ultimately resulted in more than 8000 confirmed cases, with an apparent 10% case–fatality ratio. Fortunately, SARS-CoV transmission was effectively controlled by appropriate international responses involving quarantine measures and it was last reported in April 2004 [
5]. In 2012, another CoV emerged in Saudi Arabia, named Middle East Respiratory Syndrome (MERS)-CoV. MERS-CoV causes sporadic cases of a severe respiratory syndrome and outbreaks in healthcare settings [
6]. Both viruses are suspected to originate from bats and to have acquired adaptations that allow transmission to other species [
7]. The natural host of the new coronavirus, SARS-CoV-2, is yet to be identified, but the virus sequence is very similar to a SARS-CoV-like virus found in bats (bat coronavirus RaTG13) and pangolins [
8,
9,
10]. The above-mentioned examples underscore the remarkable plasticity of coronaviruses and their ability to easily switch between hosts. Thus, different animal species may play a role in future coronavirus outbreaks and also can be used as suitable models for studying the infection, transmission and pathogenesis of the corresponding viruses.
Infection of humans with SARS-CoV-2 causes Coronavirus Disease 19 (COVID-19). The clinical symptoms of COVID-19 can vary substantially, from mild respiratory disease to acute respiratory distress syndrome (ARDS) and death. SARS-CoV-2 spread globally within one year, resulting in a pandemic with 181.3 million confirmed cases and 3.9 million deaths (Source: John Hopkins University and Medicine website, as of 28 June 2021). As soon as it became clear that the new coronavirus was rapidly spreading and would not be easily contained, a global effort in the scientific community arose to find suitable animal models for studying the pathogenesis and transmission of SARS-CoV-2. Diverse in silico predictions of species susceptibility were published [
11,
12,
13,
14,
15]. Based on in silico modeling of interactions between the receptor for the virus, angiotensin converting enzyme 2 (ACE2) and the viral receptor-binding domain (RBD), these studies aimed at identifying animal species that are susceptible to SARS-CoV-2 infection. In parallel, many labs worldwide started exploring different animal species in the search for those that best reproduce the moderate to severe human disease. A starting point for many of these studies was results obtained previously with SARS-CoV, since this virus utilizes the same receptor, ACE2.
To tackle the initial lack of animal models, to minimize the ethical concerns of using irrelevant animal species and to reduce the number of animals used for the development of models, we evaluated the use of precision-cut lung slices (PCLS) as an ex vivo model to predict the susceptibility of different animal species to infection with SARS-CoV-2. In PCLS, the lung 3D architecture and physiology are largely preserved, and disease pathogenesis at least partly can be mimicked, rendering this system suitable for studying chronic inflammatory conditions such as asthma, chronic obstructive pulmonary disease (COPD) and idiopathic pulmonary fibrosis, but also for toxicological studies and infectious diseases (for a recent review, see [
16]). Although the PCLS can be cultivated for only relatively short periods of time (up to 14 days), infection of PCLS can be reliably quantified by PCR or visualized by immunohistochemistry (IHC) and/or immunofluorescence (IF). For preparation of PCLS, animals still need to be sacrificed, but several experiments can be performed with samples collected from only one donor, and multiple variables can be tested concurrently, as opposed to in vivo experiments. Moreover, the donor animals are euthanized without any discomfort related to infection and disease.
In this study, we evaluated the susceptibility of PCLS derived from pigs, hamsters and cats to infection with a Dutch SARS-CoV-2 isolate. Virus replication was assessed by PCR and immunofluorescence (IF). The results obtained from experiments with PCLS demonstrate that this ex vivo model can be successfully employed to predict the susceptibility of different animal species to SARS-CoV-2 and possibly other betacoronaviruses. Furthermore, we verified the susceptibility of hamsters from the same genetic background as used for the PCLS preparations. We infected adult and juvenile hamsters to look for possible age-related clinical differences, as seen in humans. Next to using well-defined parameters to establish the infection progress, we enriched and refined the hamster model by adding measurements of the individual animal activity by an activity wheel and demonstrated that daily activity counts can be employed as a clinical readout parameter of animal wellbeing post-challenge infection. Furthermore, we propose a scoring system for the quantification of lung pathology.
3. Discussion
Here, we report the evaluation of PCLS as an ex vivo system that can be used to predict the susceptibility of different animal species to SARS-CoV-2. This approach can be used for other coronaviruses or other newly emerged viruses, when suitable animal models need to be established. Furthermore, we assessed the susceptibility of juvenile versus adult Syrian hamsters using the same SARS-CoV-2 strain and hamster breed as used in the PCLS experiments.
The experiments with PCLS derived from pigs showed no evidence of SARS-CoV-2 replication despite the fact that cell lines of pig origin (swine testicle (ST) and porcine kidney (PK-15)) were previously shown to be susceptible to SARS-CoV-2 infection [
19]. Our results are, however, fully in agreement with in vivo infection experiments that showed that pigs do not support SARS-CoV-2 replication [
19,
20,
21,
22] and only occasionally seroconvert [
22,
23]. A recent report showed that SARS-CoV-2 was recovered from the submandibular lymph node of one pig 13 days after inoculation, and viral RNA was detected in nasal washes of 2 out of 16 pigs and in the oral fluid of 1 out of 16 pigs on DPI 3 [
23]. Furthermore, specific antibodies were detected in the serum of 2 out of 16 pigs and in the oral fluid of one pig. Nevertheless, these so far rare events are still in line with the overall very low susceptibility of pigs to infection with SARS-CoV-2 and with our results of PCLS infection.
In addition to PCLS from pigs, we also incubated PCLS derived from domestic cats with SARS-CoV-2. The virus efficiently replicated in cat pulmonary tissue, probably even more efficiently compared to hamsters, although it should be noted that the data were obtained from a single donor due to the limited accessibility of cats. The cat PCLS results are in agreement with experimental findings by various groups [
21,
24,
25,
26]. Interestingly, viral antigen was detected in alveoli in cat PCLS, whereas in in vivo-infected cats, only the acinar glands seemed to support virus replication [
24]. We speculate that the buffer/agarose gel of the ex vivo system might have influenced the viral attachment. Another speculation is that the alveolar walls are sensitive also in vivo, but that the limited viral replication in the large bronchi in vivo [
21,
24] hampers the virus infection to affect the deeper airways. Finally, PCLS are deprived of immune cell infiltrates, which might modify the dynamics of virus replication in vivo in the cat lungs.
In addition to efficient replication in cat PCLS, SARS-CoV-2 also efficiently replicated in PCLS derived from hamsters. Virus was detected in the alveolar and bronchiolar epithelium, similarly to in vivo infected hamsters, where the virus was predominantly found in the same type of cells as observed by our group and others [
27,
28,
29].
Hamsters are widely accepted as a good model for SARS-CoV-2 infection, recapitulating mild to moderate human disease, prominent lung pathology, efficient virus replication and seroconversion. Our in vivo data obtained with the Lelystad strain of SARS-CoV-2 are in good agreement with the findings of other groups with various strains [
27,
28,
29,
30,
31]. As a clinical readout, next to the frequently used body weight loss, here, we demonstrate that activity (measured by a wheel with a rotation counter) can be used as a very sensitive parameter of discomfort. Both body weight loss and activity measurements were sufficiently robust to be reliably used as endpoints of disease. Other clinical parameters seem less reliable. Clinical signs, although present, are mild and therefore clinical scoring is sensitive to subjectivity. Body temperature did not increase post-challenge. In contrast, a clear temperature decrease was observed between DPI 2 and 5. Hypothermia could be used as an additional readout parameter, measured together with activity and body weight. The lowest body temperatures measured post-inoculation coincided with the highest decrease in activity. However, since a surgical procedure is required to implant the temperature transponders into the abdominal cavity, it is questionable whether the temperature measurement adds sufficient value to justify the discomfort resulting from the surgical procedure of implantation.
Lung pathology and lung IHC were most prominent between DPI 4 and 7 and therefore a necropsy at any DPI between 4 and 7 should give sufficient information about lung pathology or lack thereof. Thus, timely necropsy should be applied to reduce animal discomfort when this does not interfere with the interpretation of the experimental results. To facilitate a more accurate evaluation of lung pathology, as suggested and described previously [
32], we present a detailed scoring scheme that includes several parameters for quantification of lung pathology. Depending on the timepoint of necropsy after infection, one or more of these pathological parameters can be used.
SARS-CoV-2 was found to replicate exclusively in the respiratory organs (mainly conchae and lungs). Determination of viral loads in these organs by both RT-qPCR and virus isolation is robust and gives clear readout parameters. As an alternative sample for the detection of replication in the upper respiratory tract, we used oropharyngeal swabs. Collecting these samples is less invasive than nasal washes (nasal washes require anesthesia) and can therefore be used as a refinement. Virus isolation from respiratory organs was successful only until DPI 4. Virus could not be isolated from the swabs, but subgenomic RNA PCR was positive, suggesting the presence of replicating virus in cells collected during the swabbing process. Finally, hamsters seroconverted early after inoculation and the presence and/or boost of antibody titers may thus be used to evaluate vaccine efficacy.
As part of this study, we also assessed the susceptibility of juvenile and adult hamsters to infection with SARS-CoV-2. Adult hamsters were used in an attempt to mimic the more severe symptoms most commonly reported in elderly humans after infection with SARS-CoV-2 [
33,
34]. Although the body weights of adult hamsters did not recover as quickly as those of juvenile hamsters, we did not observe differences in the lowest body weight between the two age groups as a result of challenge infection. The hypothermia in the adult hamsters post-infection seemed more pronounced than in the juvenile hamsters. However, the average baseline body temperature of the adult hamsters was also lower. When accounting for the difference in the baseline temperatures before challenge, the temperature reduction was not significantly different between adult and juvenile hamsters. In terms of pathology, the differences were limited to the progression of the lesions, which advanced and subsequently resolved slightly faster in juvenile hamsters. Our data also suggest that the virus may replicate slightly better in the conchae of adult hamsters and in the lungs of juvenile hamsters. It has to be noted that the statistical analysis was performed on a very limited number of samples per timepoint (N = 2 for DPI 0–10). Therefore, generalization of these observations for age-related differences should be performed with caution. Our main conclusion is that the differences between adult and juvenile hamsters are not substantial and that there is no advantage of using adult over juvenile animals in this model for SARS-CoV-2 infection. No differences between young and older hamsters have been reported by others either [
29,
35].
In conclusion, with this work, we demonstrate that PCLS can be used as a convenient substitute for the testing of animal susceptibility towards SARS-CoV-2. This approach can also be applied for other newly emerged viruses, for which good animal models are lacking. Multiple PCLS from different species can be prepared and tested before animals are used in subsequent experiments to establish animal models. By using this method, a relatively fast screening can be performed with the use of only a few animals, before experiments with many animals are undertaken. Furthermore, we describe a refined hamster model, to which we added activity tracking as a valuable clinical parameter, and a histopathological scoring system for a standardized semi-quantitative evaluation of the lung pathology.
4. Materials and Methods
4.1. Animals
For both PCLS and in vivo inoculations, Syrian hamsters (Mesocricetus auratus; female), strain RjHan:AURA, were obtained from Janvier (Le Genest-Saint-Isle, France). They had an SPF (Specific Pathogen-Free) health status and were either 7 weeks (juvenile) or 8 to 12 months of age (adult) at arrival. To obtain PCLS samples from pigs, conventional pigs (Sus scrofa domesticus; male), TOPIGS Norsvin 70, were purchased from a high health status farm (Van Beek; Lelystad, The Netherlands). Upon arrival, the pigs were 4–5 months of age. To obtain PCLS samples from cats, domestic shorthair cats (Felis catus; male and female) were purchased from Marshall Bioresources (North Rose, NY, USA). Cats were raised as SPF cats, vaccinated against Rabies and Feline Rhinotracheitis-Calici-Panleukopenia, and were 4 months old upon arrival.
4.2. Virus and Cells
A SARS-CoV-2 isolate, referred to as SARS-CoV-2/human/NL/Lelystad/2020, was used in this study. The virus stock was prepared by two low-MOI passages (p1 and p2) in Vero E6 cells (ATCC® CRL-1586™; Manassas, VA, USA) after initial isolation from an oropharyngeal swab from a human subject. Cells were maintained on MEM (Gibco, RefNo 21090; Thermo Fischer Scientific; Waltham, MA, USA), supplemented with 5% FCS, 1% antibiotic/antimycotic (Gibco; Thermo Fischer Scientific; Waltham, MA, USA), 1% L-glutamine, 1% Minimal Essential Medium Non-Essential Amino Acids (MEM-NEAA) (all from Gibco; Thermo Fischer Scientific; Waltham, MA, USA). This medium is referred to as complete medium throughout the manuscript and was also used for cell culturing and virus propagation.
4.3. Next-Generation Sequencing (NGS)
Virus stock material from both passages (p1 and p2) was treated with benzonase (Sigma; Saint Louis, MO, USA) to remove host RNA/DNA (2 mL virus stock was incubated with 1 µL benzonase (250 U) and 2 µL 2 mM MgCl2 (Merck; Kenilworth, NJ, USA) for 4 h at 37 °C). RNA was isolated using the NucliSens easyMAG automated RNA isolation robot and NucliSENS easyMAG kit (Biomerieux; Marcy-l’Étoile, France), with 500 µL input material and 50 µL elution material. The obtained four elution fractions were pooled and concentrated using the Zymoclean RNA Clean&Concentrator kit (Zymo Research, RefNo R1013; Irvine, CA, USA). The cleaning/concentration was repeated twice to remove excessive content of small RNAs. The prepared RNA was subsequently analyzed with next-generation sequencing on an Illumina platform (MySeq System, Illumina; San Diego, CA, USA), using both an RNA-seq technique (RNA preparation with KAPA RNA HyperPrep Kit, Roche, RefNo 08098107702; Basel, Switzerland) and preparation of a cDNA library (KAPA HyperPlus Kit, Roche, RefNo 07962428001). Full sequences were obtained from both methods.
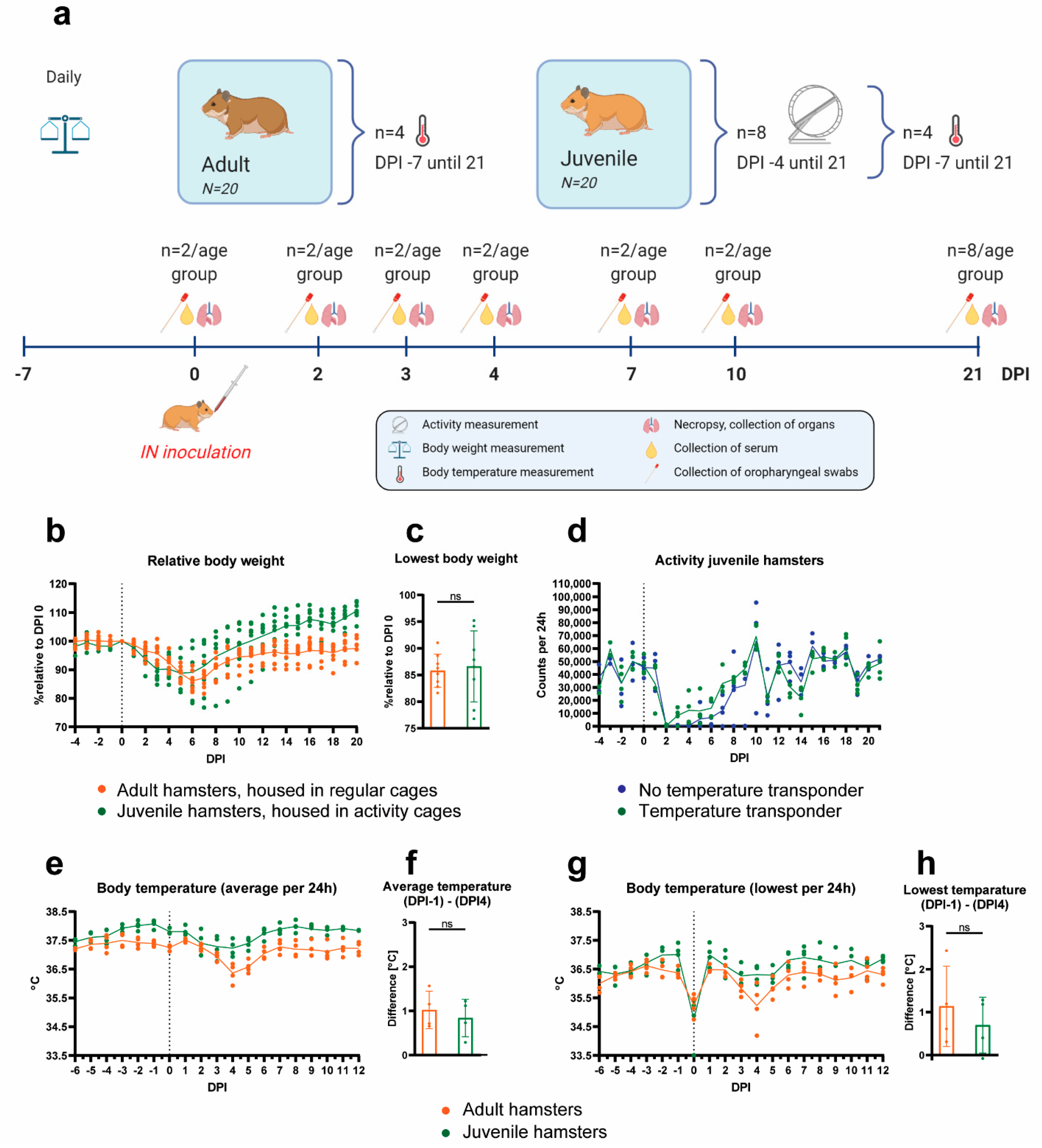
4.4. Hamster Experiment
Twenty juvenile (7 weeks old) and twenty adult (8–12 months old) Syrian hamsters were housed in individual cages and chipped subcutaneously upon arrival for identification. After an acclimatization period of 7 days, four juvenile and four adult hamsters were randomly selected and were implanted with a temperature transponder in the abdominal cavity (Anipill
®, Hérouville Saint-Clair, France). The temperature probes were set to measure the abdominal temperature every 15 min throughout the whole experiment. Seven days post-surgery, the four juvenile hamsters with the temperature transponders and another four juvenile hamsters without transponders were individually housed in cages with activity tracking wheels (Tecniplast, Buguggiate, Italy). The individual activity of the hamsters was measured by automatized counting of the wheel rotations when hamsters ran in the wheels, with four counts equaling one full rotation (perimeter approximately 97 cm). The counters were read out daily, approximately at the same time of the day (±2 h), and set back to zero. Activity was monitored from DPI −4 (days post-infection) until the end of the study (DPI 21). Body weights of the hamsters were measured daily from DPI −7 until the end of the study. Before inoculation, two juvenile and two adult hamsters were necropsied to collect uninfected tissues for comparison. All other hamsters (N = 36) were exposed to SARS-CoV-2 via intranasal inoculation with 100 µL of undiluted virus stock (dose of 10
4.5 TCID
50), which is the highest virus concentration that could be achieved with our virus stock. Virus was applied in a volume of 50 µL per nostril (100 µL in total) synchronous to the hamster’s breathing rhythm under anesthesia with 0.15 mg/kg medetomidine (Sedastart, ASTfarma; Oudewater, The Netherlands) and 100 mg/kg ketamine (Narketan, Vetoquinol; Breda, The Netherlands), which was antagonized by atipamezole (Sedastop, ASTfarma; Oudewater, The Netherlands). At 2, 3, 4, 7 and 10 days post-infection, two juvenile and two adult hamsters were euthanized to collect organs for pathohistological and virological analysis and blood for detection of antibodies. The remaining N = 16 hamsters (8 juvenile and 8 adult) were followed up to day 21, when they were euthanized and organs and blood was collected. Hamsters were monitored daily for general health and body weight loss. Details of the experimental scheme are shown in
Figure 3a. Humane endpoints were defined as follows: four or more infection-related clinical signs (such as ruffled fur, nasal or ocular discharge, coughing, sneezing, reduced activity while being handled, curled up position, neurological signs, impaired breathing (abdominal breathing or increased frequency) and 20% or more body weight loss compared to the day of infection); lethargy for more than 24 h; neurological seizures; severe respiratory distress.
4.5. Preparation of Precise-Cut Lung Slices (PCLS)
Donor animals for PCLS were euthanized by deep general anesthesia (hamsters and cats: medetomidine and ketamine, pigs: tiletamine, zolazepam, xylazine) followed by exsanguination. The lungs were carefully removed and either filled completely (hamster, cat) via the bronchus with 37 °C 1.5% agarose in RPMI 1640 (type VII-A low gelling temperature, Sigma) or a right cranial lung lobe (pig) was used. After agarose solidification on ice (10 min), the tissue was sliced into cubes of around 1 cm
3. The tissue cubes containing 1.5% agarose were subsequently embedded in 4% agarose by placing them in a 10 mL syringe with 3 mL of unsolidified 4% agarose (
Figure 1a). After filling of the syringe with additional agarose, solidification was continued for 10 min at 4 °C. The embedded lung tissue was subsequently cut into slices of 350 µm using a VT1000S vibratome (Leica, Amsterdam, Netherlands) set to a frequency of 80 Hz, speed of 2.5 mm/s, angle of 1.5 degrees and with a cycle speed of 60 slices/min. Each slice was placed in one well of a 24-well plate, pre-filled with 1 ml of RPMI 1640 medium (Gibco; Thermo Fischer Scientific; Waltham, MA, USA) supplemented with 5% FBS and 1% antibiotic/antimycotic solution (Gibco; Thermo Fischer Scientific; Waltham, MA, USA). After 2–4 h incubation at 37 °C and 5% CO
2, the slices were incubated with 100 µL SARS-COV-2 (5.5 log10 TCID
50/mL). Following overnight incubation, slices were washed three times with PBS (to remove extracellular virions), and at 24, 48 and 96 h post-infection, three slices per timepoint were placed in Trizol and frozen at ≤−70 °C until RNA isolation. In addition to the 350 µm PCLS, thicker slices (600 µm) were also incubated with SARS-CoV-2 (100 µL; 5.5 Log10 TCID
50/mL) for histology (hematoxylin and eosin stain) and IHC analysis following routine formalin fixation and paraffin embedding as described below.
4.6. Pathological and Histological Evaluation of Tissues
Upon necropsy, all major organs were examined macroscopically. Lungs were weighed and scored by a board-certified veterinary pathologist. A gross pathology score was assigned after evaluating the dorsal and ventral aspect of each lung (
Table 2). For histopathology, the respiratory tract (lower half of trachea, left lung lobe inflated with 10% neutral-buffered formalin, left nasal conchae), the gastro-intestinal tract, heart, spleen, liver, kidney and brain (left sagittal section of brainstem, cerebrum, cerebellum) were fixed in 10% neutral-buffered formalin (Klinipath BV; VWR; Radnor, PA, USA) for 1 week and embedded in paraffin. Sections of 3 μm were stained with hematoxylin and eosin (H&E) for histologic analysis. All organs were evaluated and lungs were semi-quantitatively scored for extent of lung pathology and severity of different histopathological parameters: alveoli (degeneration/necrosis of alveolar wall, type II pneumocyte proliferation, presence of inflammatory cells), inflammation of bronchi/bronchiole, inflammation of blood vessels and the presence of edema (
Table 2).
4.7. Immunohistochemistry (IHC) and Immunofluorescence (IF)
SARS-CoV-2 antigen was detected by immunohistochemistry in 10% formalin-fixed and paraffin-embedded tissue (left lung, left nasal conchae and trachea). Briefly, heat-induced epitope retrieval (HIER) method was used to prepare slides for IHC stain. After routinely dewaxing and inhibition of endogenous peroxidase (methanol/H
2O
2), the sections were heated for 15 min at 100 °C (Pascal, Dako, pressure cooker), in 10 mmol citrate buffer pH 6.0 (Dako S1699; Agilent; Santa Clara, CA, USA). Subsequently, the slides were blocked with 10% normal goat serum (Dako; Agilent; Santa Clara, CA, USA) and, after this, the primary polyclonal antibody rabbit anti-SARS-CoV NucleoProtein (Sino Biological, 40163-T62; Beijing, China) was added at a dilution of 1:2500 for 45 min at room temperature (RT). As secondary antibody, rabbit Envision peroxidase polymer (Dako, K4003; Agilent; Santa Clara, CA, USA) was used for 30 min at RT. The reaction was revealed with 3.3’-diaminobenzidine (DAB) (Dako K3468; Agilent; Santa Clara, CA, USA) as substrate. Slides were counterstained with hematoxylin. For immunofluorescence, sections were incubated with Alexa Fluor™ tyramide reagent (Invitrogen, Carlsbad, CA, USA) and mounted in antifading mounting medium containing DAPI (Vector laboratories, Peterborough, UK). Sections were photographed with an Olympus BX51 (fluorescence) microscope (Olympus corporation, Shinjuku, Tokyo, Japan) equipped with a high-resolution digital camera. Monochromatic digital photographs for immunofluorescence were false-colored using CellSense
® software (Olympus corporation, Shinjuku, Tokyo, Japan). Lungs were scored semi-quantitatively for antigen expression in the lungs, as shown in
Table 3.
4.8. Organ Suspensions and Oropharyngeal Swabs
The following organs were collected for evaluation of viral loads: respiratory tract (upper half of trachea, right lung lobes, right nasal conchae), gastro-intestinal tract (duodenum, colon spiral), brain (right part of the brainstem, cerebrum, cerebellum after a sagittal section). The organs were kept on melting ice during necropsy and subsequently stored at ≤−70 °C until further processing. To prepare organ homogenates, each organ was weighed individually and then added to 6 mL of MEM, supplemented with 1% antibiotic/antimycotic solution (Gibco; Thermo Fischer Scientific; Waltham, MA, USA). When suspensions of intestines or feces were prepared, 100 µg/mL gentamycin was added to the medium. The organs/feces were ground for 50 s at 6000 rpm using Ultra Turrax tubes and an Ultra Turrax Tube Drive (IKA; Staufen, Germany). Lungs were ground for a shorter period of time (30 s) because of their less compact consistency compared to other organs. All organ suspensions were cleared by centrifuging for 15 min at 3400× g at 4 °C. Subsequently, cleared organ homogenates were either directly suspended in Trizol-LS (Sigma; Saint Louis, MO, USA) at a ratio of 1:3 (one part suspension and 3 parts Trizol) and stored at ≤−15 °C until RNA isolation or directly frozen at ≤−70 °C for virus isolation.
Oropharyngeal swabs (MW100 DryswabTM Fine Tip, sterile, MWE, Essex, UK) were obtained from the hamsters and directly submerged in 2 mL cell culture medium. During necropsy, the tubes with swabs were kept on melting ice until further processing. In the lab, the tubes were vigorously vortexed for 30 s on a vortex (Labdancer, VWR) and then centrifuged for 5 min at 1500× g and 4 °C in a precooled centrifuge. Aliquots for RNA or virus isolation were prepared as described above for the organ suspensions.
4.9. RNA Extraction and PCR
4.9.1. RNA Extraction
From Trizol-LS-lysed samples, total RNA was extracted with the Direct-zol™ RNA MiniPrep kit (Zymo Research, RefNo R1013; Irvine, CA, USA) according to the manufacturer’s instructions, without DNase treatment. RNA was stored at ≤−70 °C until used for PCR.
4.9.2. PCR on E Gene RNA (Total E Gene PCR)
To detect the viral genomic RNA, we used the method described by Corman et al. [
36] with the E_Sarbeco primer/probe set. Of note, this PCR detects both genomic viral RNA and subgenomic RNA of the E gene, generated during virus replication. A one-step reverse-transcriptase quantitative PCR (RT-qPCR) protocol was used with the TaqMan
® Fast Virus 1-Step Master Mix (Applied Biosystems; Foster City, CA, USA), 500 mM of the forward and reverse primers, 250 nM probe and 5 µL RNA template. The RT-qPCR was performed on a LightCycler480 platform with the following cycling conditions: 55 °C 10 min, 95 °C for 2 min, 95 °C for 15 s and 58 °C for 30 s for 40 cycles. Primers were synthesized by Eurogentec (Luik, Belgium). All PCR results were expressed as RNA copy number per ng RNA (PCLS), per swab or per gram tissue (organs). Quantifications were performed based on a standard curve that was included in each PCR run. The standard curve was prepared by 10-fold serial dilutions of RNA isolated from a virus stock (SARS-CoV-2/human/NED/Lelystad/2020) and calibrated against an RNA standard obtained from the European Virus Archives (EVAg; Marseille, France) [
36].
4.9.3. PCR on Subgenomic E-Gene RNA (Subgenomic PCR)
For detection of subgenomic (sg) RNA produced by the virus during genome replication, the procedure described by Wölfel et al. [
18] was used, with the following modifications: the TaqMan
® Fast Virus 1-Step Master Mix (Applied Biosystems; Foster City, CA; USA) was utilized and the cycling conditions were the same as described above for the total viral RNA detection. Primers were synthesized by Eurogentec (Luik, Belgium). For the sgRNA PCR, a standard curve was prepared from serial 10-fold dilutions of synthetic RNA with a sequence identical to the PCR amplicon. The concentration of the synthetic RNA was determined photometrically. A standard curve was included in each PCR run.
4.10. Virus Isolation and Quantification
Clarified organ suspensions were serially diluted (1:10 initial and 4-fold dilutions) in complete cell culture medium. From each suspension, three independent dilutions were prepared and 50 µL of each dilution was added to wells of 96-well plates containing 15,000 Vero E6 cells per well in 100 µL. After 1.5 h incubation at 37 °C and 5% CO2, the supernatants from the first two serial dilutions (1:10 and 1:40) were replaced with fresh culture medium. The monolayers were subsequently incubated for 6 days and then fixed with 4% formaldehyde (VWR; Radnor, PA, USA) (15 min incubation), followed by fixation and permeabilization with ice-cold 100% methanol for 10 min and subsequent washing with PBS. For titer determination, the plates were stained with an immunoperoxidase monolayer assay (IPMA; see below) and scored for positive (stained) wells under the microscope. Each well where at least one focus of infection was found was scored as positive. Titers were calculated using the Spearman–Kärber algorithm and were expressed as TCID50/mL.
4.11. Immuno-Peroxidase Monolayer Assay (IPMA)
Fixed cell monolayers were treated with 1% Triton X-100 (MP Biomedicals LLC; Irvine, CA, USA) solution for 10 min (RT), washed with PBS (media preparation at Wageningen Bioveterinary Research (WBVR, Lelystad, The Netherlands)) containing 0.5% Tween-80 (Sigma; Saint Louis, MO, USA) and blocked for 30 min with PBS supplemented with 5% horse serum (Sigma; Saint Louis, MO, USA) at RT. Next, the blocking solution was discarded, and the monolayers were incubated with a rabbit antiserum for 1 h at 37 °C. This rabbit antiserum (rabbit-anti-SARS-CoV-2-S1-2ST (619F)) was raised against the S1A subdomain of the SARS-CoV-2 spike protein (residues 1-294), fused to a triple Strep-Tag and produced in HEK293T cells (Davids Biotechnologie GmbH). After washing the plates three times with PBS-Tween, the monolayers were incubated with goat-anti-rabbit-HRP (Dako; Agilent; Santa Clara, CA, USA) for 1 h at 37 °C. After another three washes with PBS-Tween solution, a freshly prepared AEC (3-Amino-9-ethylcarbazole) substrate solution (19 mL substrate buffer (0.05 M NaAc buffer, pH adjusted to 5.0 using 0.05 M HAc) + 1 mL 4 mg/mL AEC (Sigma; Saint Louis, MO, USA) stock solution in DMSO (MP Biomedicals; Santa Ana, CA, USA) + 50 µL 3% H2O2 (Merck; Kenilworth, NJ, USA)) was added and the monolayers were incubated until clear red-brown color developed (usually within 30–40 min). Plates were evaluated under a standard light microscope.
4.12. Detection of Neutralizing Antibodies
Upon necropsy of hamsters, blood was collected from the aorta in VACUETTE® CAT Serum Clot Activator tubes (Greiner Bio One; Krensmünster, Austria). Blood was allowed to clot for 1 h at RT and separated by subsequent centrifugation for 10 min at 1250× g at RT. Resultant serum aliquots were stored at ≤−15 °C before heat inactivation for 2 h at 56 °C before analysis.
Virus neutralization tests (VNT) were performed in 96-well plates by serially diluting serum samples in complete medium (initial 1:10 and then 3-fold serial dilutions; 50 µL per well). Each sample was diluted in two independent replicates. Subsequently, diluted sera were combined with 50 µL of SARS-CoV-2 in complete medium, at a dose of ~100 TCID50. After incubating for 1.5 h at RT, 15,000 Vero E6 cells/well in 50 µL complete medium were added to each well. Plates were incubated for 4 days at 37 °C and 5% CO2 before fixation with 4% formaldehyde (Boom, Meppel, the Netherlands) (15 min incubation), followed by fixation and permeabilization with ice-cold 100% methanol (10 min). After washing with PBS, the plates were stained with IPMA (see above). The titer of each duplicate was determined as the reciprocal value of the last dilution that showed ≥50% neutralization, as assessed visually under a standard light microscope. The titer of each sample was calculated as the average of the duplicate titers after log transformation and expressed as virus microneutralization titer 50 (MN50).
4.13. Statistical Analysis
Virus growth in PCLS over time (
Figure 1b,c) was assessed per species, by fitting a linear regression model, where the mean RNA copy numbers (total E-gene PCR or subgenomic PCR) at 48 and 96 h were compared to the mean copy number at 24 h.
Comparison between the greatest body weight loss, as well as the temperature drop at DPI 4 in adult and juvenile hamsters was performed with a t-test after checking the data’s normality using the D’Agostino–Pearson omnibus K2 test.
To compare differences in pathological scores or relative lung weights between adult and juvenile hamsters (
Figure 4c,d and
Figure 5j–l), linear or Poisson regression models were used. In these models, the response was the pathological score/relative lung weights and the explanatory variables were the age of the hamsters, time post-infection and their interaction. To account for non-linearity in time post-infection, basic spline terms were used. The model (linear or Poisson) that best fit the data (based on Akaike’s information criterion) was chosen for each particular dataset. The model variable significance was assessed using the ANOVA test.
To compare differences in viral load dynamics in respiratory organs and oropharyngeal swabs between adult and juvenile hamsters in time post-infection (
Figure 6a), a multivariate linear regression model was fit, where RNA copies or VI units were the response variable and days post-infection and age of the hamsters (juvenile/adult) were the explanatory variables. An identical approach was used for the neutralizing antibody titers (
Figure 6b), but for the analysis, the titer values were log-transformed by taking the natural log of each titer. To account for non-linearity in the relationship between time and virus copies/antibody titers, natural splines were used on the variable days post-infection. The model variable significance was assessed using the ANOVA test.
Statistical analysis was performed using the statistical software R version 4.0.2 [
37] to fit the linear and Poisson regression models. For the
t-tests and all figures, GraphPad Prism version 8.3.0 was used (GraphPad, San Diego, CA, USA).


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}