
Zoonotic Malaria: Non-Laverania Plasmodium Biology and Invasion Mechanisms


 and
and 
Abstract
:1. Introduction
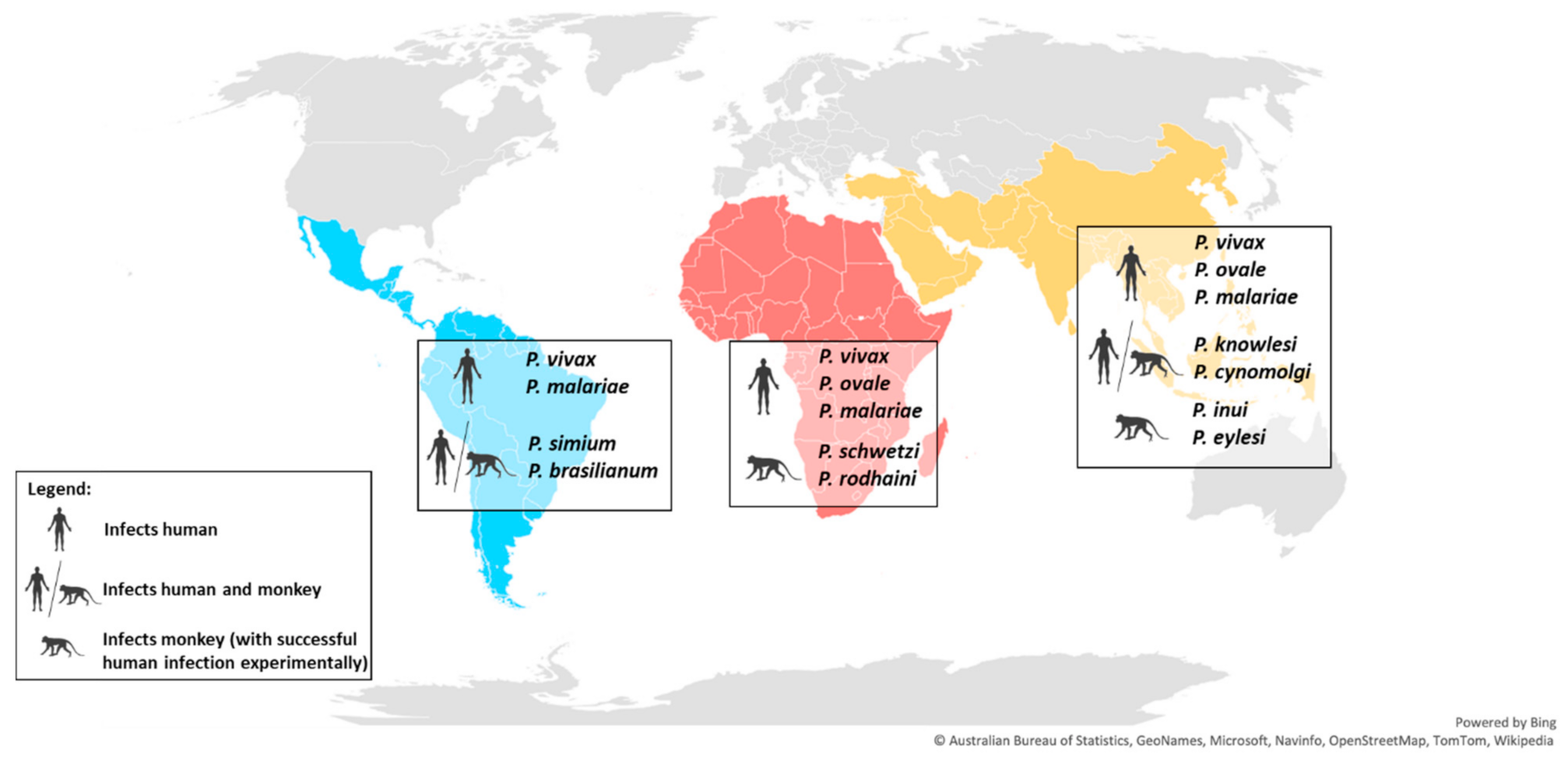
2. Geographical Distribution and Vertebrate Hosts of Zoonotic Plasmodium Species
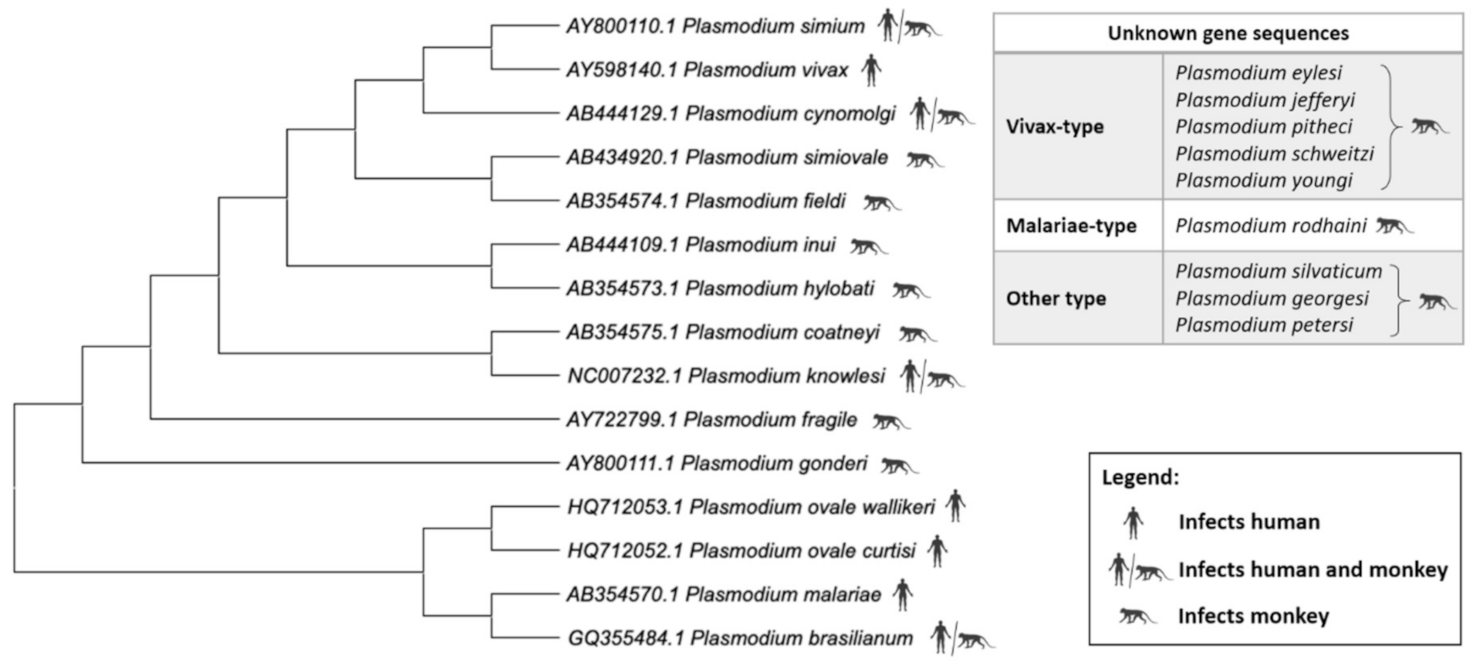
3. Genome Characteristics of Zoonotic Plasmodium Species
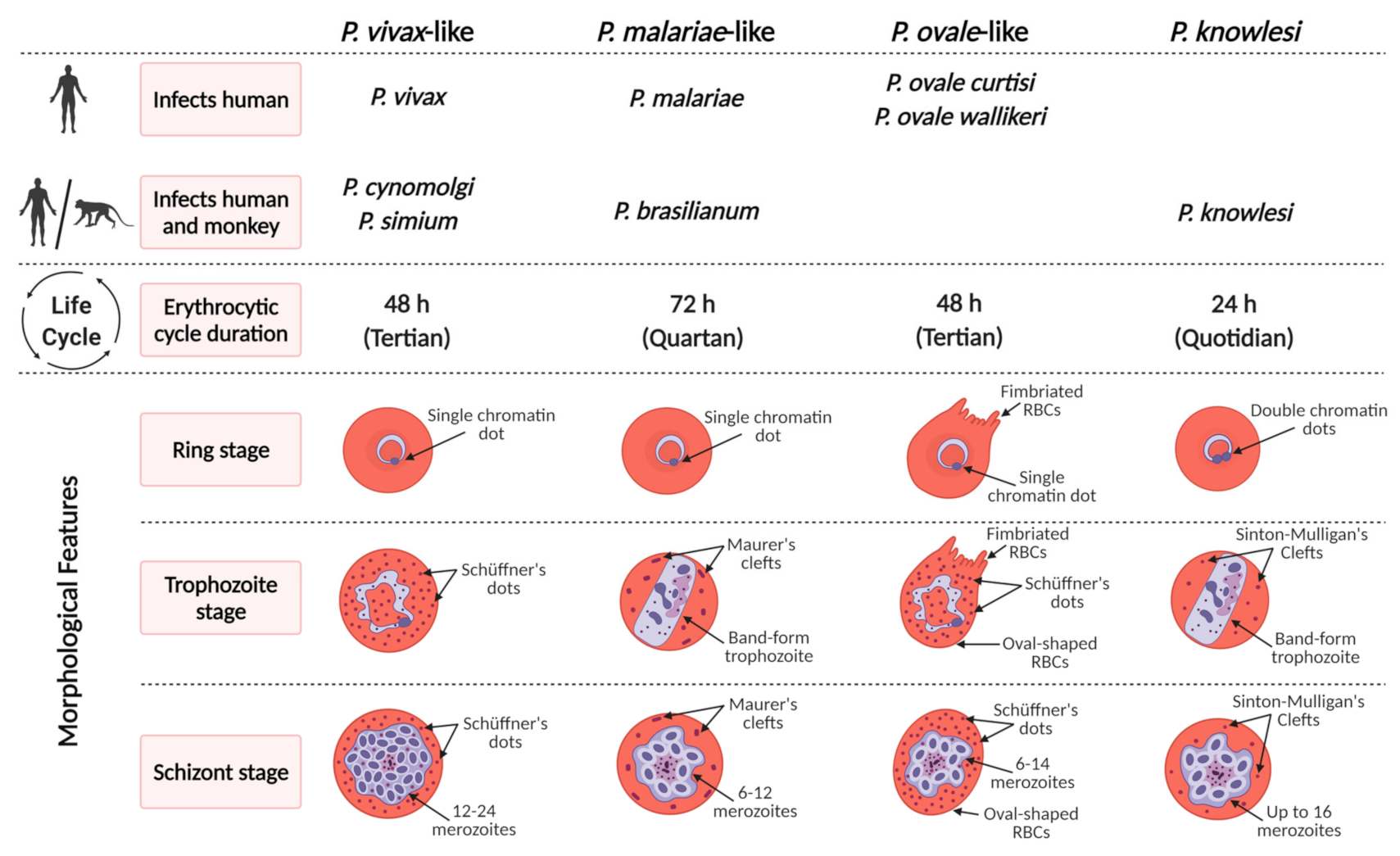
4. Characteristics and Pathology Associated with Individual Zoonotic Plasmodium Species
4.1. Plasmodium knowlesi
4.2. Plasmodium cynomolgi
4.3. Plasmodium simium
4.4. Plasmodium brasilianum
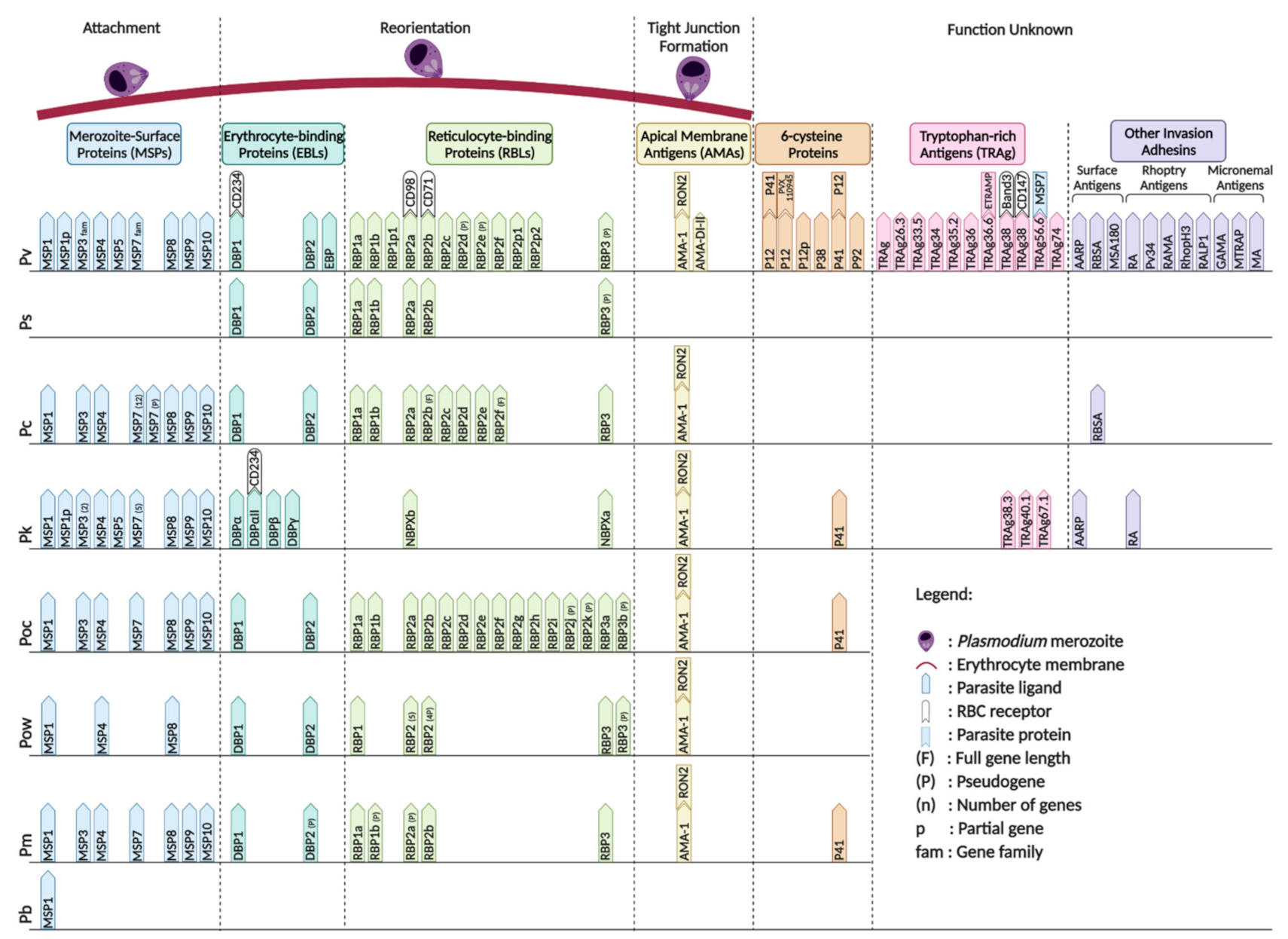
5. Role of Plasmodium Ligands in Erythrocytic Invasion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. World Malaria Report 2019; WHO: Geneva, Switzerland, 2019; p. 232. [Google Scholar]
- Koch, M.; Baum, J. The mechanics of malaria parasite invasion of the human erythrocyte-towards a reassessment of the host cell contribution. Cell Microbiol. 2016, 18, 319–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boddey, J.A.; Cowman, A.F. Plasmodium nesting: Remaking the erythrocyte from the inside out. Annu. Rev. Microbiol. 2013, 67, 243–269. [Google Scholar] [CrossRef]
- Ngwa, C.J.; Rosa, T.F.D.A.; Pradel, G. The Biology of Malaria Gametocytes; Rodriguez-Morales, A.J., Ed.; IntechOpen: Rijeka, Croatia, 2016. [Google Scholar]
- Smith, M.L.; Styczynski, M.P. Systems biology-based investigation of host-Plasmodium interactions. Trends Parasitol. 2018, 34, 617–632. [Google Scholar] [CrossRef]
- Brasil, P.; Zalis, M.G.; de Pina-Costa, A.; Siqueira, A.M.; Junior, C.B.; Silva, S.; Areas, A.L.L.; Pelajo-Machado, M.; de Alvarenga, D.A.M.; da Silva Santelli, A.C.F.; et al. Outbreak of human malaria caused by Plasmodium simium in the Atlantic Forest in Rio de Janeiro: A molecular epidemiological investigation. Lancet Glob. Health 2017, 5, e1038–e1046. [Google Scholar] [CrossRef] [Green Version]
- Lalremruata, A.; Magris, M.; Vivas-Martinez, S.; Koehler, M.; Esen, M.; Kempaiah, P.; Jeyaraj, S.; Perkins, D.J.; Mordmuller, B.; Metzger, W.G. Natural infection of Plasmodium brasilianum in humans: Man and monkey share quartan malaria parasites in the Venezuelan Amazon. EBioMedicine 2015, 2, 1186–1192. [Google Scholar] [CrossRef] [Green Version]
- Ta, T.H.; Hisam, S.; Lanza, M.; Jiram, A.I.; Ismail, N.; Rubio, J.M. First case of a naturally acquired human infection with Plasmodium cynomolgi. Malar. J. 2014, 13, 68. [Google Scholar] [CrossRef] [Green Version]
- Ramasamy, R. Zoonotic malaria-global overview and research and policy needs. Front. Public Health 2014, 2, 123. [Google Scholar] [CrossRef] [Green Version]
- Kawamoto, F.; Liu, Q.; Ferreira, M.U.; Tantular, I.S. How prevalent are Plasmodium ovale and P. malariae in East Asia? Parasitol. Today 1999, 15, 422–426. [Google Scholar] [CrossRef]
- Al-Maktari, M.T.; Bassiouny, H.K.; Al-Hamd, Z.S.; Assabri, A.M.; El-Massry, A.G.; Shatat, H.Z. Malaria status in Al-Hodeidah Governorate, Yemen: Malariometric parasitic survey & chloroquine resistance P. falciparum local strain. J. Egypt Soc. Parasitol. 2003, 33, 361–372. [Google Scholar]
- Autino, B.; Noris, A.; Russo, R.; Castelli, F. Epidemiology of malaria in endemic areas. Mediterr. J. Hematol. Infect. Dis. 2012, 4, e2012060. [Google Scholar] [CrossRef]
- Coatney, G.R.; Collins, W.E.; Warren, M.; Contacos, P.G. The Primate Malarias; CDC: Atlanta, GA, USA, 2003. Available online: https://stacks.cdc.gov/view/cdc/6538 (accessed on 1 July 2021).
- Eyles, D.E. The species of simian malaria: Taxonomy, morphology, life cycle, and geographical distribution of the monkey species. J. Parasitol. 1963, 49, 866–887. [Google Scholar] [CrossRef] [PubMed]
- Chin, W.; Moss, D.; Collins, W.E. The continuous cultivation of Plasmodium fragile by the method of Trager-Jensen. Am. J. Trop. Med. Hyg. 1979, 28, 591–592. [Google Scholar] [CrossRef]
- Handunnetti, S.M.; Mendis, K.N.; David, P.H. Antigenic variation of cloned Plasmodium fragile in its natural host Macaca sinica. Sequential appearance of successive variant antigenic types. J. Exp. Med. 1987, 165, 1269–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coatney, G.R.; Chin, W.; Contacos, P.G.; King, H.K. Plasmodium inui, a quartan-type malaria parasite of Old World monkeys transmissible to man. J. Parasitol. 1966, 52, 660–663. [Google Scholar] [CrossRef] [PubMed]
- Peters, W. Malaria of the orang-utan (Pongo pygmaeus) in Borneo. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1976, 275, 439–482. [Google Scholar] [CrossRef] [PubMed]
- Eyles, D.E.; Fong, Y.L.; Dunn, F.L.; Guinn, E.; Warren, M.; Sandosham, A.A. Plasmodium youngi N. sp., a malaria parasite of the Malayan Gibbon. Hylobates Lar Lar. Am. J. Trop. Med. Hyg. 1964, 13, 248–255. [Google Scholar] [CrossRef]
- Alvarenga, D.A.; Pina-Costa, A.; Bianco, C., Jr.; Moreira, S.B.; Brasil, P.; Pissinatti, A.; Daniel-Ribeiro, C.T.; Brito, C.F. New potential Plasmodium brasilianum hosts: Tamarin and marmoset monkeys (family Callitrichidae). Malar. J. 2017, 16, 71. [Google Scholar] [CrossRef] [Green Version]
- de Alvarenga, D.A.; de Pina-Costa, A.; de Sousa, T.N.; Pissinatti, A.; Zalis, M.G.; Suarez-Mutis, M.C.; Lourenco-de-Oliveira, R.; Brasil, P.; Daniel-Ribeiro, C.T.; de Brito, C.F. Simian malaria in the Brazilian Atlantic forest: First description of natural infection of capuchin monkeys (Cebinae subfamily) by Plasmodium simium. Malar. J. 2015, 14, 81. [Google Scholar] [CrossRef] [Green Version]
- Poirriez, J.; Dei-Cas, E.; Dujardin, L.; Landau, I. The blood-stages of Plasmodium georgesi, P. gonderi and P. petersi: Course of untreated infection in their natural hosts and additional morphological distinctive features. Parasitology 1995, 111, 547–554. [Google Scholar] [CrossRef]
- Poirriez, J.; Dei-Cas, E.; Landau, I. Further description of blood stages of Plasmodium petersi from Cercocebus albigena monkey. Folia Parasitol. 1994, 41, 168–172. [Google Scholar]
- Contacos, P.G.; Coatney, G.R.; Orihel, T.C.; Collins, W.E.; Chin, W.; Jeter, M.H. Transmission of Plasmodium schwetzi from the chimpanzee to man by mosquito bite. Am. J. Trop. Med. Hyg. 1970, 19, 190–195. [Google Scholar] [CrossRef]
- Li, M.I.; Mailepessov, D.; Vythilingam, I.; Lee, V.; Lam, P.; Ng, L.C.; Tan, C.H. Prevalence of simian malaria parasites in macaques of Singapore. PLoS Negl. Trop. Dis. 2021, 15, e0009110. [Google Scholar] [CrossRef]
- Iwagami, M.; Nakatsu, M.; Khattignavong, P.; Soundala, P.; Lorphachan, L.; Keomalaphet, S.; Xangsayalath, P.; Kawai, S.; Hongvanthong, B.; Brey, P.T.; et al. First case of human infection with Plasmodium knowlesi in Laos. PLoS Negl. Trop. Dis. 2018, 12, e0006244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jongwutiwes, S.; Putaporntip, C.; Iwasaki, T.; Sata, T.; Kanbara, H. Naturally acquired Plasmodium knowlesi malaria in human, Thailand. Emerg. Infect. Dis. 2004, 10, 2211–2213. [Google Scholar] [CrossRef] [PubMed]
- Luchavez, J.; Espino, F.; Curameng, P.; Espina, R.; Bell, D.; Chiodini, P.; Nolder, D.; Sutherland, C.; Lee, K.S.; Singh, B. Human Infections with Plasmodium knowlesi, the Philippines. Emerg. Infect. Dis. 2008, 14, 811–813. [Google Scholar] [CrossRef]
- Singh, B.; Kim Sung, L.; Matusop, A.; Radhakrishnan, A.; Shamsul, S.S.; Cox-Singh, J.; Thomas, A.; Conway, D.J. A large focus of naturally acquired Plasmodium knowlesi infections in human beings. Lancet 2004, 363, 1017–1024. [Google Scholar] [CrossRef]
- Tanizaki, R.; Ujiie, M.; Kato, Y.; Iwagami, M.; Hashimoto, A.; Kutsuna, S.; Takeshita, N.; Hayakawa, K.; Kanagawa, S.; Kano, S.; et al. First case of Plasmodium knowlesi infection in a Japanese traveller returning from Malaysia. Malar. J. 2013, 12, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Kadir, K.A.; Quintanilla-Zarinan, L.F.; Villano, J.; Houghton, P.; Du, H.; Singh, B.; Smith, D.G. Distribution and prevalence of malaria parasites among long-tailed macaques (Macaca fascicularis) in regional populations across Southeast Asia. Malar. J. 2016, 15, 450. [Google Scholar] [CrossRef] [Green Version]
- Imwong, M.; Madmanee, W.; Suwannasin, K.; Kunasol, C.; Peto, T.J.; Tripura, R.; von Seidlein, L.; Nguon, C.; Davoeung, C.; Day, N.P.J.; et al. Asymptomatic natural human infections with the simian malaria parasites Plasmodium cynomolgi and Plasmodium knowlesi. J. Infect. Dis. 2019, 219, 695–702. [Google Scholar] [CrossRef] [Green Version]
- McManus, K.F.; Taravella, A.M.; Henn, B.M.; Bustamante, C.D.; Sikora, M.; Cornejo, O.E. Population genetic analysis of the DARC locus (Duffy) reveals adaptation from standing variation associated with malaria resistance in humans. PLoS Genet. 2017, 13, e1006560. [Google Scholar] [CrossRef] [Green Version]
- Haynes, J.D.; Dalton, J.P.; Klotz, F.W.; McGinniss, M.H.; Hadley, T.J.; Hudson, D.E.; Miller, L.H. Receptor-like specificity of a Plasmodium knowlesi malarial protein that binds to Duffy antigen ligands on erythrocytes. J. Exp. Med. 1988, 167, 1873–1881. [Google Scholar] [CrossRef] [PubMed]
- Wertheimer, S.P.; Barnwell, J.W. Plasmodium vivax interaction with the human Duffy blood group glycoprotein: Identification of a parasite receptor-like protein. Exp. Parasitol. 1989, 69, 340–350. [Google Scholar] [CrossRef]
- Mendes, C.; Dias, F.; Figueiredo, J.; Mora, V.G.; Cano, J.; de Sousa, B.; do Rosario, V.E.; Benito, A.; Berzosa, P.; Arez, A.P. Duffy negative antigen is no longer a barrier to Plasmodium vivax—Molecular evidences from the African West Coast (Angola and Equatorial Guinea). PLoS Negl. Trop. Dis. 2011, 5, e1192. [Google Scholar] [CrossRef]
- Poirier, P.; Doderer-Lang, C.; Atchade, P.S.; Lemoine, J.P.; de l‘Isle, M.C.; Abou-Bacar, A.; Pfaff, A.W.; Brunet, J.; Arnoux, L.; Haar, E.; et al. The hide and seek of Plasmodium vivax in West Africa: Report from a large-scale study in Beninese asymptomatic subjects. Malar. J. 2016, 15, 570. [Google Scholar] [CrossRef] [Green Version]
- Motshoge, T.; Ababio, G.K.; Aleksenko, L.; Read, J.; Peloewetse, E.; Loeto, M.; Mosweunyane, T.; Moakofhi, K.; Ntebele, D.S.; Chihanga, S.; et al. Molecular evidence of high rates of asymptomatic P. vivax infection and very low P. falciparum malaria in Botswana. BMC Infect. Dis. 2016, 16, 520. [Google Scholar] [CrossRef] [Green Version]
- Fru-Cho, J.; Bumah, V.V.; Safeukui, I.; Nkuo-Akenji, T.; Titanji, V.P.; Haldar, K. Molecular typing reveals substantial Plasmodium vivax infection in asymptomatic adults in a rural area of Cameroon. Malar. J. 2014, 13, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, E.; Yewhalaw, D.; Zhong, D.; Zemene, E.; Degefa, T.; Tushune, K.; Ha, M.; Lee, M.C.; James, A.A.; Yan, G. Molecular epidemiology of Plasmodium vivax and Plasmodium falciparum malaria among Duffy-positive and Duffy-negative populations in Ethiopia. Malar. J. 2015, 14, 84. [Google Scholar] [CrossRef] [Green Version]
- Ryan, J.R.; Stoute, J.A.; Amon, J.; Dunton, R.F.; Mtalib, R.; Koros, J.; Owour, B.; Luckhart, S.; Wirtz, R.A.; Barnwell, J.W.; et al. Evidence for transmission of Plasmodium vivax among a duffy antigen negative population in Western Kenya. Am. J. Trop. Med. Hyg. 2006, 75, 575–581. [Google Scholar] [CrossRef] [Green Version]
- Menard, D.; Barnadas, C.; Bouchier, C.; Henry-Halldin, C.; Gray, L.R.; Ratsimbasoa, A.; Thonier, V.; Carod, J.F.; Domarle, O.; Colin, Y.; et al. Plasmodium vivax clinical malaria is commonly observed in Duffy-negative Malagasy people. Proc. Natl. Acad. Sci. USA 2010, 107, 5967–5971. [Google Scholar] [CrossRef] [Green Version]
- Niangaly, A.; Karthigayan, G.; Amed, O.; Coulibaly, D.; Sa, J.M.; Adams, M.; Travassos, M.A.; Ferrero, J.; Laurens, M.B.; Kone, A.K.; et al. Plasmodium vivax infections over 3 years in Duffy blood group negative Malians in Bandiagara, Mali. Am. J. Trop. Med. Hyg. 2017, 97, 744–752. [Google Scholar] [CrossRef] [Green Version]
- Wurtz, N.; Mint Lekweiry, K.; Bogreau, H.; Pradines, B.; Rogier, C.; Ould Mohamed Salem Boukhary, A.; Hafid, J.E.; Ould Ahmedou Salem, M.S.; Trape, J.F.; Basco, L.K.; et al. Vivax malaria in Mauritania includes infection of a Duffy-negative individual. Malar. J. 2011, 10, 336. [Google Scholar] [CrossRef] [Green Version]
- Niang, M.; Thiam, L.G.; Sow, A.; Loucoubar, C.; Bob, N.S.; Diop, F.; Diouf, B.; Niass, O.; Mansourou, A.; Varela, M.L.; et al. A molecular survey of acute febrile illnesses reveals Plasmodium vivax infections in Kedougou, southeastern Senegal. Malar. J. 2015, 14, 281. [Google Scholar] [CrossRef] [Green Version]
- Abdelraheem, M.H.; Albsheer, M.M.; Mohamed, H.S.; Amin, M.; Mahdi Abdel Hamid, M. Transmission of Plasmodium vivax in Duffy-negative individuals in central Sudan. Trans. R. Soc. Trop. Med. Hyg. 2016, 110, 258–260. [Google Scholar] [CrossRef]
- Asua, V.; Tukwasibwe, S.; Conrad, M.; Walakira, A.; Nankabirwa, J.I.; Mugenyi, L.; Kamya, M.R.; Nsobya, S.L.; Rosenthal, P.J. Plasmodium species infecting children presenting with malaria in Uganda. Am. J. Trop. Med. Hyg. 2017, 97, 753–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haiyambo, D.H.; Uusiku, P.; Mumbengegwi, D.; Pernica, J.M.; Bock, R.; Malleret, B.; Renia, L.; Greco, B.; Quaye, I.K. Molecular detection of P. vivax and P. ovale foci of infection in asymptomatic and symptomatic children in Northern Namibia. PLoS Negl. Trop. Dis. 2019, 13, e0007290. [Google Scholar] [CrossRef] [PubMed]
- Malleret, B.; Renia, L.; Russell, B. The unhealthy attraction of Plasmodium vivax to reticulocytes expressing transferrin receptor 1 (CD71). Int. J. Parasitol. 2017, 47, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Popovici, J.; Roesch, C.; Rougeron, V. The enigmatic mechanisms by which Plasmodium vivax infects Duffy-negative individuals. PLoS Pathog. 2020, 16, e1008258. [Google Scholar] [CrossRef] [Green Version]
- Bray, R.S. Studies on malaria in chimpanzees. V. The sporogonous cycle and mosquito transmission of Plasmodium vivax schwetzi. J. Parasitol. 1958, 44, 46–51. [Google Scholar] [CrossRef]
- Coatney, G.R. Simian malarias in man: Facts, implications, and predictions. Am. J. Trop. Med. Hyg. 1968, 17, 147–155. [Google Scholar] [CrossRef]
- Gilabert, A.; Otto, T.D.; Rutledge, G.G.; Franzon, B.; Ollomo, B.; Arnathau, C.; Durand, P.; Moukodoum, N.D.; Okouga, A.P.; Ngoubangoye, B.; et al. Plasmodium vivax-like genome sequences shed new insights into Plasmodium vivax biology and evolution. PLoS Biol. 2018, 16, e2006035. [Google Scholar] [CrossRef]
- Lourenco-de-Oliveira, R.; Deane, L.M. Simian malaria at two sites in the Brazilian Amazon. I--The infection rates of Plasmodium brasilianum in non-human primates. Mem. Inst. Oswaldo Cruz 1995, 90, 331–339. [Google Scholar] [CrossRef]
- Barnwell, J.W. Antigens of Plasmodium vivax blood stage parasites identified by monoclonal antibodies. Mem. Inst. Oswaldo Cruz 1986, 81, 59–61. [Google Scholar] [CrossRef]
- Cochrane, A.H.; Barnwell, J.W.; Collins, W.E.; Nussenzweig, R.S. Monoclonal antibodies produced against sporozoites of the human parasite Plasmodium malariae abolish infectivity of sporozoites of the simian parasite Plasmodium brasilianum. Infect. Immun. 1985, 50, 58–61. [Google Scholar] [CrossRef] [Green Version]
- Fandeur, T.; Volney, B.; Peneau, C.; de Thoisy, B. Monkeys of the rainforest in French Guiana are natural reservoirs for P. brasilianum/P. malariae malaria. Parasitology 2000, 120, 11–21. [Google Scholar] [CrossRef]
- Seed, T.M. Plasmodium simium:ultrastructure of erythrocytic phase. Exp. Parasitol. 1976, 39, 262–276. [Google Scholar] [CrossRef]
- Goldman, I.F.; Qari, S.H.; Millet, P.G.; Collins, W.E.; Lal, A.A. Circumsporozoite protein gene of Plasmodium simium, a Plasmodium vivax-like monkey malaria parasite. Mol. Biochem. Parasitol. 1993, 57, 177–180. [Google Scholar] [CrossRef]
- Early, A.M.; Daniels, R.F.; Farrell, T.M.; Grimsby, J.; Volkman, S.K.; Wirth, D.F.; MacInnis, B.L.; Neafsey, D.E. Detection of low-density Plasmodium falciparum infections using amplicon deep sequencing. Malar. J. 2019, 18, 219. [Google Scholar] [CrossRef] [Green Version]
- Carlton, J.; Silva, J.; Hall, N. The genome of model malaria parasites, and comparative genomics. Curr. Issues Mol. Biol. 2005, 7, 23–37. [Google Scholar]
- Kooij, T.W.; Carlton, J.M.; Bidwell, S.L.; Hall, N.; Ramesar, J.; Janse, C.J.; Waters, A.P. A Plasmodium whole-genome synteny map: Indels and synteny breakpoints as foci for species-specific genes. PLoS Pathog. 2005, 1, e44. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Sullivan, S.A.; Carlton, J.M. The biology of Plasmodium vivax explored through genomics. Ann. N. Y. Acad. Sci. 2015, 1342, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Carlton, J.M.; Adams, J.H.; Silva, J.C.; Bidwell, S.L.; Lorenzi, H.; Caler, E.; Crabtree, J.; Angiuoli, S.V.; Merino, E.F.; Amedeo, P.; et al. Comparative genomics of the neglected human malaria parasite Plasmodium vivax. Nature 2008, 455, 757–763. [Google Scholar] [CrossRef]
- Pain, A.; Bohme, U.; Berry, A.E.; Mungall, K.; Finn, R.D.; Jackson, A.P.; Mourier, T.; Mistry, J.; Pasini, E.M.; Aslett, M.A.; et al. The genome of the simian and human malaria parasite Plasmodium knowlesi. Nature 2008, 455, 799–803. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.L. Analysis of sequences from the extremely A + T-rich genome of Plasmodium falciparum. Gene 1987, 52, 103–109. [Google Scholar] [CrossRef]
- Aravind, L.; Iyer, L.M.; Wellems, T.E.; Miller, L.H. Plasmodium biology: Genomic gleanings. Cell 2003, 115, 771–785. [Google Scholar] [CrossRef] [Green Version]
- Mourier, T.; Alvarenga, D.A.M.d.; Kaushik, A.; Pina-Costa, A.d.; Guzmán-Vega, F.J.; Douvropoulou, O.; Guan, Q.; Forrester, S.; Abreu, F.V.S.d.; Júnior, C.B.; et al. The genome of the zoonotic malaria parasite Plasmodium simium reveals adaptions to host-switching. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Rutledge, G.G.; Bohme, U.; Sanders, M.; Reid, A.J.; Cotton, J.A.; Maiga-Ascofare, O.; Djimde, A.A.; Apinjoh, T.O.; Amenga-Etego, L.; Manske, M.; et al. Plasmodium malariae and P. ovale genomes provide insights into malaria parasite evolution. Nature 2017, 542, 101–104. [Google Scholar] [CrossRef] [Green Version]
- Pasini, E.M.; Bohme, U.; Rutledge, G.G.; Voorberg-Van der Wel, A.; Sanders, M.; Berriman, M.; Kocken, C.H.; Otto, T.D. An improved Plasmodium cynomolgi genome assembly reveals an unexpected methyltransferase gene expansion. Wellcome Open Res. 2017, 2, 42. [Google Scholar] [CrossRef] [Green Version]
- Ansari, H.R.; Templeton, T.J.; Subudhi, A.K.; Ramaprasad, A.; Tang, J.; Lu, F.; Naeem, R.; Hashish, Y.; Oguike, M.C.; Benavente, E.D.; et al. Genome-scale comparison of expanded gene families in Plasmodium ovale wallikeri and Plasmodium ovale curtisi with Plasmodium malariae and with other Plasmodium species. Int. J. Parasitol. 2016, 46, 685–696. [Google Scholar] [CrossRef] [Green Version]
- Daron, J.; Boissiere, A.; Boundenga, L.; Ngoubangoye, B.; Houze, S.; Arnathau, C.; Sidobre, C.; Trape, J.F.; Durand, P.; Renaud, F.; et al. Population genomic evidence of Plasmodium vivax Southeast Asian origin. Sci. Adv. 2021, 7. [Google Scholar] [CrossRef]
- Liu, W.; Li, Y.; Shaw, K.S.; Learn, G.H.; Plenderleith, L.J.; Malenke, J.A.; Sundararaman, S.A.; Ramirez, M.A.; Crystal, P.A.; Smith, A.G.; et al. African origin of the malaria parasite Plasmodium vivax. Nat. Commun. 2014, 5, 3346. [Google Scholar] [CrossRef] [Green Version]
- Loy, D.E.; Plenderleith, L.J.; Sundararaman, S.A.; Liu, W.; Gruszczyk, J.; Chen, Y.J.; Trimboli, S.; Learn, G.H.; MacLean, O.A.; Morgan, A.L.K.; et al. Evolutionary history of human Plasmodium vivax revealed by genome-wide analyses of related ape parasites. Proc. Natl. Acad. Sci. USA 2018, 115, E8450–E8459. [Google Scholar] [CrossRef] [Green Version]
- Mueller, R.L. Evolutionary rates, divergence dates, and the performance of mitochondrial genes in Bayesian phylogenetic analysis. Syst. Biol. 2006, 55, 289–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setiadi, W.; Sudoyo, H.; Trimarsanto, H.; Sihite, B.A.; Saragih, R.J.; Juliawaty, R.; Wangsamuda, S.; Asih, P.B.; Syafruddin, D. A zoonotic human infection with simian malaria, Plasmodium knowlesi, in Central Kalimantan, Indonesia. Malar. J. 2016, 15, 218. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, T.C.; Rodrigues, P.T.; Early, A.M.; Duarte, A.; Buery, J.C.; Bueno, M.G.; Catao-Dias, J.L.; Cerutti, C., Jr.; Rona, L.D.P.; Neafsey, D.E.; et al. Plasmodium simium: Population genomics reveals the origin of a reverse zoonosis. J. Infect. Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Winter, G.; Kawai, S.; Haeggstrom, M.; Kaneko, O.; von Euler, A.; Kawazu, S.; Palm, D.; Fernandez, V.; Wahlgren, M. SURFIN is a polymorphic antigen expressed on Plasmodium falciparum merozoites and infected erythrocytes. J. Exp. Med. 2005, 201, 1853–1863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frech, C.; Chen, N. Variant surface antigens of malaria parasites: Functional and evolutionary insights from comparative gene family classification and analysis. BMC Genom. 2013, 14, 427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talundzic, E.; Ravishankar, S.; Nayak, V.; Patel, D.S.; Olsen, C.; Sheth, M.; Batra, D.; Loparev, V.; Vannberg, F.O.; Udhayakumar, V.; et al. First full draft genome sequence of Plasmodium brasilianum. Genome Announc. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Hegner, R. Relative frequency of ring-stage plasmodia in reticulocytes and mature erythrocytes in man and monkey. Am. J. Epidemiol. 1938, 27, 690–718. [Google Scholar] [CrossRef]
- Kerlin, D.H.; Gatton, M.L. Preferential invasion by Plasmodium merozoites and the self-regulation of parasite burden. PLoS ONE 2013, 8, e57434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitchen, S.F. The infection of mature and immature erythrocytes by Plasmodium falciparum and Plasmodium malariae. Am. Soc. Trop. Med. Hyg. 1939, s1-19, 47–62. [Google Scholar] [CrossRef]
- Lingnau, A.; Doehring-Schwerdtfeger, E.; Maier, W.A. Evidence for 6-day cultivation of human Plasmodium malariae. Parasitol. Res. 1994, 80, 265–266. [Google Scholar] [CrossRef]
- Malleret, B.; Li, A.; Zhang, R.; Tan, K.S.; Suwanarusk, R.; Claser, C.; Cho, J.S.; Koh, E.G.; Chu, C.S.; Pukrittayakamee, S.; et al. Plasmodium vivax: Restricted tropism and rapid remodeling of CD71-positive reticulocytes. Blood 2015, 125, 1314–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aikawa, M. Morphological changes in erythrocytes induced by malarial parasites. Biol. Cell. 1988, 64, 173–181. [Google Scholar] [CrossRef]
- Russell, B.M.; Cooke, B.M. The rheopathobiology of Plasmodium vivax and other important primate malaria parasites. Trends Parasitol. 2017, 33, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Costa, F.T.; Lopes, S.C.; Ferrer, M.; Leite, J.A.; Martin-Jaular, L.; Bernabeu, M.; Nogueira, P.A.; Mourao, M.P.; Fernandez-Becerra, C.; Lacerda, M.V.; et al. On cytoadhesion of Plasmodium vivax: Raison d’etre? Mem. Inst. Oswaldo Cruz 2011, 106 (Suppl. S1), 79–84. [Google Scholar] [CrossRef] [Green Version]
- Suwanarusk, R.; Cooke, B.M.; Dondorp, A.M.; Silamut, K.; Sattabongkot, J.; White, N.J.; Udomsangpetch, R. The deformability of red blood cells parasitized by Plasmodium falciparum and P. vivax. J. Infect. Dis. 2004, 189, 190–194. [Google Scholar] [CrossRef] [Green Version]
- Kho, S.; Qotrunnada, L.; Leonardo, L.; Andries, B.; Wardani, P.A.I.; Fricot, A.; Henry, B.; Hardy, D.; Margyaningsih, N.I.; Apriyanti, D.; et al. Evaluation of splenic accumulation and colocalization of immature reticulocytes and Plasmodium vivax in asymptomatic malaria: A prospective human splenectomy study. PLoS Med. 2021, 18, e1003632. [Google Scholar] [CrossRef]
- Kho, S.; Qotrunnada, L.; Leonardo, L.; Andries, B.; Wardani, P.A.I.; Fricot, A.; Henry, B.; Hardy, D.; Margyaningsih, N.I.; Apriyanti, D.; et al. Hidden biomass of intact malaria parasites in the human spleen. N. Engl. J. Med. 2021, 384, 2067–2069. [Google Scholar] [CrossRef]
- Singh, B.; Daneshvar, C. Human infections and detection of Plasmodium knowlesi. Clin. Microbiol. Rev. 2013, 26, 165–184. [Google Scholar] [CrossRef] [Green Version]
- Grigg, M.J.; William, T.; Barber, B.E.; Rajahram, G.S.; Menon, J.; Schimann, E.; Piera, K.; Wilkes, C.S.; Patel, K.; Chandna, A.; et al. Age-related clinical spectrum of Plasmodium knowlesi malaria and predictors of severity. Clin. Infect. Dis. 2018, 67, 350–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Blanch, A.J.; Namvar, A.; Carmo, O.; Tiash, S.; Andrew, D.; Hanssen, E.; Rajagopal, V.; Dixon, M.W.A.; Tilley, L. Multimodal analysis of Plasmodium knowlesi-infected erythrocytes reveals large invaginations, swelling of the host cell, and rheological defects. Cell Microbiol. 2019, 21, e13005. [Google Scholar] [CrossRef] [Green Version]
- Asare, K.K.; Sakaguchi, M.; Lucky, A.B.; Asada, M.; Miyazaki, S.; Katakai, Y.; Kawai, S.; Song, C.; Murata, K.; Yahata, K.; et al. The Plasmodium knowlesi MAHRP2 ortholog localizes to structures connecting Sinton Mulligan‘s clefts in the infected erythrocyte. Parasitol. Int. 2018, 67, 481–492. [Google Scholar] [CrossRef]
- Anderios, F.; Noorrain, A.; Vythilingam, I. In vivo study of human Plasmodium knowlesi in Macaca fascicularis. Exp. Parasitol. 2010, 124, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Barber, B.E.; William, T.; Grigg, M.J.; Menon, J.; Auburn, S.; Marfurt, J.; Anstey, N.M.; Yeo, T.W. A prospective comparative study of knowlesi, falciparum, and vivax malaria in Sabah, Malaysia: High proportion with severe disease from Plasmodium knowlesi and Plasmodium vivax but no mortality with early referral and artesunate therapy. Clin. Infect. Dis. 2013, 56, 383–397. [Google Scholar] [CrossRef]
- Cox-Singh, J.; Davis, T.M.; Lee, K.S.; Shamsul, S.S.; Matusop, A.; Ratnam, S.; Rahman, H.A.; Conway, D.J.; Singh, B. Plasmodium knowlesi malaria in humans is widely distributed and potentially life threatening. Clin. Infect. Dis. 2008, 46, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Rajahram, G.S.; Barber, B.E.; William, T.; Menon, J.; Anstey, N.M.; Yeo, T.W. Deaths due to Plasmodium knowlesi malaria in Sabah, Malaysia: Association with reporting as Plasmodium malariae and delayed parenteral artesunate. Malar. J. 2012, 11, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figtree, M.; Lee, R.; Bain, L.; Kennedy, T.; Mackertich, S.; Urban, M.; Cheng, Q.; Hudson, B.J. Plasmodium knowlesi in human, Indonesian Borneo. Emerg. Infect. Dis. 2010, 16, 672–674. [Google Scholar] [CrossRef] [PubMed]
- Willmann, M.; Ahmed, A.; Siner, A.; Wong, I.T.; Woon, L.C.; Singh, B.; Krishna, S.; Cox-Singh, J. Laboratory markers of disease severity in Plasmodium knowlesi infection: A case control study. Malar. J. 2012, 11, 363. [Google Scholar] [CrossRef] [Green Version]
- Daneshvar, C.; Davis, T.M.; Cox-Singh, J.; Rafa’ee, M.Z.; Zakaria, S.K.; Divis, P.C.; Singh, B. Clinical and laboratory features of human Plasmodium knowlesi infection. Clin. Infect. Dis. 2009, 49, 852–860. [Google Scholar] [CrossRef] [Green Version]
- Anstey, N.M.; Handojo, T.; Pain, M.C.; Kenangalem, E.; Tjitra, E.; Price, R.N.; Maguire, G.P. Lung injury in vivax malaria: Pathophysiological evidence for pulmonary vascular sequestration and posttreatment alveolar-capillary inflammation. J. Infect. Dis. 2007, 195, 589–596. [Google Scholar] [CrossRef] [Green Version]
- Cox-Singh, J.; Singh, B.; Daneshvar, C.; Planche, T.; Parker-Williams, J.; Krishna, S. Anti-inflammatory cytokines predominate in acute human Plasmodium knowlesi infections. PLoS ONE 2011, 6, e20541. [Google Scholar] [CrossRef] [Green Version]
- Andrade, B.B.; Reis-Filho, A.; Souza-Neto, S.M.; Clarencio, J.; Camargo, L.M.; Barral, A.; Barral-Netto, M. Severe Plasmodium vivax malaria exhibits marked inflammatory imbalance. Malar. J. 2010, 9, 13. [Google Scholar] [CrossRef] [Green Version]
- Chua, A.C.Y.; Ong, J.J.Y.; Malleret, B.; Suwanarusk, R.; Kosaisavee, V.; Zeeman, A.M.; Cooper, C.A.; Tan, K.S.W.; Zhang, R.; Tan, B.H.; et al. Robust continuous in vitro culture of the Plasmodium cynomolgi erythrocytic stages. Nat. Commun. 2019, 10, 3635. [Google Scholar] [CrossRef] [Green Version]
- Baird, J.K. Evidence and implications of mortality associated with acute Plasmodium vivax malaria. Clin. Microbiol. Rev. 2013, 26, 36–57. [Google Scholar] [CrossRef] [Green Version]
- Hemmer, C.J.; Holst, F.G.; Kern, P.; Chiwakata, C.B.; Dietrich, M.; Reisinger, E.C. Stronger host response per parasitized erythrocyte in Plasmodium vivax or ovale than in Plasmodium falciparum malaria. Trop. Med. Int. Health 2006, 11, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Joyner, C.; Moreno, A.; Meyer, E.V.; Cabrera-Mora, M.; Ma, H.C.; Kissinger, J.C.; Barnwell, J.W.; Galinski, M.R. Plasmodium cynomolgi infections in rhesus macaques display clinical and parasitological features pertinent to modelling vivax malaria pathology and relapse infections. Malar. J. 2016, 15, 451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuvin, S.F.; Beye, H.K.; Stohlman, F., Jr.; Contacos, P.G.; Coatney, G.R. Clinical and physiological responses in sporozoite-induced B strain Plasmodium cynomolgi and Plasmodium vivax infections in normal volunteers. Trans. R. Soc. Trop. Med. Hyg. 1962, 56, 371–378. [Google Scholar] [CrossRef]
- Hartmeyer, G.N.; Stensvold, C.R.; Fabricius, T.; Marmolin, E.S.; Hoegh, S.V.; Nielsen, H.V.; Kemp, M.; Vestergaard, L.S. Plasmodium cynomolgi as cause of malaria in tourist to Southeast Asia, 2018. Emerg. Infect. Dis. 2019, 25, 1936–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, L.H. Compatibility of relapse patterns of Plasmodium cynomolgi infections in rhesus monkeys with continuous cyclical development and hypnozoite concepts of relapse. Am. J. Trop. Med. Hyg. 1986, 35, 1077–1099. [Google Scholar] [CrossRef]
- Deane, L.M. Simian malaria in Brazil. Mem. Inst. Oswaldo Cruz 1992, 87 (Suppl. S3), 1–20. [Google Scholar] [CrossRef]
- Lal, A.A.; de la Cruz, V.F.; Campbell, G.H.; Procell, P.M.; Collins, W.E.; McCutchan, T.F. Structure of the circumsporozoite gene of Plasmodium malariae. Mol. Biochem. Parasitol. 1988, 30, 291–294. [Google Scholar] [CrossRef]
- Collins, W.E.; Jeffery, G.M. Plasmodium malariae: Parasite and disease. Clin. Microbiol. Rev. 2007, 20, 579–592. [Google Scholar] [CrossRef] [Green Version]
- Li, A.; Russell, B.; Renia, L.; Lek-Uthai, U.; Nosten, F.; Lim, C.T. High density of ‘spiky‘ excrescences covering the surface of an erythrocyte infected with Plasmodium malariae. Br. J. Haematol. 2010, 151, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erkenswick, G.A.; Watsa, M.; Pacheco, M.A.; Escalante, A.A.; Parker, P.G. Chronic Plasmodium brasilianum infections in wild Peruvian tamarins. PLoS ONE 2017, 12, e0184504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitchen, S.F. The infection of reticulocytes by Plasmodium vivax. Am. Soc. Trop. Med. Hyg. 1938, s1-18, 347–359. [Google Scholar] [CrossRef]
- Kosaisavee, V.; Suwanarusk, R.; Chua, A.C.Y.; Kyle, D.E.; Malleret, B.; Zhang, R.; Imwong, M.; Imerbsin, R.; Ubalee, R.; Samano-Sanchez, H.; et al. Strict tropism for CD71(+)/CD234(+) human reticulocytes limits the zoonotic potential of Plasmodium cynomolgi. Blood 2017, 130, 1357–1363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tazi, L.; Ayala, F.J. Unresolved direction of host transfer of Plasmodium vivax v. P. simium and P. malariae v. P. brasilianum. Infect. Genet. Evol. 2011, 11, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Lim, C.; Hansen, E.; DeSimone, T.M.; Moreno, Y.; Junker, K.; Bei, A.; Brugnara, C.; Buckee, C.O.; Duraisingh, M.T. Expansion of host cellular niche can drive adaptation of a zoonotic malaria parasite to humans. Nat. Commun. 2013, 4, 1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, R.W.; Hall, J.; Rangkuti, F.; Ho, Y.S.; Almond, N.; Mitchell, G.H.; Pain, A.; Holder, A.A.; Blackman, M.J. Adaptation of the genetically tractable malaria pathogen Plasmodium knowlesi to continuous culture in human erythrocytes. Proc. Natl. Acad. Sci. USA 2013, 110, 531–536. [Google Scholar] [CrossRef] [Green Version]
- Galinski, M.R.; Barnwell, J.W. Plasmodium vivax: Merozoites, invasion of reticulocytes and considerations for malaria vaccine development. Parasitol. Today 1996, 12, 20–29. [Google Scholar] [CrossRef]
- Drew, D.R.; Sanders, P.R.; Weiss, G.; Gilson, P.R.; Crabb, B.S.; Beeson, J.G. Functional conservation of the AMA1 host-cell invasion ligand between P. falciparum and P. vivax: A novel platform to accelerate vaccine and drug development. J. Infect. Dis. 2018, 217, 498–507. [Google Scholar] [CrossRef]
- Rodríguez, L.E.; Urquiza, M.; Ocampo, M.; Curtidor, H.; Suárez, J.; García, J.; Vera, R.; Puentes, Á.; López, R.; Pinto, M.; et al. Plasmodium vivax MSP-1 peptides have high specific binding activity to human reticulocytes. Vaccine 2002, 20, 1331–1339. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, Y.; Ito, D.; Kong, D.H.; Ha, K.S.; Chen, J.H.; Lu, F.; Li, J.; Wang, B.; Takashima, E.; et al. The Plasmodium vivax merozoite surface protein 1 paralog is a novel erythrocyte-binding ligand of P. vivax. Infect. Immun. 2013, 81, 1585–1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beeson, J.G.; Drew, D.R.; Boyle, M.J.; Feng, G.; Fowkes, F.J.; Richards, J.S. Merozoite surface proteins in red blood cell invasion, immunity and vaccines against malaria. FEMS Microbiol. Rev. 2016, 40, 343–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, L.J.; Dietrich, M.H.; Nguitragool, W.; Tham, W.H. Plasmodium vivax reticulocyte binding proteins for invasion into reticulocytes. Cell Microbiol. 2020, 22, e13110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malleret, B.; Sahili, A.E.; Tay, M.Z.; Carissimo, G.; Ong, A.S.M.; Novera, W.; Lin, J.; Suwanarusk, R.; Kosaisavee, V.; Chu, T.T.T.; et al. Plasmodium vivax binds host CD98hc (SLC3A2) to enter immature red blood cells. Nat. Microbiol. 2021. [Google Scholar] [CrossRef]
- Gruszczyk, J.; Huang, R.K.; Chan, L.J.; Menant, S.; Hong, C.; Murphy, J.M.; Mok, Y.F.; Griffin, M.D.W.; Pearson, R.D.; Wong, W.; et al. Cryo-EM structure of an essential Plasmodium vivax invasion complex. Nature 2018, 559, 135–139. [Google Scholar] [CrossRef]
- Gruszczyk, J.; Kanjee, U.; Chan, L.J.; Menant, S.; Malleret, B.; Lim, N.T.Y.; Schmidt, C.Q.; Mok, Y.F.; Lin, K.M.; Pearson, R.D.; et al. Transferrin receptor 1 is a reticulocyte-specific receptor for Plasmodium vivax. Science 2018, 359, 48–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scully, E.J.; Kanjee, U.; Duraisingh, M.T. Molecular interactions governing host-specificity of blood stage malaria parasites. Curr. Opin. Microbiol. 2017, 40, 21–31. [Google Scholar] [CrossRef]
- Lim, K.L.; Amir, A.; Lau, Y.L.; Fong, M.Y. The Duffy binding protein (PkDBPalphaII) of Plasmodium knowlesi from Peninsular Malaysia and Malaysian Borneo show different binding activity level to human erythrocytes. Malar. J. 2017, 16, 331. [Google Scholar] [CrossRef] [Green Version]
- Gunalan, K.; Lo, E.; Hostetler, J.B.; Yewhalaw, D.; Mu, J.; Neafsey, D.E.; Yan, G.; Miller, L.H. Role of Plasmodium vivax Duffy-binding protein 1 in invasion of Duffy-null Africans. Proc. Natl. Acad. Sci. USA 2016, 113, 6271–6276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hester, J.; Chan, E.R.; Menard, D.; Mercereau-Puijalon, O.; Barnwell, J.; Zimmerman, P.A.; Serre, D. De novo assembly of a field isolate genome reveals novel Plasmodium vivax erythrocyte invasion genes. PLoS Negl. Trop. Dis. 2013, 7, e2569. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.H.; Mason, S.J.; Dvorak, J.A.; McGinniss, M.H.; Rothman, I.K. Erythrocyte receptors for (Plasmodium knowlesi) malaria: Duffy blood group determinants. Science 1975, 189, 561–563. [Google Scholar] [CrossRef]
- Golassa, L.; Amenga-Etego, L.; Lo, E.; Amambua-Ngwa, A. The biology of unconventional invasion of Duffy-negative reticulocytes by Plasmodium vivax and its implication in malaria epidemiology and public health. Malar. J. 2020, 19, 299. [Google Scholar] [CrossRef]
- Arévalo-Pinzón, G.; Bermúdez, M.; Hernández, D.; Curtidor, H.; Patarroyocorresponding, M.A. Plasmodium vivax ligand-receptor interaction: PvAMA-1 domain I contains the minimal regions for specific interaction with CD71+ reticulocytes. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Ito, D.; Chen, J.H.; Lu, F.; Cheng, Y.; Wang, B.; Ha, K.S.; Cao, J.; Torii, M.; Sattabongkot, J.; et al. Pv12, a 6-Cys antigen of Plasmodium vivax, is localized to the merozoite rhoptry. Parasitol. Int. 2012, 61, 443–449. [Google Scholar] [CrossRef]
- Hostetler, J.B.; Sharma, S.; Bartholdson, S.J.; Wright, G.J.; Fairhurst, R.M.; Rayner, J.C. A library of Plasmodium vivax recombinant merozoite proteins reveals new vaccine candidates and protein-protein interactions. PLoS Negl. Trop. Dis. 2015, 9, e0004264. [Google Scholar] [CrossRef]
- Mongui, A.; Angel, D.I.; Guzman, C.; Vanegas, M.; Patarroyo, M.A. Characterisation of the Plasmodium vivax Pv38 antigen. Biochem. Biophys. Res. Commun. 2008, 376, 326–330. [Google Scholar] [CrossRef]
- Angel, D.I.; Mongui, A.; Ardila, J.; Vanegas, M.; Patarroyo, M.A. The Plasmodium vivax Pv41 surface protein: Identification and characterization. Biochem. Biophys. Res. Commun. 2008, 377, 1113–1117. [Google Scholar] [CrossRef]
- Arredondo, S.A.; Kappe, S.H.I. The s48/45 six-cysteine proteins: Mediators of interaction throughout the Plasmodium life cycle. Int. J. Parasitol. 2017, 47, 409–423. [Google Scholar] [CrossRef]
- Ahmed, M.A.; Chu, K.B.; Quan, F.S. The Plasmodium knowlesi Pk41 surface protein diversity, natural selection, sub population and geographical clustering: A 6-cysteine protein family member. PeerJ 2018, 6, e6141. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Lu, F.; Cheng, Y.; Chen, J.H.; Jeon, H.Y.; Ha, K.S.; Cao, J.; Nyunt, M.H.; Han, J.H.; Lee, S.K.; et al. Immunoprofiling of the tryptophan-rich antigen family in Plasmodium vivax. Infect. Immun. 2015, 83, 3083–3095. [Google Scholar] [CrossRef] [Green Version]
- Zeeshan, M.; Bora, H.; Sharma, Y.D. Presence of memory T cells and naturally acquired antibodies in Plasmodium vivax malaria-exposed individuals against a group of tryptophan-rich antigens with conserved sequences. J. Infect. Dis. 2013, 207, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Zeeshan, M.; Tyagi, K.; Sharma, Y.D. CD4+ T cell response correlates with naturally acquired antibodies against Plasmodium vivax tryptophan-rich antigens. Infect. Immun. 2015, 83, 2018–2029. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, K.; Hossain, M.E.; Thakur, V.; Aggarwal, P.; Malhotra, P.; Mohmmed, A.; Sharma, Y.D. Plasmodium vivax tryptophan rich antigen PvTRAg36.6 interacts with PvETRAMP and PvTRAg56.6 interacts with PvMSP7 during erythrocytic stages of the parasite. PLoS ONE 2016, 11, e0151065. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.S.; Choudhary, V.; Zeeshan, M.; Tyagi, R.K.; Rathore, S.; Sharma, Y.D. Interaction of Plasmodium vivax tryptophan-rich antigen PvTRAg38 with Band 3 on human erythrocyte surface facilitates parasite growth. J. Biol. Chem. 2015, 290, 20257–20272. [Google Scholar] [CrossRef] [Green Version]
- Rathore, S.; Dass, S.; Kandari, D.; Kaur, I.; Gupta, M.; Sharma, Y.D. Basigin interacts with Plasmodium vivax tryptophan-rich antigen PvTRAg38 as a second erythrocyte receptor to promote parasite growth. J. Biol. Chem. 2017, 292, 462–476. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, K.; Gupta, D.; Saini, E.; Choudhary, S.; Jamwal, A.; Alam, M.S.; Zeeshan, M.; Tyagi, R.K.; Sharma, Y.D. Recognition of human erythrocyte receptors by the tryptophan-rich antigens of monkey malaria parasite Plasmodium knowlesi. PLoS ONE 2015, 10, e0138691. [Google Scholar] [CrossRef]
- Muh, F.; Ahmed, M.A.; Han, J.H.; Nyunt, M.H.; Lee, S.K.; Lau, Y.L.; Kaneko, O.; Han, E.T. Cross-species analysis of apical asparagine-rich protein of Plasmodium vivax and Plasmodium knowlesi. Sci. Rep. 2018, 8, 5781. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Perez, D.A.; Baquero, L.A.; Chitiva-Ardila, D.M.; Patarroyo, M.A. Characterising PvRBSA: An exclusive protein from Plasmodium species infecting reticulocytes. Parasit. Vectors 2017, 10, 243. [Google Scholar] [CrossRef] [Green Version]
- Muh, F.; Han, J.H.; Nyunt, M.H.; Lee, S.K.; Jeon, H.Y.; Ha, K.S.; Park, W.S.; Hong, S.H.; Ahmed, M.A.; Na, S.; et al. Identification of a novel merozoite surface antigen of Plasmodium vivax, PvMSA180. Malar. J. 2017, 16, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Changrob, S.; Wang, B.; Han, J.H.; Lee, S.K.; Nyunt, M.H.; Lim, C.S.; Tsuboi, T.; Chootong, P.; Han, E.T. Correction: Naturally-acquired immune response against Plasmodium vivax rhoptry-associated membrane antigen. PLoS ONE 2016, 11, e0153110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mongui, A.; Perez-Leal, O.; Rojas-Caraballo, J.; Angel, D.I.; Cortes, J.; Patarroyo, M.A. Identifying and characterising the Plasmodium falciparum RhopH3 Plasmodium vivax homologue. Biochem. Biophys. Res. Commun. 2007, 358, 861–866. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Li, J.; Ito, D.; Kong, D.H.; Ha, K.S.; Lu, F.; Wang, B.; Sattabongkot, J.; Lim, C.S.; Tsuboi, T.; et al. Antigenicity and immunogenicity of PvRALP1, a novel Plasmodium vivax rhoptry neck protein. Malar. J. 2015, 14, 186. [Google Scholar] [CrossRef] [Green Version]
- Anand, G.; Reddy, K.S.; Pandey, A.K.; Mian, S.Y.; Singh, H.; Mittal, S.A.; Amlabu, E.; Bassat, Q.; Mayor, A.; Chauhan, V.S.; et al. A novel Plasmodium falciparum rhoptry associated adhesin mediates erythrocyte invasion through the sialic-acid dependent pathway. Sci. Rep. 2016, 6, 29185. [Google Scholar] [CrossRef] [Green Version]
- Baquero, L.A.; Moreno-Perez, D.A.; Garzon-Ospina, D.; Forero-Rodriguez, J.; Ortiz-Suarez, H.D.; Patarroyo, M.A. PvGAMA reticulocyte binding activity: Predicting conserved functional regions by natural selection analysis. Parasit. Vectors 2017, 10, 251. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Lu, F.; Wang, B.; Li, J.; Han, J.H.; Ito, D.; Kong, D.H.; Jiang, L.; Wu, J.; Ha, K.S.; et al. Plasmodium vivax GPI-anchored micronemal antigen (PvGAMA) binds human erythrocytes independent of Duffy antigen status. Sci. Rep. 2016, 6, 35581. [Google Scholar] [CrossRef]
- Malleret, B. Plasmodium kinase disruption: New hopes for anti-malarial drug discovery. Br. J. Haematol. 2020, 188, 603–604. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parasite Distribution | Plasmodium Species | Human Infection | Natural Host(s) | References |
|---|---|---|---|---|
| Asia Africa America | P. vivax | ● | Human (Homo sapiens) | [1] |
| P. malariae | ● | Human (Homo sapiens) | [6,10] | |
| Asia Africa | P. ovale | ● | Human (Homo sapiens) | [10,11,12] |
| Asia | P. coatneyi | ● | Long-tailed macaque (Macaca fascicularis) | [13,14] |
| P. cynomolgi | ●● | Long-tailed macaque (Macaca fascicularis), Pig-tailed macaque (Macaca nemestrina), Bonnet macaque (Macaca radiata), Formosanrock macaque (Macaca cyclopis), Rhesus macaque (Macaca mulatta), Silvered leaf monkey (Presbytis cristatus), Hanuman langur (Presbytis entellus) | [13,14] | |
| P. eylesi | ● | Lar gibbon (Hylobates lar) | [13] | |
| P. fieldi | ● | Long-tailed macaque (Macaca fascicularis), Pig-tailed macaque (Macaca nemestrina), Bonnet macaque (Macaca radiata), Rhesus macaque (Macaca mulatta), Baboon (Papio doguera) | [13,14] | |
| P. fragile | ● | Bonnet macaque (Macaca radiata), Toque macaque (Macaca sinica) | [13,15,16] | |
| P. hylobati | ● | Silvery gibbon (Hylobates moloch) | [13] | |
| P. inui | ● | Cynopithecus niger (Macaca nigra), Formosanrock macaque (Macaca cyclopis), Long-tailed macaque (Macaca fascicularis), Rhesus macaque (Macaca mulatta), Pig-tailed macaque (Macaca nemestrina), Bonnet macaque (Macaca radiata), Silvered leaf monkey (Presbytis cristatus), Dusky leaf monkey (Presbytis obscurus) | [13,14,17] | |
| P. jefferyi | ● | Lar gibbon (Hylobates lar) | [13] | |
| P. knowlesi | ● | Long-tailed macaque (Macaca fascicularis), Pig-tailed macaque (Macaca nemestrina), Black-crested Sumatran langur (Presbytis melalophos) | [13,14] | |
| P. pitheci | ● | Orangutans (Pongo pygmaeus) | [13,18] | |
| P. simiovale | ● | Toque macaque (Macaca sinica) | [13] | |
| P. silvaticum | ● | Orangutans (Pongo pygmaeus) | [18] | |
| P. youngi | ● | Lar gibbon (Hylobates lar) | [13,19] | |
| America | P. brasilianum | ●● | Capuchin monkey (Cebus albifrons, C. apella, C. tapucinus, C. c. tapucinus, C. c. imitator), Spider monkey (Ateles fusciceps, A. geoffroyi, A. g. geoffroyi, A. g. grisescens, A. panistus, A. p. paniscus, A. p. chamek), Squirrel monkey (Saimiri sciurea, S. boliviensis), Woolly monkey (Lagothrix cana, L. infumata, L. lagotricha), Howler monkey (Alouatta fusca, A. palliata, A. seniculus straminea, A. villosa), Bald uakari (Cacajao calvus), Woolly spider monkey (Brachyteles arachnoides), Titi monkey (Callicebus moloch ornatus, C. torquatus) | [13,14,20] |
| P. simium | ●● | Black howler monkey (Alouatta fusca), Woolly spider monkey (Brachyteles arachnoides), Capuchin monkey (Cebus and Sapajus spp) | [6,13,14,21] | |
| Africa | P. georgesi | ● | Mangabey (Cercocebus albigena, C. galeritus agilis) | [22] |
| P. gonderi | ● | Mangabey (Cercocebus galeritus agilis, C. aterrimus, C. atys), Drill (Mandrillus leucophacus) | [13,14,22] | |
| P. petersi | ● | Mangabey (Cercocebus albigena) | [23] | |
| P. rodhaini | ● | Chimpanzee (Pan troglodytes) | [13] | |
| P. schwetzi | ● | Chimpanzee (Pan troglodytes), Gorilla (Gorilla spp.) | [13,24] |
| Species | Pv P01 | Pv Sal1 | Pc B | Ps Howler | Pk H | Pow_1 | Pow_2 | Poc, Nigeria 1 | Poc_2 | Pm UG01 | Pb Bolivian 1 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Features | ||||||||||||
| Total length (Mp) | 29 | 26.8 | 26.2 | 29 | 24.4 | 35.2 | 35.1 | 34.5 | 38 | 33.6 | 30 | |
| No. of scaffolds | 374 | 374 | 1663 | 2192 | 28 | 1378 | 1593 | 4015 | 2227 | 7270 | 953 | |
| GC% | 39.7 | 42.3 | 40.4 | 44.6 | 37.5 | 28.91 | 29.12 | 28.46 | 27.76 | 24.74 | 24.8 | |
| Total no. of chromosomes | 14 | 14 | 14 | 14 | 14 | 14 | 14 | 14 | 14 | 14 | 14 | |
| No. of protein coding genes | 6642 | 5433 | 5776 | - | 5188 | 8582 | 8813 | 7950 | 8825 | 6540 | 6050 | |
| References | [68,69] | [69] | [70] | [68] | [65,69] | [71] | [71] | [71] | [71] | [69,71] | Accession number: GCA_001885115.2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hang, J.-W.; Tukijan, F.; Lee, E.-Q.-H.; Abdeen, S.R.; Aniweh, Y.; Malleret, B. Zoonotic Malaria: Non-Laverania Plasmodium Biology and Invasion Mechanisms. Pathogens 2021, 10, 889. https://doi.org/10.3390/pathogens10070889
Hang J-W, Tukijan F, Lee E-Q-H, Abdeen SR, Aniweh Y, Malleret B. Zoonotic Malaria: Non-Laverania Plasmodium Biology and Invasion Mechanisms. Pathogens. 2021; 10(7):889. https://doi.org/10.3390/pathogens10070889
Chicago/Turabian StyleHang, Jing-Wen, Farhana Tukijan, Erica-Qian-Hui Lee, Shifana Raja Abdeen, Yaw Aniweh, and Benoit Malleret. 2021. "Zoonotic Malaria: Non-Laverania Plasmodium Biology and Invasion Mechanisms" Pathogens 10, no. 7: 889. https://doi.org/10.3390/pathogens10070889
APA StyleHang, J. -W., Tukijan, F., Lee, E. -Q. -H., Abdeen, S. R., Aniweh, Y., & Malleret, B. (2021). Zoonotic Malaria: Non-Laverania Plasmodium Biology and Invasion Mechanisms. Pathogens, 10(7), 889. https://doi.org/10.3390/pathogens10070889