Abstract
Infectious bursal disease virus is an immunosuppressive pathogen that, despite applied vaccination, is affecting the poultry industry worldwide. This report presents the genetic and pathotypic characterization of a natural reassortant emerging in Europe (Latvia). Genetic characterization showed that strain 25/11/Latvia/2011 represents genotype A3B1, whose segment A is derived from very virulent strains, while segment B is from the classic-like genogroup. Phylogenetic maximum likelihood inference of the B-segment sequence clustered the reassortant strain together with the US antigenic variant E strain. However, the obtained full-length sequence of 25/11/Latvia/2011 revealed that not only reassortment but also dozens of mutations shaped the unique genetic makeup. Phenotypic characterization showed no mortality and no clinical signs of disease but a severe bursa of Fabricius atrophy and splenomegaly in the convalescent birds at 10 days post infection. The results obtained indicate that the acquired genetic constellation contributed to a decrease in virulence; nevertheless, the infection causes severe damage to lymphoid organs, which can lead to impaired immune responses.
1. Introduction
Infectious bursal disease virus (IBDV), a member of the Birnaviridae family, Avibirnavirus genus, is a global pathogen that leads to an immunosuppressive disorder known as Gumboro disease (IBD). The virus has a bisegmented double-stranded RNA genome [1] that encodes five viral proteins. Segment A contains information about the structure of four proteins (VP2-VP5), including the VP2 capsid protein, which is responsible for antigenicity and tissue tropism [2]. The monocistronic segment B encodes viral polymerase (RdRp, VP1) that manages replication, transcription and translation [3,4]. Using reverse genetics, it has been shown that both segments affect virus virulence [5]; however, the precise mechanism of pathogenicity is still unknown. IBDVs can be differentiated based on antigenicity, virulence or genetic composition; although genotype should not be strictly combined with antigenicity or pathotype, they often overlap [6]. Thus, point genome mutations within an antigenic epitope can change the antigenicity of IBDV without significantly changing its position on the phylogenetic tree [7]. Similarly, a mutation in the genome at a virulence-relevant site can alter the pathotype of a virus without changing its genotype [5].
The current global epidemiological situation is shaped by both low-to-moderate [8,9,10,11,12] and highly pathogenic strains [13,14]. In the last decade, a shift has been observed in the population of IBDV genetic lineages circulating in Europe, previously dominated by very virulent strains. Molecular surveys confirmed the emergence of three types of natural reassortants representing genotypes A3B4, A3B1 and A9B1. The first mentioned are prevalent in Poland [12], but their presence has also been confirmed in Finland [15]. Strains of the A3B4 genotype show a pathotype comparable to classical virulent strains, and the infection of SPF chickens resulted in 20-30% mortality [12]. In turn, genotype A3B1 has been confirmed in many European countries including the Netherlands, Belgium, Denmark, Germany, the Czech Republic and Sweden [10]. These strains are less virulent compared to very virulent IBDV, and no mortality or clinical signs were observed in the experimental infection; however, marked atrophy of the bursa of Fabricius indicates major immunosuppressive implications. Moreover, a new genotype A9B1 of IBDV was recently identified in Portugal, but the pathogenicity of the virus has not yet been determined [16].
The presented study reports the genetic and pathotypic characteristics of an IBDV reassortant detected in Latvia, belonging to the A3B1 genotype. Interestingly, full-sequence analysis of the B segment did not confirm phylogenetic relatedness to classical attenuated strains, as in previously reported reassortants in Europe [10], indicating the circulation of more IBDV mosaic groups.
2. Results
2.1. Molecular and Phylogenetic Analysis
The obtained complete consensus sequence of 25/11/Latvia/2011 of both segments was deposited in the GenBank database under accession numbers OP225970 and OP225971. The maximum likelihood (ML) phylogenetic inference based on the full coding sequence confirmed that IBDV strain 25/11/Latvia/2011 is a reassortant whose segment A is derived from very virulent strains (genogroup A3) (Figure 1), while segment B represents the B1 genogroup (Figure 2). The B1 group is the most abundant and includes classical (virulent and attenuated), US antigenic variant, distinct (dIBDV), Italian atypical and serotype 2 strains, as indicated by the calculated nucleotide identity of all full-length sequences deposited in GenBank, which ranges from 92.3 to 100%. However, the 25/11/Latvia/2011 strain forms a monophyletic branch together with the US and Chinese antigenic variants and classical virulent IBDVs (Figure 2). The highest nucleotide similarity for full-length segment A was found with very virulent IBDV strain D6948 (AF240686), which was 97.0%. In contrast, for segment B, 97.0% and 96,9% identity was found with strains GA/1479/2004 (MN814844, classical virulent) and variant E (AF133905, US antigenic variant), respectively. The demonstrated 3% difference in nucleotide similarity for the full sequences of the two segments shows that there are no closely related IBDV strains in the GenBank database. In contrast, partial sequence analysis of VP2 and VP1 showed high nucleotide similarity, 99.5-99.7% and 99.4-99.5%, respectively, with previously reported IBDV strains from Latvia [17]. Amino acid analysis of deduced viral proteins revealed the presence of 18 and 7 altered residues within proteins encoded by segments A and B, compared to strains D6948 and GA/1479/2004 (Table 1). Some of these alterations are unique to the characterized strain, including 5A, 47T and 134N in nonstructural VP5; 545T, 570V and 680F in viral protease VP4; 935V and 960D in structural VP3 protein; and 71D, 157L, 455S and 759R in viral polymerase (VP1).
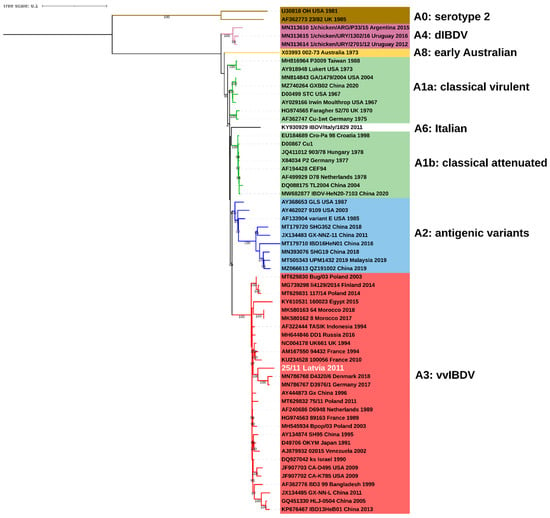
Figure 1.
Phylogenetic tree based on full coding sequence of IBDV for viral segment A (3076nt) using 60 reference strains retrieved from GenBank and maximum likelihood (ML) method, GTR+F+I+G4 model. The main IBDV genogroups according to classification of Islam et al. [18] are denoted (A1: green, A2: blue, A3: red, A4: purple, A6: white, A8: yellow, A0: brown).
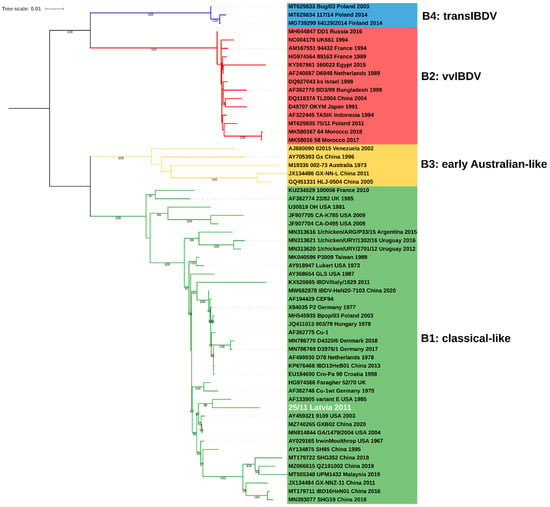
Figure 2.
Phylogenetic tree based on full coding sequence of IBDV for viral segment B (2640nt) using 59 reference strains retrieved from GenBank and maximum likelihood (ML) method, GTR+F+I+G4 model. The main IBDV genogroups according to classification of Islam et al. [18] are denoted (B1: green, B2: red, B3: yellow, B4: blue).

Table 1.
Amino acid differences in viral proteins encoded by both genome segments between 25/11/Latvia/2011 and IBDV representatives of different genogroups.
2.2. Animal Experimental Study
During observation, no mortality or clinical symptoms of the disease were observed in the normal group and in birds infected with the reassortant 25/11/Latvia/2011 (A3B1), while in the very virulent (A3B2) group, birds showed clinical signs such as ruffling of the feather, depression, diarrhea and a 60% mortality rate. At 10 dpi, severe bursa atrophy and splenic enlargement were observed at necropsy in both infected groups, as confirmed by the calculated bursa and spleen development ratios (Figure 3A). At 10 dpi, both groups had significantly lower bursa to body weight (B-BW) ratios (G1 p < 0.005, G2 p < 0.05) compared to the uninfected control group. In contrast, the calculated spleen to body weight (S-BW) ratio was significantly higher only in the reassortant group compared to both the control (p < 0.01) and very virulent groups (p < 0.05) (Figure 3B). Analysis of the mean body weight showed no significant differences in birds after recovery, and the results are shown in Figure 3C. The histopathological examinations of bursal tissue collected from euthanized birds at 10 dpi from both infected groups showed severe lymphoid necrosis due to follicle and lymphocyte depletion and hyperplasia of interfollicular fibrous connective tissue with moderate lymphoid and heterophilic infiltration.
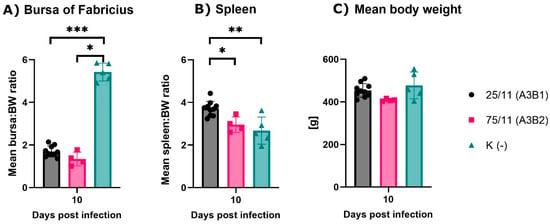
Figure 3.
Mean bursa (A) and spleen (B) to body weight (BW) ratios and mean body weight of experimental groups. Each bar indicates the mean ± SD and statistical significance (* p < 0.05, ** p < 0.01, *** p < 0.001).
3. Discussion
The reassortment phenomenon is a common evolutionary mechanism among RNA viruses with a segmented genome that can lead to a large shift in phenotype, including virulence and transmission efficiency, thus posing a threat to public and animal health [19,20]. As retrospective studies have shown, the IBDV reassortment event was the genesis of a worldwide epidemic of acute Gumboro disease caused by the emergence of very virulent strains [21]. The implementation of routine immunoprophylaxis and the maintenance of biosecurity rules have reduced direct losses in poultry production, but the virus still causes economic losses due to immunosuppression. Thus, continuous field monitoring of important pathogens is key to maintaining the status quo.
Recently, IBDV reassortants of genotype A3B1 were reported in North-West Europe [10,17]. Genetic constellation of these mosaic strains exhibited that they arose from the exchange of segments A and B from very virulent and classical vaccine strains, respectively. As shown by in vivo studies, these strains do not cause mortality and clinical signs in infected SPF chickens but cause severe immunosuppression [10]. Presented genetic studies confirmed that strain 25/11/Latvia/2011 also represents A3B1 genotype reassortants, but the B segment is not vaccine-like but more closely related to classical virulent strains or US antigenic variants, indicating more recombinant virus types in Europe. To date, a similar IBDV reassortant has been detected in China [22]; the SH95 strain also showed the highest similarity of segment B to the variant E strain and segment A to the very virulent strains. Nonetheless, phylogenetic analysis revealed distinct origins of the Latvian and Chinese strains (Figure 2), which was supported by the relatively distant nucleotide identity of 96.4% and 95.9% for the A and B segments, respectively.
The obtained full-length sequence of 25/11/Latvia/2011 revealed that not only reassortment but also mutations shaped the unique genetic makeup. A total of 18 residue changes were found within proteins encoded by segment A compared to the closest reference strain D6948 (Table 1). As many as six amino acid changes were found within the non-structural VP5 protein, which is responsible for the release of virus progeny [23] by regulating the process of apoptosis [24,25] of the infected cell. Two alterations were found within the polycationic C-terminal region (residues 132–143), which is essential for targeting VP5 in the cell plasma membrane and efficient virus dissemination and in the transmembrane domain within the central region of the polypeptide (residues 69–88) [26]. However, the very limited information available on VP5 makes it difficult to conclude whether the changes shown may have affected the function of the protein. Among the changes found in the capsid protein, three are located within the antigenic domains (219L, 220F and 254D), which may contribute to antigenic drift rendering the vaccine less effective [27]. In contrast, the demonstrated replacement of aspartic acid to asparagine at position 279 of the VP2 protein was observed during attenuation of the very virulent OKYM strain [28]. All detected non-synonymous mutations within VP3 were located at the C-terminal part (Table 1). This multifunctional structural protein plays a role as a scaffold for the capsid [29] and viral replication [30]. Using reverse genetics, it was shown that the replacement of the C-terminal part of VP3 of the vvIBDV strain with the corresponding part of the serotype 2 isolate resulted in reduced virulence in SPF chickens [31]. Interestingly, the swapped VP3 fragment differed by six amino acids, and one of them, proline, at position 906 is present in the 25/11/Latvia/2011 strain. Another indicated amino acid substitution, alanine, to valine at position 990 reduced viral replication of the Gt strain [32]. Also within VP4, a serine protease, three altered residues were found; however, no link between these changes and the function of the enzyme was demonstrated so far. The VP1 protein, which is a viral polymerase encoded by segment B, has a number of amino acid changes typical of strains belonging to genogroup B1 (Table 1); in addition, five rare residue replacements were found, including the presence of proline at position 687 typically found in vvIBDV and the B3 genogroup. As indicated, strain 25/11/Latvia/2011 acquired a number of altered amino acids that may affect IBDV antigenicity and replication, or lead to attenuation, but only further study employing reverse genetics will demonstrate the impact of the listed changes. Interestingly, an unusually high number of altered amino acid residues were also observed in the 25/11/Latvia/2011 sequence compared to IBDV found in Europe representing the A3B1 [10] and A3B4 genotypes [12]. It is difficult to determine whether this is the result of an accumulation of mutations or whether we are dealing with the circulation of a distinct genetic lineage of the virus, previously undetected. High nucleotide similarity to Latvian strains (D1526/2/1/10LV, D2323/1/13LV) for VP2 and VP1 partial sequences indicates that this IBDV genetic lineage circulated in Latvia between 2010 and 2013, demonstrating the fitness of the resulting genetic constellation.
Animal trials showed a decrease in the virulence of the tested A3B1 genotype reassortant compared to the very virulent A3B2 strain. Infection with the 25/11/Latvia/2011 strain did not cause mortality or disease symptoms. In contrast, the observed severe lymphoid necrotic lesions in the bursa of Fabricius leading to significant atrophy and splenomegaly at day 10 post infection may indicate that infection can affect the function of these organs, especially the impairment of the host immune response [33]; nevertheless, further studies should be conducted to demonstrate their importance in this process. Loss of IBDV strain virulence resulting from reassortment has been frequently reported, specifically when a very virulent A segment is combined with a B segment of various origins, including vaccine-like (B1 genogroup) [10,34], serotype 2 (B1) [35], early Australian-like (B3) [36] or transitional IBDV (B4) [12]. In contrast, viruses belonging to the A3B1 genotype have also been reported to retain virulence, causing high mortality rates, such as the reassortants detected in Algeria [37] and South Korea [38]. Escaffre et al. (2013) indicated that both genome segments contribute to the virulence of very virulent strains [5], but given the lack of knowledge regarding the precise mechanisms of IBDV virulence, it cannot be ruled out that both the acquired B segment and the observed changes in viral proteins affect the pathogenicity of the detected strain. Nevertheless, only undertaking studies using reverse genetics would clarify the contribution of particular changes in the strain’s genome to the virulence process. Unfortunately, the pathogenicity of SH95, the only strain with a similar genetic composition, has not been studied.
Epidemiological data on the incidence of Gumboro disease show that IBDV reassortants are increasingly being isolated from field cases [10,39,40,41], indicating that the virus is continuously evolving, seeking new characteristics to facilitate its persistence in the environment. Strain 25/11/Latvia/2011 is a good example of this trend; its reduced virulence results in a subclinical course of the disease. Detection of such viruses is more difficult because the observed symptoms are nonspecific or less visible, which promotes the spread of the virus in the field. Thus, it is extremely important to conduct continuous field monitoring of IBDV using molecular methods based on both segments and to tighten biosecurity standards in case of field strain emerges. From a practical point of view, it would also be very important to determine whether the identified amino acid changes within the antigenic epitopes can affect the effectiveness of the vaccines used.
4. Materials and Methods
4.1. Full-length Genome Sequencing
In the present study, bursal tissue from a reported case of Gumboro disease [42] was used and prepared for high-throughput sequencing as previously described [43]. Sequencing was performed using the MiSeq System (Ilumina, San Diego, CA, USA) with 2 × 250 paired-end reads by commercial service (Genomed, Warsaw, Poland). CLC Genomics Workbench v7.0.4 was used for all bioinformatics analyses.
4.2. Phylogenetic Analysis
Sixty full coding sequences of IBDV reference strains downloaded from GenBank were included in the analyses (Table 2). Sequences of recent A3B1 European reassortants were also used for partial VP2 and VP1 analysis. The alignment of all sequence sets was performed using ClustalW method implemented in MEGAX program [44]. IQ-TREE software (version 1.6.12) was used both to estimate the best evolutionary model and to infer phylogenetic trees by maximum likelihood algorithm, and confidence levels for the branches were determined by Shimodaira–Hasegawa test and 1000 replicates of bootstrap [45]. The nucleotide identity was assessed using Geneious Prime 2022.1.1. The tree visualization was performed using the iTOL v6 online tool [46].
Table 2.
List of IBDV reference strains used for genetic analysis.
4.3. Animal Experimental Study
Animal experiments were carried out in accordance with the requirements and authorization of the local ethical commission (Permit no. 79/2015). A total of 25 five-week-old SPF chickens (VALO BioMedia, Osterholz-Scharmbeck, Germany) were randomly divided into 3 experimental groups: 10 birds in groups 1 and 2 and 5 birds in group 3. Chickens of groups 1 (G1) and 2 (G2) were intraocularly inoculated with 105 EID50/0,1ml of the reassortant 25/11/Latvia/2011 (A3B1) and very virulent IBDV (A3B2) reference strain 75/11/Poland/2011, respectively, whereas birds from group 3 served as normal control and were inoculated with the same volume of phosphate-buffered saline. Chickens from all groups were separately housed in HEPA-filtered isolators (Montair Andersen B.V., Kronenberg, Netherlands) with access to water and feed ad libitum and monitored daily for 10 days for clinical symptoms. At the end of the experiment, all surviving birds were euthanized, and bursas of Fabricius and spleens were taken for further investigations. At 10 dpi, the euthanized birds, bursas and spleens were weighed for calculation of ratios of bursa and spleen to body weight according to previously published formula [47]. Half of each bursa was fixed in 10% phosphate-buffered formalin for HE staining and histopathological examination as previously described [43].
4.4. Statistical Analysis
The obtained data were analyzed using Graphpad Prism software 9.4.1 (Graphpad Prism Software Inc., San Diego, CA, USA). The mean with SD of ratios of bursa or spleen to body weight (B-BW and S-BW) and mean body weight calculated for each bird from the experimental groups were compared using the Mann–Whitney test. The results with p < 0.05 were considered statistically significant.
Author Contributions
Conceptualization, A.P.; methodology, A.P.; software, A.P.; validation, A.P. and A.L.; formal analysis, A.P. and A.L.; investigation, A.P. and A.L.; data curation A.P. and A.L.; writing—original draft preparation, A.P.; writing—review and editing, A.P. and A.L.; visualization, A.P.; supervision, A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The animal study protocol was approved by the Local Ethical Commission in Lublin (Permit no. 79/2015) in agreement with the rules in place in the EU (Directive 2010/63/UE).
Data Availability Statement
The complete genome sequences generated in this study were submitted to the GenBank database (https://www.ncbi.nlm.nih.gov/genbank/; accessed on 16 August 2022) under accession numbers OP225970 and OP225971.
Acknowledgments
We thank Agnieszka Jasik and Paulina Sadurska for excellent technical assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dobos, P.; Hill, B.J.; Hallett, R.; Kells, D.T.; Becht, H.; Teninges, D. Biophysical and biochemical characterization of five animal viruses with bisegmented double-stranded RNA genomes. J. Virol. 1979, 32, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Coulibaly, F.; Chevalier, C.; Gutsche, I.; Pous, J.; Navaza, J.; Bressanelli, S.; Delmas, B.; Rey, F.A. The birnavirus crystal structure reveals structural relationships among icosahedral viruses. Cell 2005, 120, 761–772. [Google Scholar] [CrossRef] [PubMed]
- von Einem, U.I.; Gorbalenya, A.E.; Schirrmeier, H.; Behrens, S.E.; Letzel, T.; Mundt, E. VP1 of infectious bursal disease virus is an RNA-dependent RNA polymerase. J. Gen. Virol. 2004, 85, 2221–2229. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Wang, Y.; Zhang, E.; Han, X.; Yu, Z.; Liu, H. VP1 and VP3 Are Required and Sufficient for Translation Initiation of Uncapped Infectious Bursal Disease Virus Genomic Double-Stranded RNA. J. Virol. 2018, 92, e01345-17. [Google Scholar] [CrossRef]
- Escaffre, O.; Le Nouen, C.; Amelot, M.; Ambroggio, X.; Ogden, K.M.; Guionie, O.; Toquin, D.; Muller, H.; Islam, M.R.; Eterradossi, N. Both Genome Segments Contribute to the Pathogenicity of Very Virulent Infectious Bursal Disease Virus. J. Virol. 2013, 87, 2767–2780. [Google Scholar] [CrossRef]
- Eterradossi, N.; Saif, Y.M. Infectious bursal disease. In Diseases of Poultry, 14th ed.; Swayne, D.E., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 257–283. [Google Scholar] [CrossRef]
- Eterradossi, N.; Arnauld, C.; Toquin, D.; Rivallan, G. Critical amino acid changes in VP2 variable domain are associated with typical and atypical antigenicity in very virulent infectious bursal disease viruses. Arch. Virol. 1998, 143, 1627–1636. [Google Scholar] [CrossRef]
- Fan, L.J.; Wu, T.T.; Wang, Y.L.; Hussain, A.; Jiang, N.; Gao, L.; Li, K.; Gao, Y.L.; Liu, C.J.; Cui, H.Y.; et al. Novel variants of infectious bursal disease virus can severely damage the bursa of fabricius of immunized chickens. Vet. Microbiol. 2020, 240, 108507. [Google Scholar] [CrossRef] [PubMed]
- Myint, O.; Suwanruengsri, M.; Araki, K.; Izzati, U.Z.; Pornthummawat, A.; Nueangphuet, P.; Fuke, N.; Hirai, T.; Jackwood, D.J.; Yamaguchi, R. Bursa atrophy at 28 days old caused by variant infectious bursal disease virus has a negative economic impact on broiler farms in Japan. Avian Pathol. 2021, 50, 6–17. [Google Scholar] [CrossRef]
- Mato, T.; Tatar-Kis, T.; Felfoldi, B.; Jansson, D.S.; Homonnay, Z.; Banyai, K.; Palya, V. Occurrence and spread of a reassortant very virulent genotype of infectious bursal disease virus with altered VP2 amino acid profile and pathogenicity in some European countries. Vet. Microbiol. 2020, 245, 108663. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, N.; Fan, L.; Niu, X.; Zhang, W.; Huang, M.; Gao, L.; Li, K.; Gao, Y.; Liu, C.; et al. Identification and Pathogenicity Evaluation of a Novel Reassortant Infectious Bursal Disease Virus (Genotype A2dB3). Viruses 2021, 13, 1682. [Google Scholar] [CrossRef]
- Pikuła, A.; Lisowska, A.; Jasik, A.; Perez, L.J. The Novel Genetic Background of Infectious Bursal Disease Virus Strains Emerging from the Action of Positive Selection. Viruses 2021, 13, 396. [Google Scholar] [CrossRef]
- Abbas, A.H.; Al Saegh, H.A.; FS, A.L. Sequence diversity and evolution of infectious bursal disease virus in Iraq. F1000Research 2021, 10, 293. [Google Scholar] [CrossRef] [PubMed]
- Drissi Touzani, C.; Fellahi, S.; Fihri, O.F.; Gaboun, F.; Khayi, S.; Mentag, R.; Lico, C.; Baschieri, S.; El Houadfi, M.; Ducatez, M. Complete genome analysis and time scale evolution of very virulent infectious bursal disease viruses isolated from recent outbreaks in Morocco. Infect. Genet. Evol. 2020, 77, 104097. [Google Scholar] [CrossRef]
- Tammiranta, N.; Ek-Kommonen, C.E.; Rossow, L.; Huovilainen, A. Circulation of very virulent avian infectious bursal disease virus in Finland. Avian Pathol. 2018, 47, 520–525. [Google Scholar] [CrossRef]
- Legnardi, M.; Franzo, G.; Tucciarone, C.M.; Koutoulis, K.; Duarte, I.; Silva, M.; Le Tallec, B.; Cecchinato, M. Detection and molecular characterization of a new genotype of infectious bursal disease virus in Portugal. Avian Pathol. 2022, 51, 97–105. [Google Scholar] [CrossRef]
- Pikula, A.; Domanska-Blicharz, K.; Cepulis, R.; Smietanka, K. Identification of infectious bursal disease virus with atypical VP2 amino acid profile in Latvia. J. Vet. Res. 2017, 61, 145–149. [Google Scholar] [CrossRef]
- Mato, T.; Medveczki, A.; Kiss, I. Research Note: "Hidden" infectious bursal disease virus infections in Central Europe. Poult. Sci. 2022, 101, 101958. [Google Scholar] [CrossRef]
- Islam, M.R.; Nooruzzaman, M.; Rahman, T.; Mumu, T.T.; Rahman, M.M.; Chowdhury, E.H.; Eterradossi, N.; Muller, H. A unified genotypic classification of infectious bursal disease virus based on both genome segments. Avian Pathol. 2021, 50, 190–206. [Google Scholar] [CrossRef]
- Gerrard, S.R.; Li, L.; Barrett, A.D.; Nichol, S.T. Ngari virus is a Bunyamwera virus reassortant that can be associated with large outbreaks of hemorrhagic fever in Africa. J. Virol. 2004, 78, 8922–8926. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.I.; Viboud, C.; Simonsen, L.; Bennett, R.T.; Griesemer, S.B.; George, K.S.; Taylor, J.; Spiro, D.J.; Sengamalay, N.A.; Ghedin, E.; et al. Multiple reassortment events in the evolutionary history of H1N1 influenza A virus since 1918. PLoS Pathog. 2008, 4, e1000012. [Google Scholar] [CrossRef]
- Hon, C.C.; Lam, T.Y.; Drummond, A.; Rambaut, A.; Lee, Y.F.; Yip, C.W.; Zeng, F.; Lam, P.Y.; Ng, P.T.; Leung, F.C. Phylogenetic analysis reveals a correlation between the expansion of very virulent infectious bursal disease virus and reassortment of its genome segment B. J. Virol. 2006, 80, 8503–8509. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.H.; Lu, P.; Yan, Y.X.; Hua, X.G.; Jiang, J.; Zhao, Y. Sequence and analysis of genomic segment A and B of very virulent infectious bursal disease virus isolated from China. J. Vet. Med. Ser. B 2003, 50, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.P.; Hong, L.L.; Ye, J.X.; Huang, Z.Y.; Zhou, J.Y. The VP5 protein of infectious bursal disease virus promotes virion release from infected cells and is not involved in cell death. Arch. Virol. 2009, 154, 1873–1882. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Hou, L.; Zhu, S.; Wang, J.; Zhou, J.; Liu, J. Infectious bursal disease virus activates the phosphatidylinositol 3-kinase (PI3K)/Akt signaling pathway by interaction of VP5 protein with the p85alpha subunit of PI3K. Virology 2011, 417, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, Y.; Xue, Y.; Li, X.; Cao, H.; Zheng, S.J. Critical role for voltage-dependent anion channel 2 in infectious bursal disease virus-induced apoptosis in host cells via interaction with VP5. J. Virol. 2012, 86, 1328–1338. [Google Scholar] [CrossRef] [PubMed]
- Mendez, F.; de Garay, T.; Rodriguez, D.; Rodriguez, J.F. Infectious bursal disease virus VP5 polypeptide: A phosphoinositide-binding protein required for efficient cell-to-cell virus dissemination. PLoS ONE 2015, 10, e0123470. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, D.J.; Sommer-Wagner, S.E. Amino acids contributing to antigenic drift in the infectious bursal disease Birnavirus (IBDV). Virology 2011, 409, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Ogawa, M.; Inoshima, Y.; Miyoshi, M.; Fukushi, H.; Hirai, K. Identification of sequence changes responsible for the attenuation of highly virulent infectious bursal disease virus. Virology 1996, 223, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Mata, C.P.; Mertens, J.; Fontana, J.; Luque, D.; Allende-Ballestero, C.; Reguera, D.; Trus, B.L.; Steven, A.C.; Carrascosa, J.L.; Caston, J.R. The RNA-Binding Protein of a Double-Stranded RNA Virus Acts like a Scaffold Protein. J. Virol. 2018, 92, e00968-18. [Google Scholar] [CrossRef] [PubMed]
- Tacken, M.G.; Peeters, B.P.; Thomas, A.A.; Rottier, P.J.; Boot, H.J. Infectious bursal disease virus capsid protein VP3 interacts both with VP1, the RNA-dependent RNA polymerase, and with viral double-stranded RNA. J. Virol. 2002, 76, 11301–11311. [Google Scholar] [CrossRef]
- Boot, H.J.; ter Huurne, A.A.; Hoekman, A.J.; Pol, J.M.; Gielkens, A.L.; Peeters, B.P. Exchange of the C-terminal part of VP3 from very virulent infectious bursal disease virus results in an attenuated virus with a unique antigenic structure. J. Virol. 2002, 76, 10346–10355. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, Y.Q.; Qi, X.L.; Kang, Z.H.; Yu, F.; Qin, L.T.; Gao, H.L.; Gao, Y.L.; Wang, X.M. A single amino acid in the C-terminus of VP3 protein influences the replication of attenuated infectious bursal disease virus in vitro and in vivo. Antivir. Res. 2010, 87, 223–229. [Google Scholar] [CrossRef]
- Trapp, J.; Rautenschlein, S. Infectious bursal disease virus’ interferences with host immune cells: What do we know? Avian Pathol. 2022, 51, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Zhang, L.; Wang, N.; Chen, Y.; Gao, L.; Wang, Y.; Gao, H.; Gao, Y.; Li, K.; Qi, X.; et al. Naturally occurring reassortant infectious bursal disease virus in northern China. Virus Res. 2015, 203, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Soubies, S.M.; Courtillon, C.; Briand, F.X.; Queguiner-Leroux, M.; Courtois, D.; Amelot, M.; Grousson, K.; Morillon, P.; Herin, J.B.; Eterradossi, N. Identification of a European interserotypic reassortant strain of infectious bursal disease virus. Avian Pathol. 2017, 46, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Le Nouen, C.; Rivallan, G.; Toquin, D.; Darlu, P.; Morin, Y.; Beven, V.; de Boisseson, C.; Cazaban, C.; Comte, S.; Gardin, Y.; et al. Very virulent infectious bursal disease virus: Reduced pathogenicity in a rare natural segment-B-reassorted isolate. J. Gen. Virol. 2006, 87, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Abed, M.; Soubies, S.; Courtillon, C.; Briand, F.X.; Allee, C.; Amelot, M.; De Boisseson, C.; Lucas, P.; Blanchard, Y.; Belahouel, A.; et al. Infectious bursal disease virus in Algeria: Detection of highly pathogenic reassortant viruses. Infect. Genet. Evol. 2018, 60, 48–57. [Google Scholar] [CrossRef]
- Lee, H.J.; Jang, I.; Shin, S.H.; Lee, H.S.; Choi, K.S. Genome Sequence of a Novel Reassortant and Very Virulent Strain of Infectious Bursal Disease Virus. Genome Announc. 2017, 5, e00730-17. [Google Scholar] [CrossRef]
- Pikuła, A.; Śmietanka, K.; Perez, L.J. Emergence and expansion of novel pathogenic reassortant strains of infectious bursal disease virus causing acute outbreaks of the disease in Europe. Transbound. Emerg. Dis. 2020, 67, 1739–1744. [Google Scholar] [CrossRef]
- Jiang, N.; Wang, Y.; Zhang, W.; Niu, X.; Huang, M.; Gao, Y.; Liu, A.; Gao, L.; Li, K.; Pan, Q.; et al. Genotyping and Molecular Characterization of Infectious Bursal Disease Virus Identified in Important Poultry-Raising Areas of China During 2019 and 2020. Front. Vet. Sci. 2021, 8, 759861. [Google Scholar] [CrossRef]
- Arowolo, O.A.; George, U.E.; Luka, P.D.; Maurice, N.A.; Atuman, Y.J.; Shallmizhili, J.J.; Shittu, I.; Oluwayelu, D.O. Infectious bursal disease in Nigeria: Continuous circulation of reassortant viruses. Trop. Anim. Health Prod. 2021, 53, 271. [Google Scholar] [CrossRef] [PubMed]
- Pikuła, A.; Lisowska, A.; Jasik, A.; Śmietanka, K. Identification and assessment of virulence of a natural reassortant of infectious bursal disease virus. Vet. Res. 2018, 49, 89. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Jackwood, D.J.; Sommer-Wagner, S.E.; Crossley, B.M.; Stoute, S.T.; Woolcock, P.R.; Charlton, B.R. Identification and pathogenicity of a natural reassortant between a very virulent serotype 1 infectious bursal disease virus (IBDV) and a serotype 2 IBDV. Virology 2011, 420, 98–105. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).