

Micronutrients Affect Expression of Induced Resistance Genes in Hydroponically Grown Watermelon against Fusarium oxysporum f. sp. niveum and Meloidogyne incognita

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Mineral Content in Nutrient Solutions
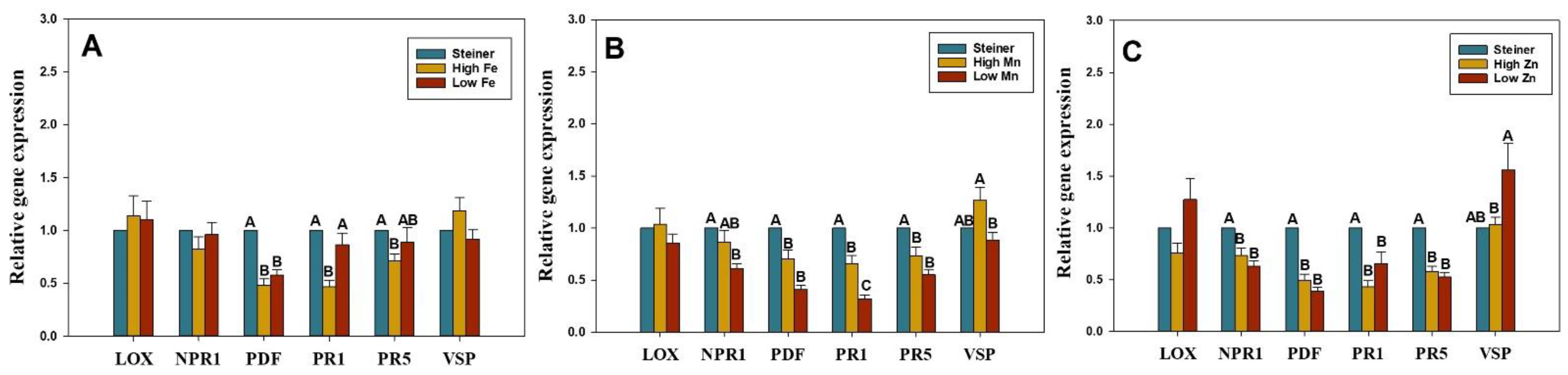
2.2. Change in Micronutrient Level Influences the Expression of SA- and JA-Genes in Watermelon Seedlings
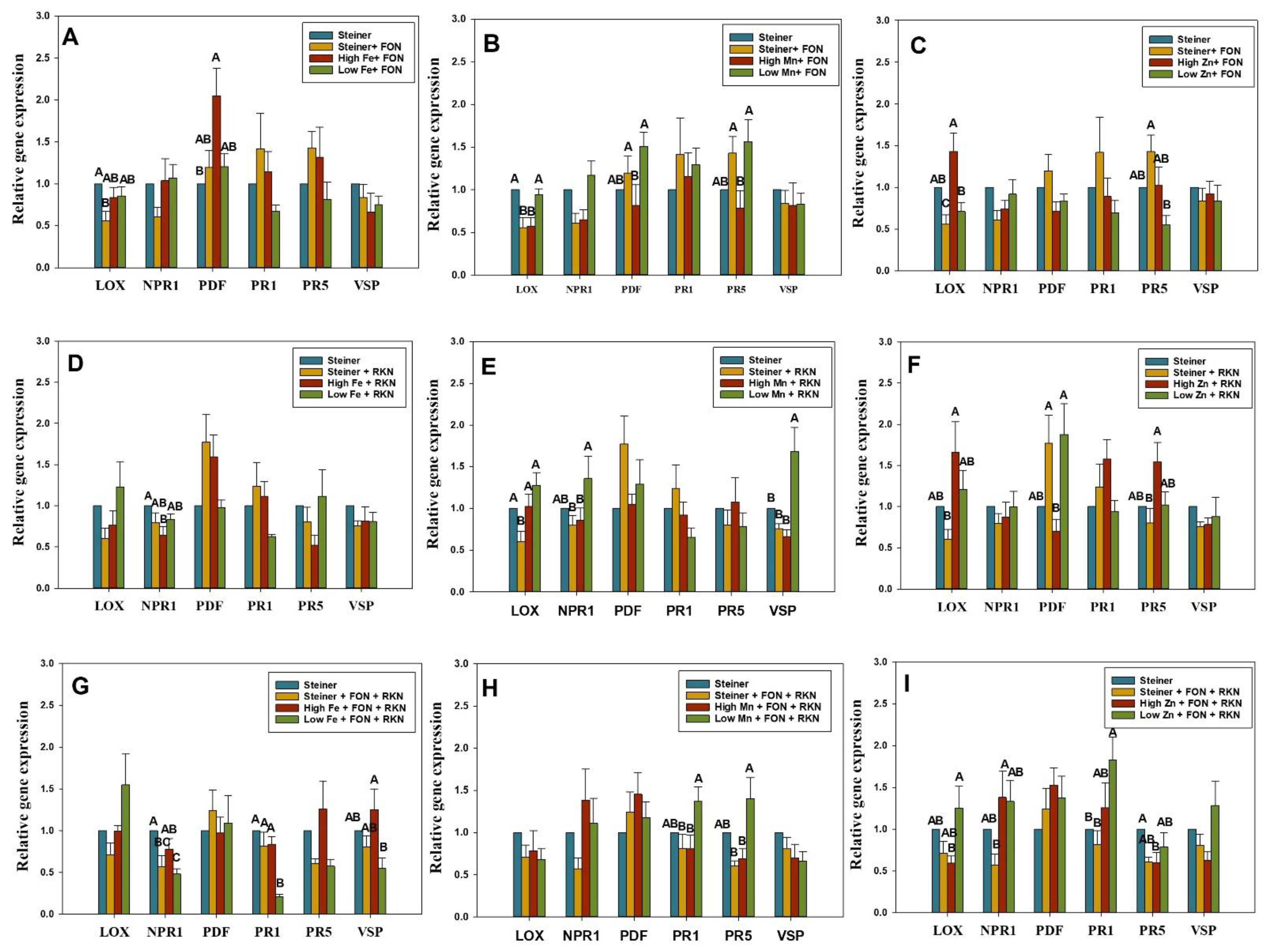
2.3. Differential Expression of SA- and JA-Genes in Fe-Treated Plants Inoculated with FON, RKN, or Both
2.4. Differential Expression of SA- and JA-Genes in Mn-Treated Plants Inoculated with FON, RKN, or Both
2.5. Differential Expression of SA- and JA-Genes in Zn-Treated Plants Inoculated with FON, RKN, or Both
2.6. FON Recovery and RKN Gall Rating
3. Discussion
4. Materials and Methods
4.1. Preparation of Inoculum of FON and RKN
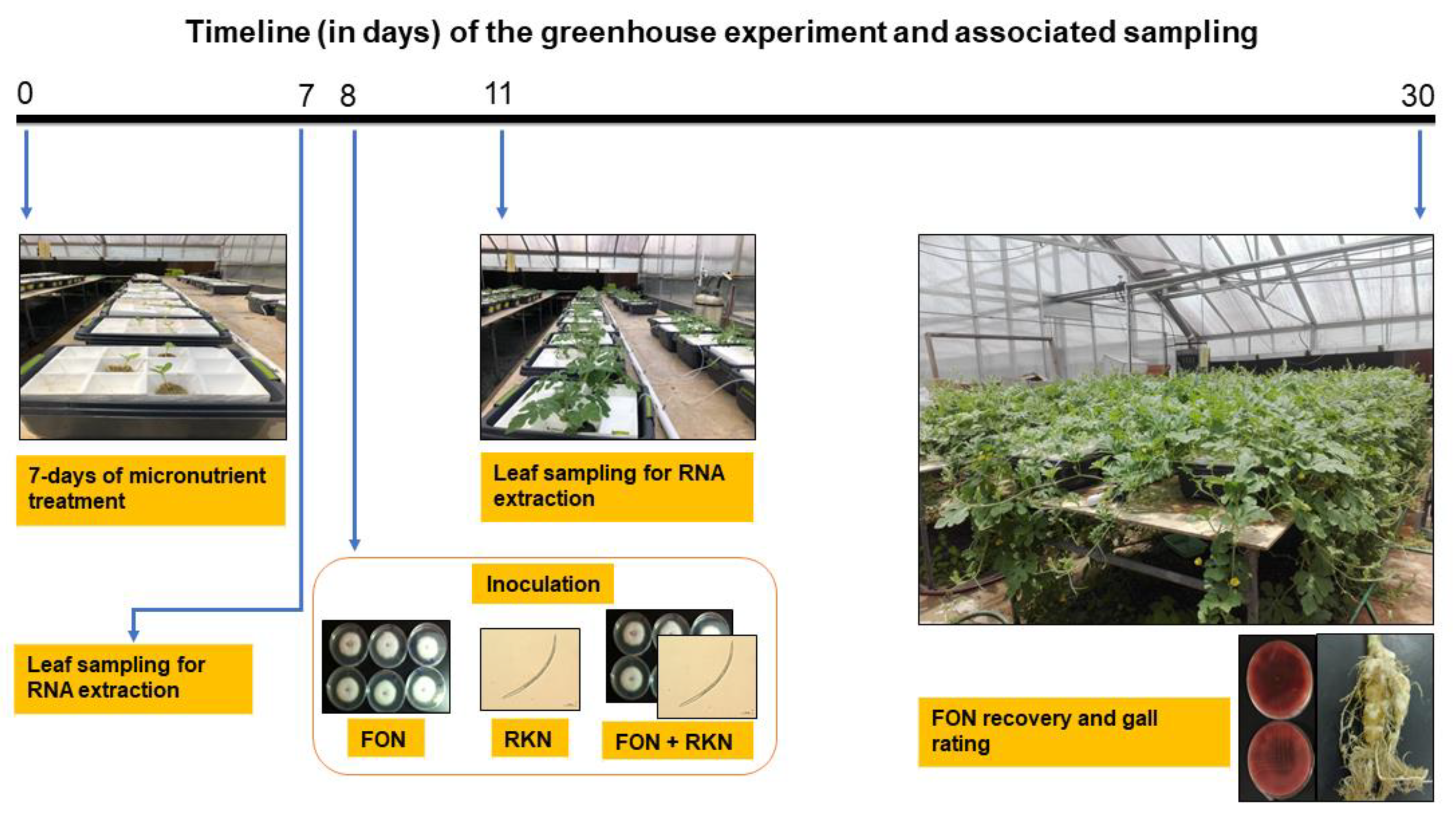
4.2. Hydroponics Set-Up under Greenhouse Conditions
4.3. Mineral Analysis
4.4. Relative Expression of SA- and JA-Genes in Watermelon Seedlings Grown in Specific Micronutrient Solutions
4.5. Relative Expression of SA- and JA-Genes in Watermelon Seedlings, Grown in Specific Micronutrient Solutions and Inoculated with FON, RKN, or Both
4.6. FON Recovery and RKN Gall Rating
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. FAO Countries by Commodities–Top Yield—Watermelons; FAO: Rome, Italy, 2018. [Google Scholar]
- USDA National Agricultural Statistics Service. USDA-NASS Quick Stats; USDA National Agricultural Statistics Service: Washington, DC, USA, 2019. [Google Scholar]
- Ali, A.; Abdalla, O.; Bruton, B.; Fish, W.; Sikora, E.; Zhang, S.; Taylor, M. Occurrence of Viruses Infecting Watermelon, Other Cucurbits, and Weeds in the Parts of Southern United States. Plant Health Prog. 2012, 13, 9. [Google Scholar] [CrossRef]
- Walsh, K.A.; Bennett, S.D.; Mahovic, M.; Gould, L.H. Outbreaks Associated with Cantaloupe, Watermelon, and Honeydew in the United States, 1973–2011. Foodborne Pathog. Dis. 2014, 11, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Schneider, S.M.; Rosskopf, E.N.; Leesch, J.G.; Chellemi, D.O.; Bull, C.T.; Mazzola, M. United States Department of Agriculture—Agricultural Research Service Research on Alternatives to Methyl Bromide: Pre-Plant and Post-Harvest. Pest Manag. Sci. 2003, 59, 814–826. [Google Scholar] [CrossRef]
- Thies, J.A.; Ariss, J.J.; Hassell, R.L.; Buckner, S.; Levi, A. Accessions of Citrullus lanatus var. citroides Are Valuable Rootstocks for Grafted Watermelon in Fields Infested with Root-Knot Nematodes. HortScience 2015, 50, 4–8. [Google Scholar] [CrossRef]
- Martyn, R.D. Fusarium Wilt of Watermelon: 120 Years of Research. In Horticultural Reviews: Volume 42; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014; pp. 349–442. ISBN 978-1-118-91682-7. [Google Scholar]
- Hua, G.K.H.; Timper, P.; Ji, P. Meloidogyne incognita Intensifies the Severity of Fusarium Wilt on Watermelon Caused by Fusarium oxysporum f. sp. niveum. Can. J. Plant Pathol. 2019, 41, 261–269. [Google Scholar] [CrossRef]
- Petkar, A.; Ji, P. Infection Courts in Watermelon Plants Leading to Seed Infestation by Fusarium oxysporum f. sp. niveum. Phytopathology 2017, 107, 828–833. [Google Scholar] [CrossRef]
- Martyn, R.D. Fusarium Wilt of Watermelon. Compend. Cucurbit Dis. 1996, 13–14. [Google Scholar]
- Zhang, M.; Xu, J.H.; Liu, G.; Yao, X.F.; Li, P.F.; Yang, X.P. Characterization of the Watermelon Seedling Infection Process by Fusarium oxysporum f. sp. niveum. Plant Pathol. 2015, 64, 1076–1084. [Google Scholar] [CrossRef]
- Cirulli, M. Variation of Pathogenicity in Fusarium oxysporum f. sp. niveum and Resistance in Watermelon Cultivars. Actas Congr. Union Fitopathol. Mediter. Oeiras 1972, 3, 491–500. [Google Scholar]
- Netzer, D. Physiological Races and Soil Population Level of Fusarium Wilt of Watermelon. Phytoparasitica 1976, 4, 131–136. [Google Scholar] [CrossRef]
- Bruton, B.D.; Damicone, J.P. Fusarium Wilt of Watermelon: Impact of Race 2 of Fusarium oxysporum f. sp. niveum on Watermelon Production in Texas And. Subtrop. Plant Sci. 1999, 51, 4–9. [Google Scholar]
- Zhou, X.G.; Everts, K.L. Races and Inoculum Density of Fusarium oxysporum f. sp. niveum in Commercial Watermelon Fields in Maryland and Delaware. Plant Dis. 2003, 87, 692–698. [Google Scholar] [PubMed]
- Zhou, X.G.; Everts, K.L.; Bruton, B.D. Race 3, a New and Highly Virulent Race of Fusarium oxysporum f. sp. niveum Causing Fusarium Wilt in Watermelon. Plant Dis. 2010, 94, 92–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaradasa, B.S.; Beckham, K.; Dufault, N.; Sanchez, T.; Ertek, T.S.; Iriarte, F.; Paret, M.; Ji, P. First Report of Fusarium oxysporum f. sp. niveum Race 3 Causing Wilt of Watermelon in Florida, U.S.A. Plant Dis. 2018, 102, 1029. [Google Scholar] [CrossRef]
- Egel, D.S.; Hoke, S.; Cardinal, C. Evaluation of Triploid Watermelon for Resistance to Fusarium Wilt, 2010. Plant Manag. Rep. 2010, 4, V135. [Google Scholar]
- Yetışır, H.; Sari, N.; Yücel, S. Rootstock Resistance to Fusarium Wilt and Effect on Watermelon Fruit Yield and Quality. Phytoparasitica 2003, 31, 163–169. [Google Scholar] [CrossRef]
- Keinath, A.P.; Hassell, R.L. Control of Fusarium Wilt of Watermelon by Grafting onto Bottlegourd or Interspecific Hybrid Squash Despite Colonization of Rootstocks by Fusarium. Plant Dis. 2014, 98, 255–266. [Google Scholar] [CrossRef]
- Keinath, A.P.; Hassell, R.L. Suppression of Fusarium Wilt Caused by Fusarium oxysporum f. sp. niveum Race 2 on Grafted Triploid Watermelon. Plant Dis. 2014, 98, 1326–1332. [Google Scholar] [CrossRef]
- Sumner, D.R.; Johnson, A.W. Effect of Root-Knot Nemotodes on Fusarium Wilt of Watermelon. Phytopathology 1973, 63, 857–861. [Google Scholar] [CrossRef]
- Davis, R.F. Effect of Meloidogyne incognita on Watermelon Yield. Nematropica 2007, 37, 287–294. [Google Scholar]
- Jepson, S.B. Identification of Root-Knot Nematodes (Meloidogyne Species); CAB International: Wallingford, UK, 1987. [Google Scholar]
- Dale, P.S. Elimination of Root-Knot Nematodes from Roses by Chemical Bare-Root Dips. N. Z. J. Exp. Agric. 1973, 1, 121–122. [Google Scholar] [CrossRef]
- Newton, A.C.; Torrance, L.; Holden, N.; Toth, I.K.; Cooke, D.E.L.; Blok, V.; Gilroy, E.M. Chapter Three—Climate Change and Defense against Pathogens in Plants. In Advances in Applied Microbiology; Gadd, G.M., Sariaslani, S., Eds.; Academic Press: Cambridge, MA, USA, 2012; Volume 81, pp. 89–132. [Google Scholar]
- Hajihassani, A.; Forghani, F.; Marquez, J. Managing Root-Knot Nematodes in Georgia Watermelons|UGA Cooperative Extension. Available online: https://extension.uga.edu/publications/detail.html?number=B1529 (accessed on 7 June 2022).
- Lynch, L.; Carpenter, J. The Economic Impacts of Banning Methyl Bromide: Where Do We Need More Research. In Proceedings of the 1999 Annual Research Conference on Methyl Bromide Alternatives and Emissions Reductions; UNEP: Paris, France, 1999. [Google Scholar]
- Thies, J.A.; Ariss, J.J.; Hassell, R.L.; Olson, S.; Kousik, C.S.; Levi, A. Grafting for Management of Southern Root-Knot Nematode, Meloidogyne incognita, in Watermelon. Plant Dis. 2010, 94, 1195–1199. [Google Scholar] [CrossRef] [PubMed]
- Thies, J.A.; Ariss, J.J.; Kousik, C.S.; Hassell, R.L.; Levi, A. Resistance to Southern Root-Knot Nematode (Meloidogyne incognita) in Wild Watermelon (Citrullus lanatus var. citroides). J. Nematol. 2016, 48, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.; Kim, Y.H. Pathological Interrelations of Soil-Borne Diseases in Cucurbits Caused by Fusarium Species and Meloidogyne incognita. Plant Pathol. J. 2017, 33, 410–423. [Google Scholar] [CrossRef]
- Keinath, A.P.; Agudelo, P.A. Retention of Resistance to Fusarium oxysporum f. sp. niveum in Cucurbit Rootstocks Infected by Meloidogyne incognita. Plant Dis. 2018, 102, 1820–1827. [Google Scholar] [CrossRef] [Green Version]
- Keinath, A.P.; Coolong, T.W.; Lanier, J.D.; Ji, P. Managing Fusarium Wilt of Watermelon with Delayed Transplanting and Cultivar Resistance. Plant Dis. 2019, 103, 44–50. [Google Scholar] [CrossRef]
- Siddiqui, I.A.; Shaukat, S.S. Rhizobacteria-Mediated Induction of Systemic Resistance (ISR) in Tomato against Meloidogyne javanica. J. Phytopathol. 2002, 150, 469–473. [Google Scholar] [CrossRef]
- Chinnasri, B.; Sipes, B.S.; Schmitt, D.P. Effects of Inducers of Systemic Acquired Resistance on Reproduction of Meloidogyne javanica and Rotylenchulus reniformis in Pineapple. J. Nematol. 2006, 38, 319–325. [Google Scholar]
- Jiang, C.-H.; Yao, X.-F.; Mi, D.-D.; Li, Z.-J.; Yang, B.-Y.; Zheng, Y.; Qi, Y.-J.; Guo, J.-H. Comparative Transcriptome Analysis Reveals the Biocontrol Mechanism of Bacillus velezensis F21 against Fusarium Wilt on Watermelon. Front. Microbiol. 2019, 10, 652. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, C.; Ahammed, G.J.; Wu, C.; Yang, Z.; Wan, C.; Chen, J. Red Light-Induced Systemic Resistance against Root-Knot Nematode Is Mediated by a Coordinated Regulation of Salicylic Acid, Jasmonic Acid and Redox Signaling in Watermelon. Front. Plant Sci. 2018, 9, 899. [Google Scholar] [CrossRef]
- Niki, T.; Mitsuhara, I.; Seo, S.; Ohtsubo, N.; Ohashi, Y. Antagonistic Effect of Salicylic Acid and Jasmonic Acid on the Expression of Pathogenesis-Related (PR) Protein Genes in Wounded Mature Tobacco Leaves. Plant Cell Physiol. 1998, 39, 500–507. [Google Scholar] [CrossRef]
- Cao, H.; Bowling, S.A.; Gordon, A.S.; Dong, X. Characterization of an Arabidopsis Mutant That Is Nonresponsive to Inducers of Systemic Acquired Resistance. Plant Cell 1994, 6, 1583–1592. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.Q.; Yan, S.; Saleh, A.; Wang, W.; Ruble, J.; Oka, N.; Mohan, R.; Spoel, S.H.; Tada, Y.; Zheng, N.; et al. NPR3 and NPR4 Are Receptors for the Immune Signal Salicylic Acid in Plants. Nature 2012, 486, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, D.; Chu, J.Y.; Boyle, P.; Wang, Y.; Brindle, I.D.; De Luca, V.; Després, C. The Arabidopsis NPR1 Protein Is a Receptor for the Plant Defense Hormone Salicylic Acid. Cell Rep. 2012, 1, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Delaney, T.P.; Friedrich, L.; Ryals, J.A. Arabidopsis Signal Transduction Mutant Defective in Chemically and Biologically Induced Disease Resistance. Proc. Natl. Acad. Sci. USA 1995, 92, 6602–6606. [Google Scholar] [CrossRef]
- Benedetti, C.E.; Xie, D.; Turner, J.G. COI1-Dependent Expression of an Arabidopsis Vegetative Storage Protein in Flowers and Siliques and in Response to Coronatine or Methyl Jasmonate. Plant Physiol. 1995, 109, 567–572. [Google Scholar] [CrossRef] [Green Version]
- Heitz, T.; Bergey, D.R.; Ryan, C.A. A Gene Encoding a Chloroplast-Targeted Lipoxygenase in Tomato Leaves Is Transiently Induced by Wounding, Systemin, and Methyl Jasmonate. Plant Physiol. 1997, 114, 1085–1093. [Google Scholar] [CrossRef]
- Penninckx, I.A.M.A.; Thomma, B.P.H.J.; Buchala, A.; Métraux, J.-P.; Broekaert, W.F. Concomitant Activation of Jasmonate and Ethylene Response Pathways Is Required for Induction of a Plant Defensin Gene in Arabidopsis. Plant Cell 1998, 10, 2103–2113. [Google Scholar] [CrossRef]
- Servin, A.; Elmer, W.; Mukherjee, A.; De la Torre-Roche, R.; Hamdi, H.; White, J.C.; Bindraban, P.; Dimkpa, C. A Review of the Use of Engineered Nanomaterials to Suppress Plant Disease and Enhance Crop Yield. J. Nanopart. Res. 2015, 17, 92. [Google Scholar] [CrossRef]
- Poschenrieder, C.; Tolrà, R.; Barceló, J. Can Metals Defend Plants against Biotic Stress? Trends Plant Sci. 2006, 11, 288–295. [Google Scholar] [CrossRef]
- Römheld, V.; Marschner, H. Function of Micronutrients in Plants. In Micronutrients in Agriculture; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1991; pp. 297–328. ISBN 978-0-89118-878-0. [Google Scholar]
- Brion, D.; Datnoff, L.E.; Elmer, W.H.; Huber, D.M. Zinc and Plant Diseases. In Mineral Nutrition and Plant Disease; APS Press: St. Paul, MN, USA, 2007; pp. 157–179. [Google Scholar]
- Huber, D.M. Nitrogen and Plant Disease. Miner. Nutr. Plant Dis. 2007, 31–44. [Google Scholar]
- Selph, A.; Dutta, B.; Burrell, D.; Gitaitis, R. Effect of Crop Rotation and of Soil and Plant Tissue Micronutrients on Sour Skin Incidence in Vidalia Onion. In Proceedings of the Phytopathology; APS Press: St. Paul, MN, USA, 2014; Volume 104, p. 9. [Google Scholar]
- Watson-Selph, A.; Gitaitis, R.; Dutta, B. Micronutrients May Affect Plant Disease Severity by Interacting with Components of the Systemic Acquired Resistance Pathway. In Proceedings of the Phytopathology; APS Press: St. Paul, MN, USA, 2016; Volume 106, p. 15. [Google Scholar]
- Gitaitis, R.; Nischwitz, C.; Mullis, S.W.; Csinos, A.; Burrell, D.; Pearce, M. Soil Fertility Related to Tomato Spotted Wilt Virus in Tobacco. In Proceedings of the Phytopathology; APS Press: St. Paul, MN, USA, 2009; Volume 99, p. 198. [Google Scholar]
- Dutta, B.; Langston, D.B.; Luo, X.; Carlson, S.; Kichler, J.; Gitaitis, R. A Risk Assessment Model for Bacterial Leaf Spot of Pepper (Capsicum annuum), Caused by Xanthomonas euvesicatoria, Based on Concentrations of Macronutrients, Micronutrients, and Micronutrient Ratios. Phytopathology 2017, 107, 1331–1338. [Google Scholar] [CrossRef] [PubMed]
- Rooks, S.A.; Watson-Selph, A.K.; Dutta, B.; Gitaitis, R.D.; Nischwitz, C.; Mullis, S.W.; Culbreath, A.K.; Csinos, A.S. Role of Minor Nutrients on Tomato Spotted Wilt of Tobacco (Nicotania Tabacum) and Superoxide-Dismutase and NPR1 Genes. In Proceedings of the Phytopathology; APS Press: St. Paul, MN, USA, 2015; Volume 105. [Google Scholar]
- Segond, D.; Dellagi, A.; Lanquar, V.; Rigault, M.; Patrit, O.; Thomine, S.; Expert, D. NRAMP Genes Function in Arabidopsis thaliana Resistance to Erwinia chrysanthemi Infection. Plant J. 2009, 58, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chien, W.-F.; Lin, N.-C.; Yeh, K.-C. Alternative Functions of Arabidopsis YELLOW STRIPE-LIKE3: From Metal Translocation to Pathogen Defense. PLoS ONE 2014, 9, e98008. [Google Scholar] [CrossRef]
- Liu, B.; Wasilwa, L.A.; Morelock, T.E.; O’Neill, N.R.; Correll, J.C. Comparison of Colletotrichum orbiculare and Several Allied Colletotrichum spp. for MtDNA RFLPs, Intron RFLP and Sequence Variation, Vegetative Compatibility, and Host Specificity. Phytopathology 2007, 97, 1305–1314. [Google Scholar]
- Ye, F.; Albarouki, E.; Lingam, B.; Deising, H.B.; von Wirén, N. An Adequate Fe Nutritional Status of Maize Suppresses Infection and Biotrophic Growth of Colletotrichum Graminicola. Physiol. Plant. 2014, 151, 280–292. [Google Scholar] [CrossRef]
- Fernando, D.R.; Baker, A.J.M.; Woodrow, I.E.; Fernando, D.R.; Baker, A.J.M.; Woodrow, I.E. Physiological Responses in Macadamia Integrifolia on Exposure to Manganese Treatment. Aust. J. Bot. 2009, 57, 406–413. [Google Scholar] [CrossRef]
- Millaleo, R.; Reyes-Diaz, M.; Ivanov, A.G.; Mora, M.L.; Alberdi, M. Manganese as Essential and Toxic Element for Plants: Transport, Accumulation and Resistance Mechanisms. J. Soil Sci. Plant Nutr. 2010, 10, 470–481. [Google Scholar] [CrossRef] [Green Version]
- Thompson, I.A.; Huber, D.M. Manganese and Plant Disease. Miner. Nutr. Plant Dis. 2007, 139, 153. [Google Scholar]
- Grewal, H.S.; Graham, R.D.; Rengel, Z. Genotypic Variation in Zinc Efficiency and Resistance to Crown Rot Disease (Fusarium graminearum Schw. Group 1) in Wheat. Plant Soil 1996, 186, 219–226. [Google Scholar] [CrossRef]
- Li, Z.; Fan, Y.; Gao, L.; Cao, X.; Ye, J.; Li, G. The Dual Roles of Zinc Sulfate in Mitigating Peach Gummosis. Plant Dis. 2016, 100, 345–351. [Google Scholar] [CrossRef]
- Steiner, A.A. The Universal Nutrient Solution. In Proceedings of the 6th International Congress on Soilless Culture, Lunteren, The Netherlands, 29 April–5 May 1984. [Google Scholar]
- Mandal, M.K.; Suren, H.; Ward, B.; Boroujerdi, A.; Kousik, C. Differential Roles of Melatonin in Plant-Host Resistance and Pathogen Suppression in Cucurbits. J. Pineal Res. 2018, 65, e12505. [Google Scholar] [CrossRef]
- Kong, Q.; Yuan, J.; Gao, L.; Zhao, S.; Jiang, W.; Huang, Y.; Bie, Z. Identification of Suitable Reference Genes for Gene Expression Normalization in QRT-PCR Analysis in Watermelon. PLoS ONE 2014, 9, e90612. [Google Scholar] [CrossRef] [PubMed]
- Mathys, J.; De Cremer, K.; Timmermans, P.; Van Kerkhove, S.; Lievens, B.; Vanhaecke, M.; Cammue, B.; De Coninck, B. Genome-Wide Characterization of ISR Induced in Arabidopsis thaliana by Trichoderma Hamatum T382 against Botrytis Cinerea Infection. Front. Plant Sci. 2012, 3, 108. [Google Scholar] [CrossRef] [PubMed]
- Tamaoki, D.; Seo, S.; Yamada, S.; Kano, A.; Miyamoto, A.; Shishido, H.; Miyoshi, S.; Taniguchi, S.; Akimitsu, K.; Gomi, K. Jasmonic Acid and Salicylic Acid Activate a Common Defense System in Rice. Plant Signal. Behav. 2013, 8, e24260. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Vaishnav, A.; Varma, A.; Choudhary, D.K. Comparative Expression Analysis of Defence-Related Genes in Bacillus-Treated Glycine Max upon Challenge Inoculation with Selective Fungal Phytopathogens. Curr. Sci. 2018, 115, 1950–1956. [Google Scholar] [CrossRef]
- Staswick, P.E. Storage Proteins of Vegetative Plant Tissues. Annu. Rev. Plant Biol. 1994, 45, 303–322. [Google Scholar] [CrossRef]
- Liu, Y.; Ahn, J.-E.; Datta, S.; Salzman, R.A.; Moon, J.; Huyghues-Despointes, B.; Pittendrigh, B.; Murdock, L.L.; Koiwa, H.; Zhu-Salzman, K. Arabidopsis Vegetative Storage Protein Is an Anti-Insect Acid Phosphatase. Plant Physiol. 2005, 139, 1545–1556. [Google Scholar] [CrossRef]
- De Oliveira Carvalho, A.; Moreira Gomes, V. Plant Defensins and Defensin-like Peptides—Biological Activities and Biotechnological Applications. Curr. Pharm. Des. 2011, 17, 4270–4293. [Google Scholar] [CrossRef] [PubMed]
- De Coninck, B.; Cammue, B.P.A.; Thevissen, K. Modes of Antifungal Action and in Planta Functions of Plant Defensins and Defensin-like Peptides. Fungal Biol. Rev. 2013, 26, 109–120. [Google Scholar] [CrossRef]
- Dos Santos, C.V.; Letousey, P.; Delavault, P.; Thalouarn, P. Defense Gene Expression Analysis of Arabidopsis thaliana Parasitized by Orobanche ramosa. Phytopathology 2003, 93, 451–457. [Google Scholar] [CrossRef]
- Vijayan, P.; Shockey, J.; Lévesque, C.A.; Cook, R.J.; Browse, J. A Role for Jasmonate in Pathogen Defense of Arabidopsis. Proc. Natl. Acad. Sci. USA 1998, 95, 7209–7214. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, J. The Essential Role of Jasmonic Acid in Plant–Herbivore Interactions—Using the Wild Tobacco Nicotiana Attenuata as a Model. J. Genet. Genom. 2013, 40, 597–606. [Google Scholar] [CrossRef]
- Upchurch, R.G.; Ramirez, M.E. Defense-Related Gene Expression in Soybean Leaves and Seeds Inoculated with Cercospora kikuchii and Diaporthe phaseolorum Var. meridionalis. Physiol. Mol. Plant Pathol. 2010, 75, 64–70. [Google Scholar] [CrossRef]
- Lv, H.; Cao, H.; Nawaz, M.A.; Sohail, H.; Huang, Y.; Cheng, F.; Kong, Q.; Bie, Z. Wheat Intercropping Enhances the Resistance of Watermelon to Fusarium Wilt. Front. Plant Sci. 2018, 9, 696. [Google Scholar] [CrossRef]
- Schaible, U.E.; Kaufmann, S.H.E. Iron and Microbial Infection. Nat. Rev. Microbiol. 2004, 2, 946–953. [Google Scholar] [CrossRef]
- Machado, P.P.; Steiner, F.; Zuffo, A.M.; Machado, R.A. Could the Supply of Boron and Zinc Improve Resistance of Potato to Early Blight? Potato Res. 2018, 61, 169–182. [Google Scholar] [CrossRef]
- Helfenstein, J.; Pawlowski, M.L.; Hill, C.B.; Stewart, J.; Lagos-Kutz, D.; Bowen, C.R.; Frossard, E.; Hartman, G.L. Zinc Deficiency Alters Soybean Susceptibility to Pathogens and Pests. J. Plant Nutr. Soil Sci. 2015, 178, 896–903. [Google Scholar] [CrossRef]
- Petkar, A.; Langston, D.B.; Buck, J.W.; Stevenson, K.L.; Ji, P. Sensitivity of Fusarium oxysporum f. sp. niveum to Prothioconazole and Thiophanate-Methyl and Gene Mutation Conferring Resistance to Thiophanate-Methyl. Plant Dis. 2017, 101, 366–371. [Google Scholar] [CrossRef] [Green Version]
- Turechek, W.W.; Stevenson, K.L. Effects of Host Resistance, Temperature, Leaf Wetness, and Leaf Age on Infection and Lesion Development of Pecan Scab. Phytopathology 1998, 88, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Freckman, D.W.; Kaplan, D.T.; Van Gundy, S.D. A Comparison of Techniques for Extraction and Study of Anhydrobiotic Nematodes from Dry Soils. J. Nematol. 1977, 9, 176–181. [Google Scholar]
- Djami-Tchatchou, A.T.; Straker, C.J. The Isolation of High Quality RNA from the Fruit of Avocado (Persea americana Mill.). S. Afr. J. Bot. 2012, 78, 44–46. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CT Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Nash, S.M.; Snyder, W.C. Quantitative Estimations by Plate Counts of Propagules of the Bean Root Rot Fusarium in Field Soils. Phytopathology 1962, 52, 567–572. [Google Scholar]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Oxford, UK, 2006. [Google Scholar]
- Lin, Y.-H.; Chen, K.-S.; Chang, J.-Y.; Wan, Y.-L.; Hsu, C.-C.; Huang, J.-W.; Chang, P.-F.L. Development of the Molecular Methods for Rapid Detection and Differentiation of Fusarium oxysporum and F. oxysporum f. sp. niveum in Taiwan. New Biotechnol. 2010, 27, 409–418. [Google Scholar] [CrossRef]
- Taylor, A.L.; Sasser, J.N. Biology, Identification and Control of Some Root-Knot Nematodes (Meloidogyne Spp.); Department of Plant Pathology, North Carolina State University, United States Agency for International Development: Raleigh, NC, USA, 1978; p. 111. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Micronutrient Treatments | N | P | K | Ca | Mg | B | Cu | Mo | Fe | Mn | Zn |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Steiner a | 256 | 48 | 304 | 180 | 48 | 1 | 0.2 | 0.1 | 3 | 1 | 0.4 |
| High Fe | 256 | 48 | 304 | 180 | 48 | 1 | 0.2 | 0.1 | 9 | 1 | 0.4 |
| Low Fe | 256 | 48 | 304 | 180 | 48 | 1 | 0.2 | 0.1 | 1.5 | 1 | 0.4 |
| High Mn | 256 | 48 | 304 | 180 | 48 | 1 | 0.2 | 0.1 | 3 | 3 | 0.4 |
| Low Mn | 256 | 48 | 304 | 180 | 48 | 1 | 0.2 | 0.1 | 3 | 0.5 | 0.4 |
| High Zn | 256 | 48 | 304 | 180 | 48 | 1 | 0.2 | 0.1 | 3 | 1 | 1.2 |
| Low Zn | 256 | 48 | 304 | 180 | 48 | 1 | 0.2 | 0.1 | 3 | 1 | 0.2 |
| Micronutrient | Micronutrient Treatment | Day 0 (ppm; Applied) | Day 7 (ppm) |
|---|---|---|---|
| Fe | High Fe | 9.0 a1 | 9.28 ± 0.29 a |
| Steiner | 3.0 b | 2.35 ± 0.33 b | |
| Low Fe | 1.5 c | 1.40 ± 0.05 c | |
| Mn | High Mn | 3.0 a | 2.25 ± 0.12 a |
| Steiner | 1.0 b | 0.57 ± 0.08 b | |
| Low Mn | 0.5 c | 0.29 ± 0.03 c | |
| Zn | High Zn | 1.2 a | 1.03 ± 0.05 a |
| Steiner | 0.4 b | 0.43 ± 0.08 b | |
| Low Zn | 0.2 c | 0.34 ± 0.07 b |
| Genes | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (3′-5′) | Comments | PCR Conditions |
|---|---|---|---|---|
| NPR1 | CGCTGCCGATATGCATGTGA | GTCAACCTTCAGCAAGTTGCCA | This study | 95 °C for 2 min; 35 cycles of 95 °C for 20 s, 62 °C for 30 s, and 72 °C for 60 s; and final extension of 72 °C for 6 min |
| PR1 | GACTCGCCTCAAGACTTTGT | GATGCGTTGGTTGGCATATTG | [66] | 95 °C for 3 min; 40 cycles of 95 °C for 10 s, 60 °C for 30 s, and 72 °C for 30 s; and final extension of 72 °C for 6 min |
| PR5 | CCTGGAGCGTCAAAGTCATTTA | CTCCAGTTAAGCAGGTGATACG | [66] | same as above |
| LOX | TCTCAACTGTGCTCCCATTC | GGAAGCAGTGGCTTTGAATTAC | This study | same as above |
| GCGAAGGTGTGCGAGAA | CATGGCAAGCTCCATGTTTG | This study | same as above | |
| VSP | ACCAAGGGAAGTCAGCAATAC | CCGAAACTGACGTACCCAATAA | This study | same as above |
| ß-Actin | CCATGTATGTGCCATCCAG | GGATAGCATGGGGTAGAGCA | [67] | [67] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karki, K.; Negi, V.S.; Coolong, T.; Petkar, A.; Mandal, M.; Kousik, C.; Gitaitis, R.; Hajihassani, A.; Dutta, B. Micronutrients Affect Expression of Induced Resistance Genes in Hydroponically Grown Watermelon against Fusarium oxysporum f. sp. niveum and Meloidogyne incognita. Pathogens 2022, 11, 1136. https://doi.org/10.3390/pathogens11101136
Karki K, Negi VS, Coolong T, Petkar A, Mandal M, Kousik C, Gitaitis R, Hajihassani A, Dutta B. Micronutrients Affect Expression of Induced Resistance Genes in Hydroponically Grown Watermelon against Fusarium oxysporum f. sp. niveum and Meloidogyne incognita. Pathogens. 2022; 11(10):1136. https://doi.org/10.3390/pathogens11101136
Chicago/Turabian StyleKarki, Kasmita, Vishal Singh Negi, Tim Coolong, Aparna Petkar, Mihir Mandal, Chandrasekar Kousik, Ron Gitaitis, Abolfazl Hajihassani, and Bhabesh Dutta. 2022. "Micronutrients Affect Expression of Induced Resistance Genes in Hydroponically Grown Watermelon against Fusarium oxysporum f. sp. niveum and Meloidogyne incognita" Pathogens 11, no. 10: 1136. https://doi.org/10.3390/pathogens11101136
APA StyleKarki, K., Negi, V. S., Coolong, T., Petkar, A., Mandal, M., Kousik, C., Gitaitis, R., Hajihassani, A., & Dutta, B. (2022). Micronutrients Affect Expression of Induced Resistance Genes in Hydroponically Grown Watermelon against Fusarium oxysporum f. sp. niveum and Meloidogyne incognita. Pathogens, 11(10), 1136. https://doi.org/10.3390/pathogens11101136