

The Bio-Diversity and the Role of Gut Microbiota in Postmenopausal Women with Luminal Breast Cancer Treated with Aromatase Inhibitors: An Observational Cohort Study

 ,
,  , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Materials and Methods
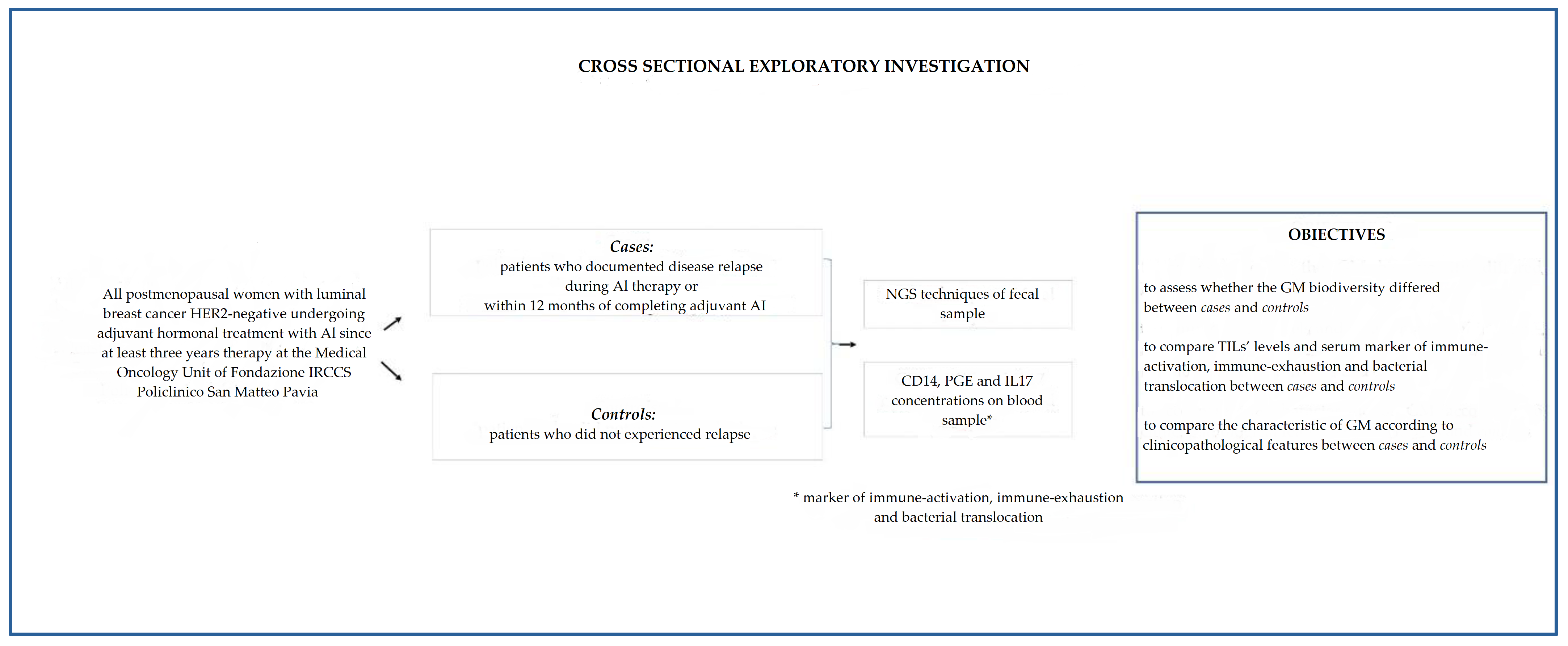
2.1. Study Setting
2.2. Outcomes
2.3. Biological Samples
2.3.1. Stool Sample Processing and DNA Extraction
2.3.2. Blood Sample Processing and DNA Extraction
2.4. TILs Levels
2.5. Statistical and Bioinformatic Analyses
3. Results
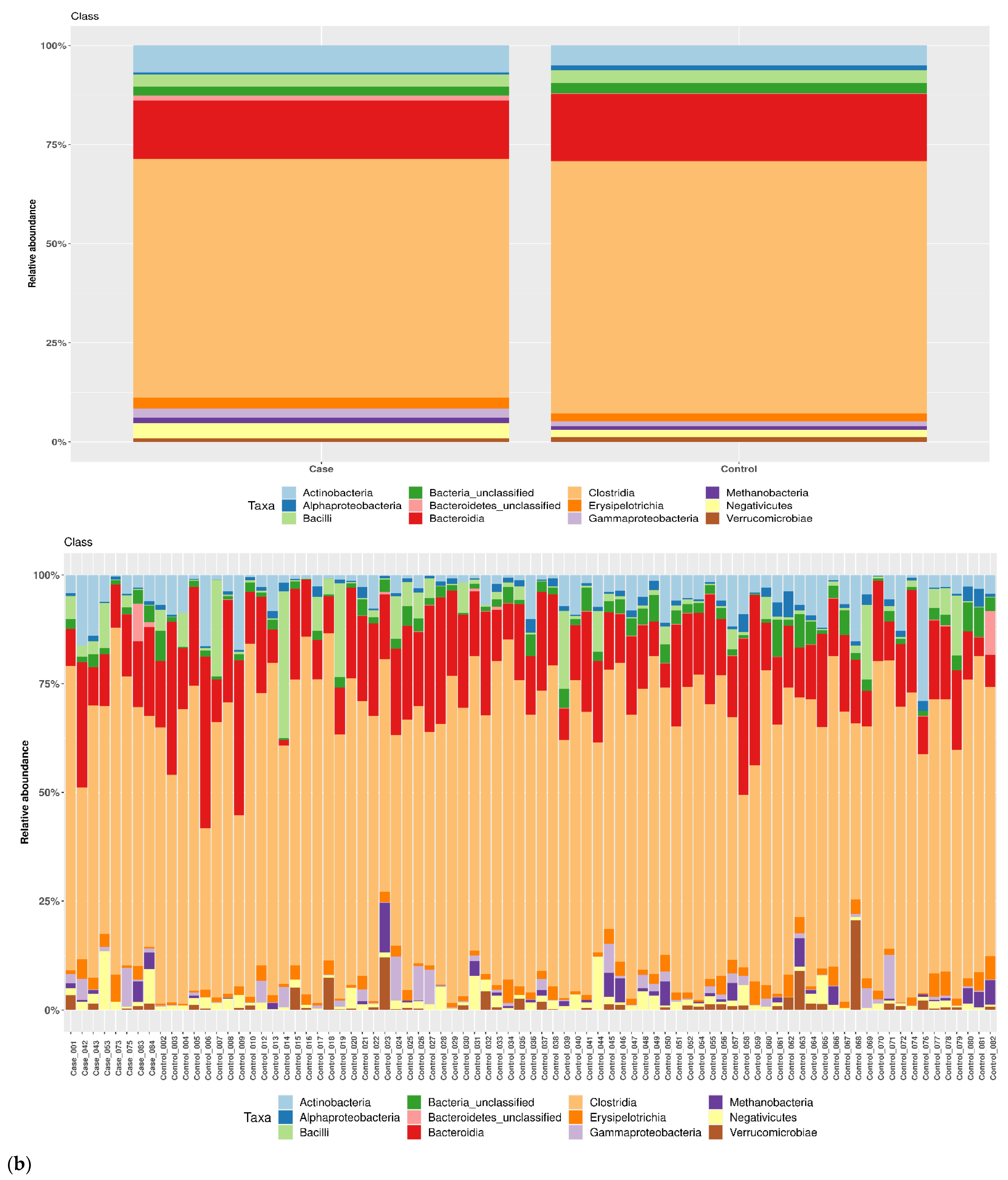
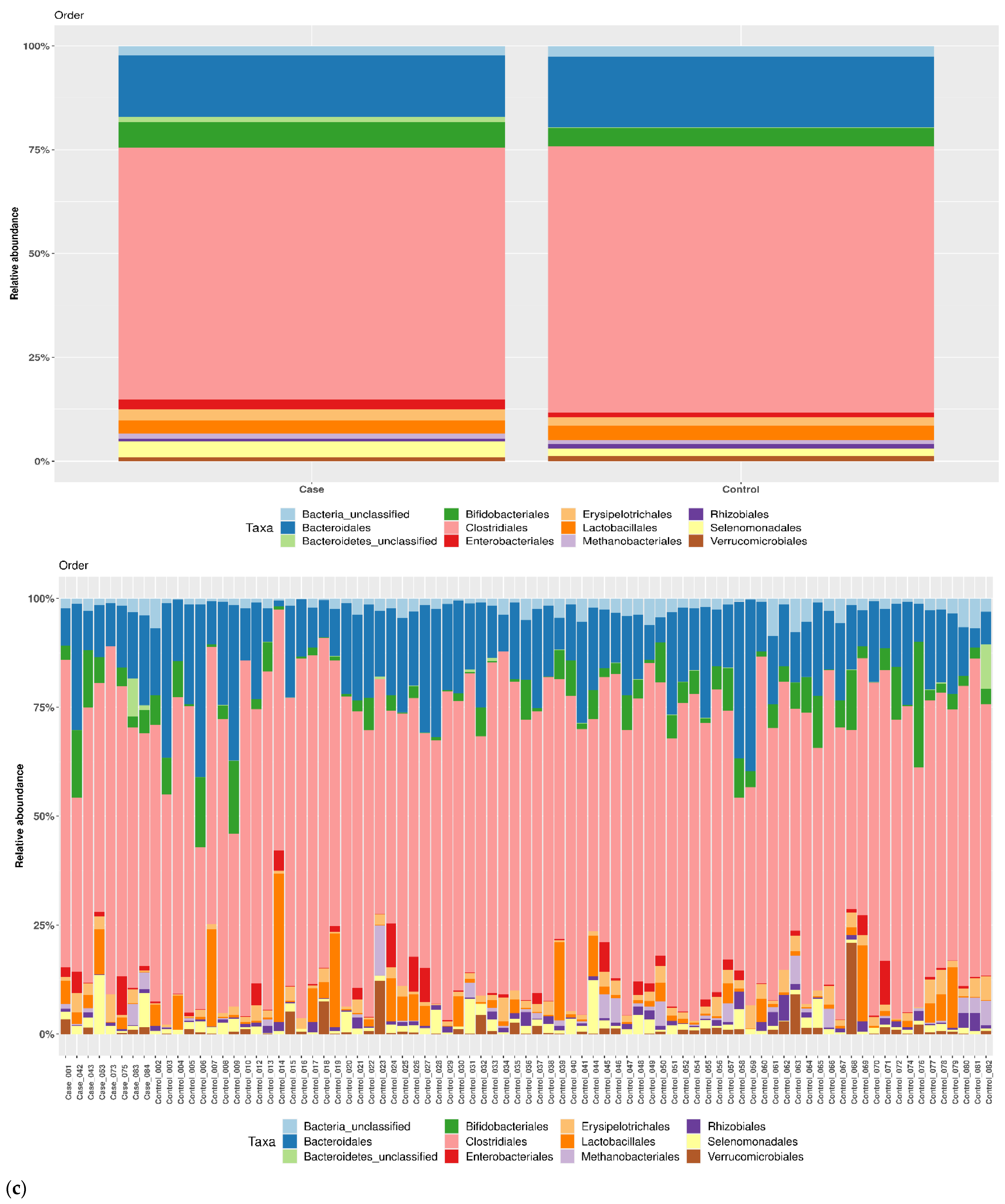
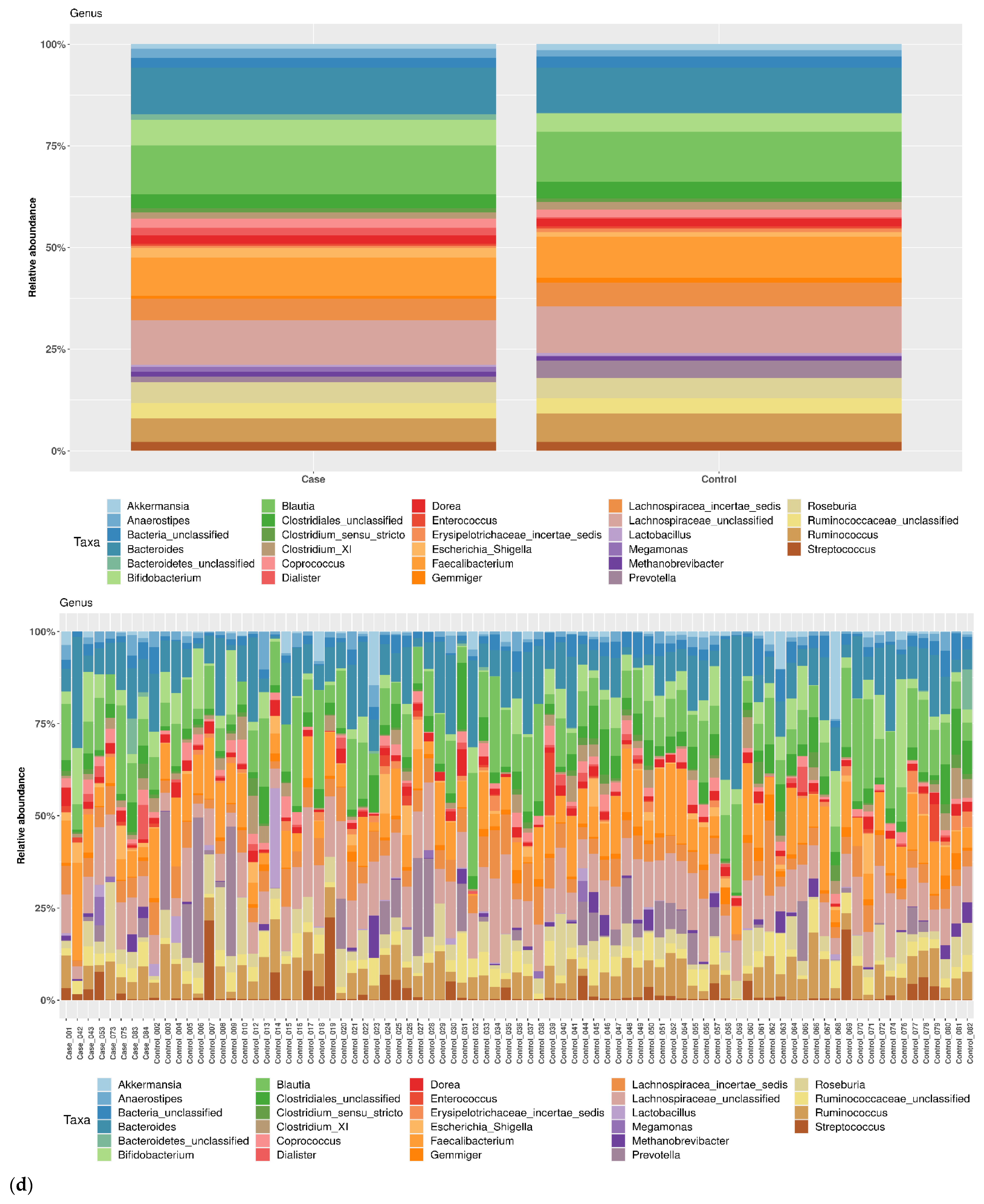
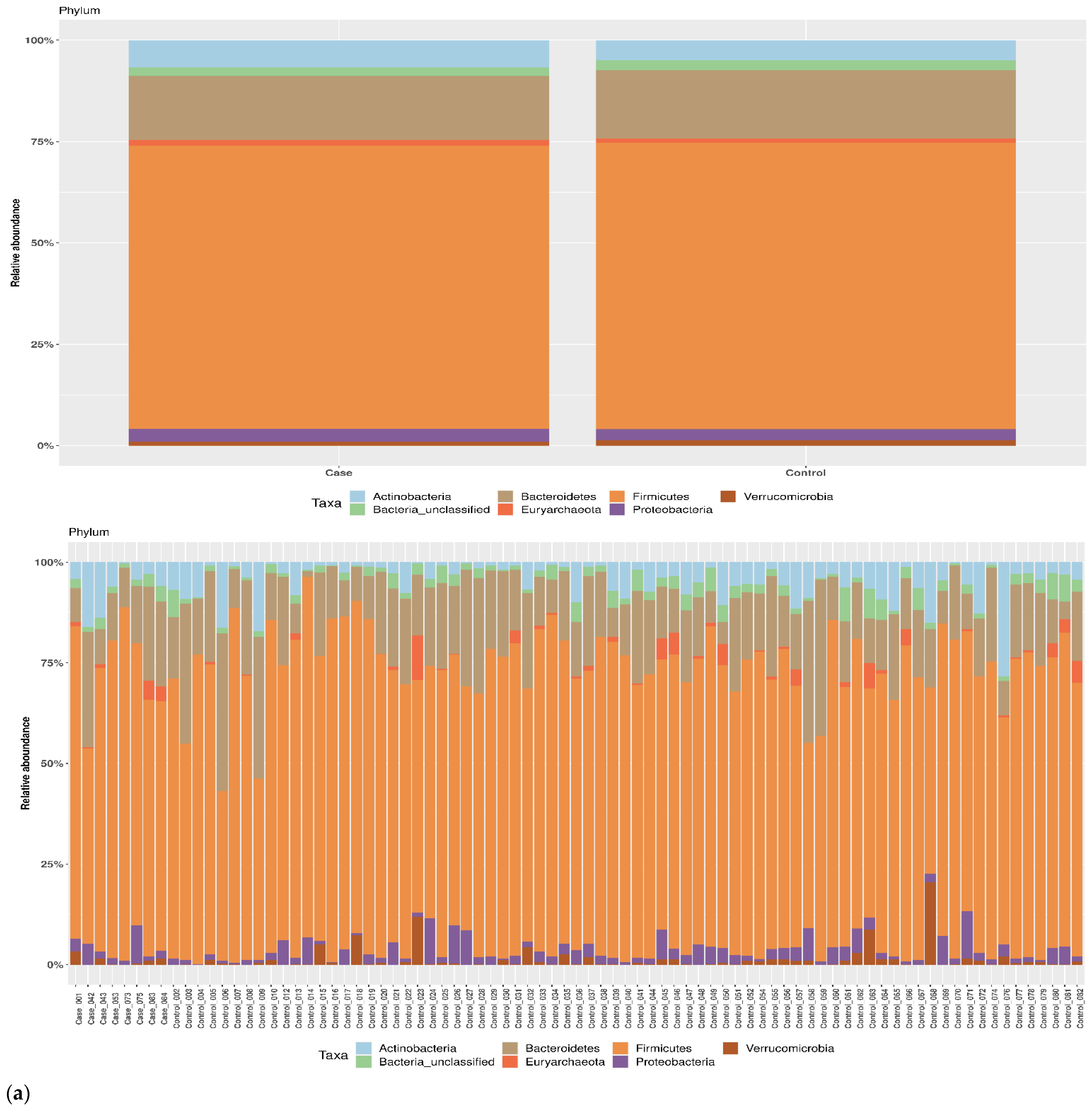
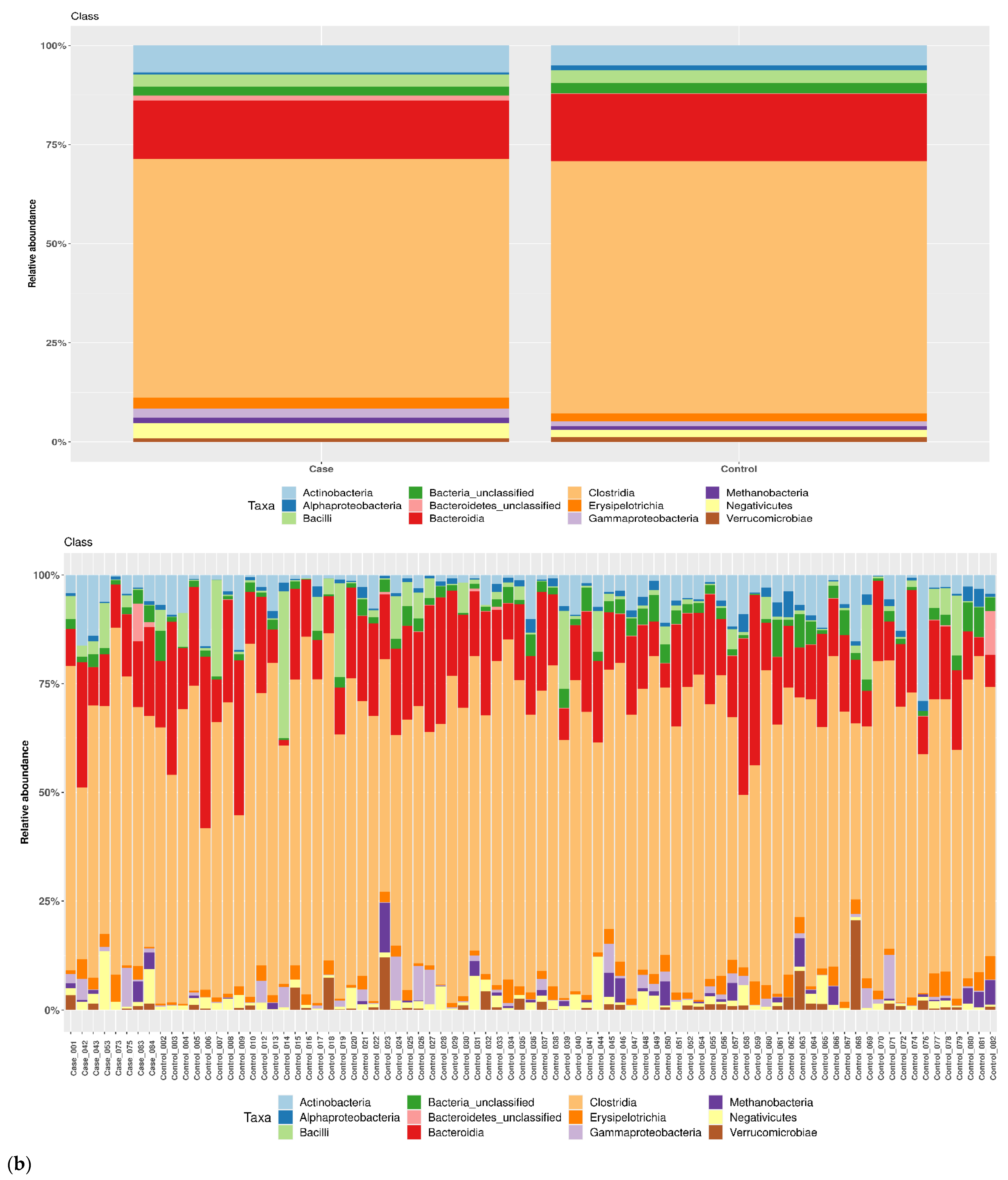
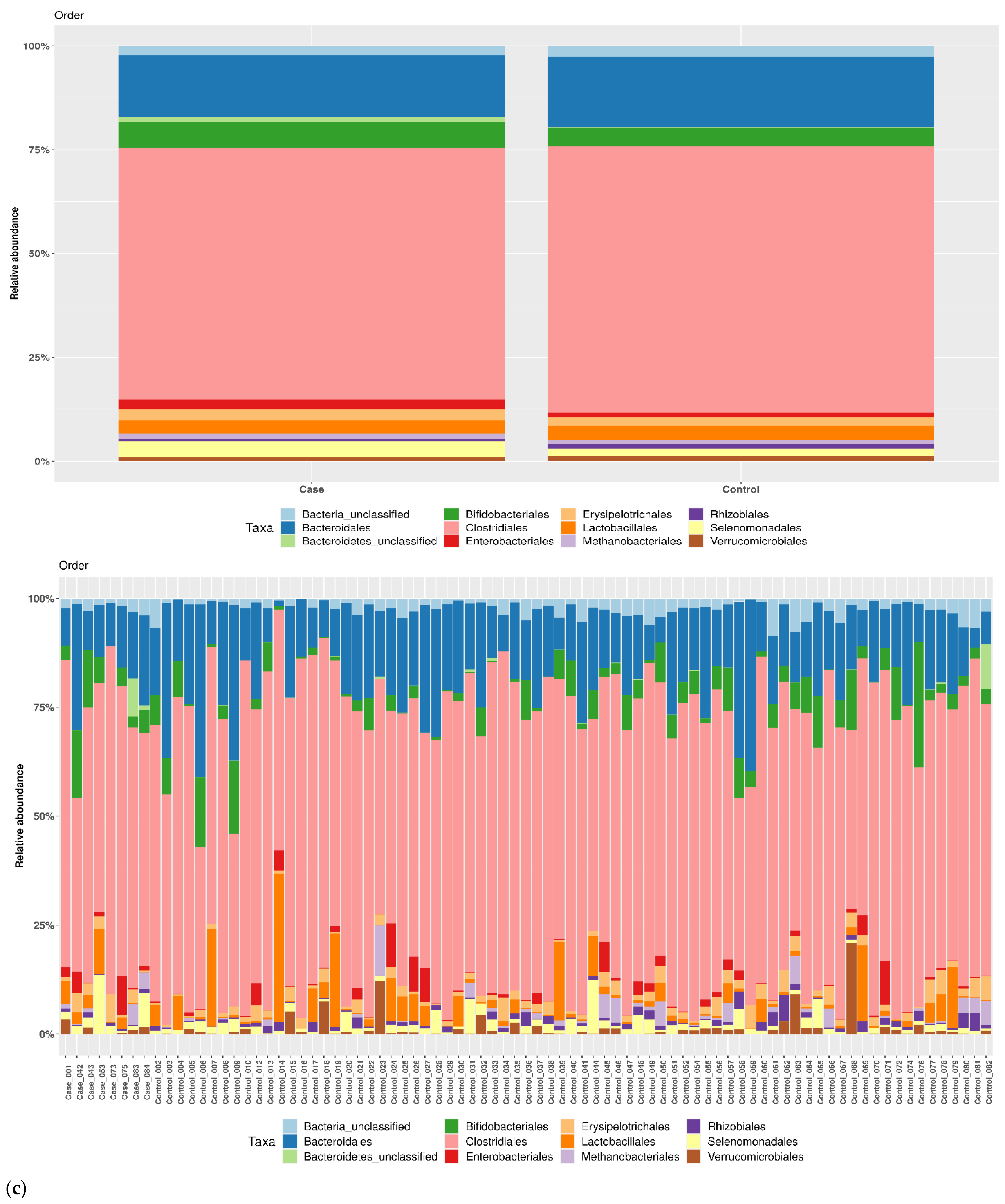
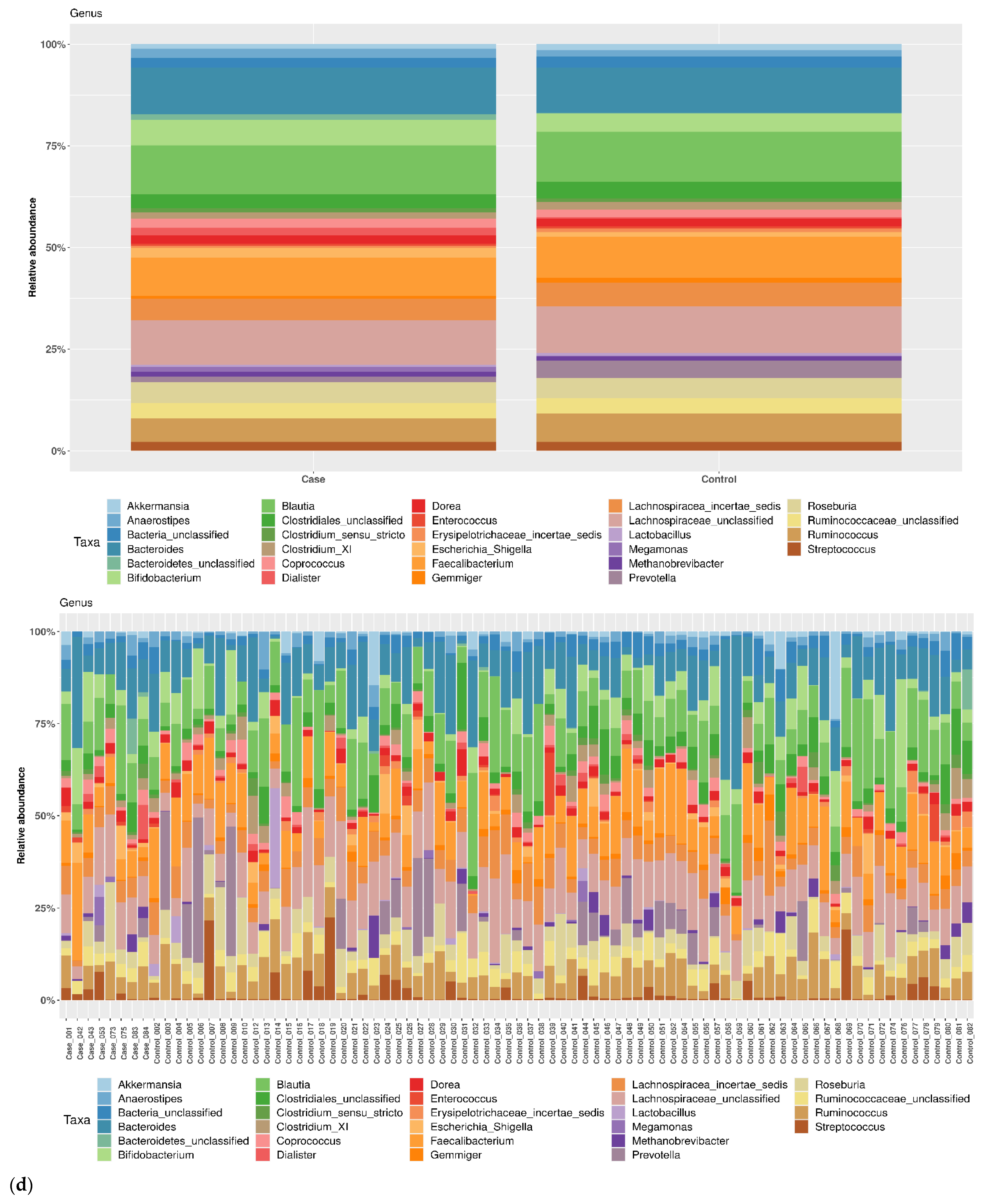
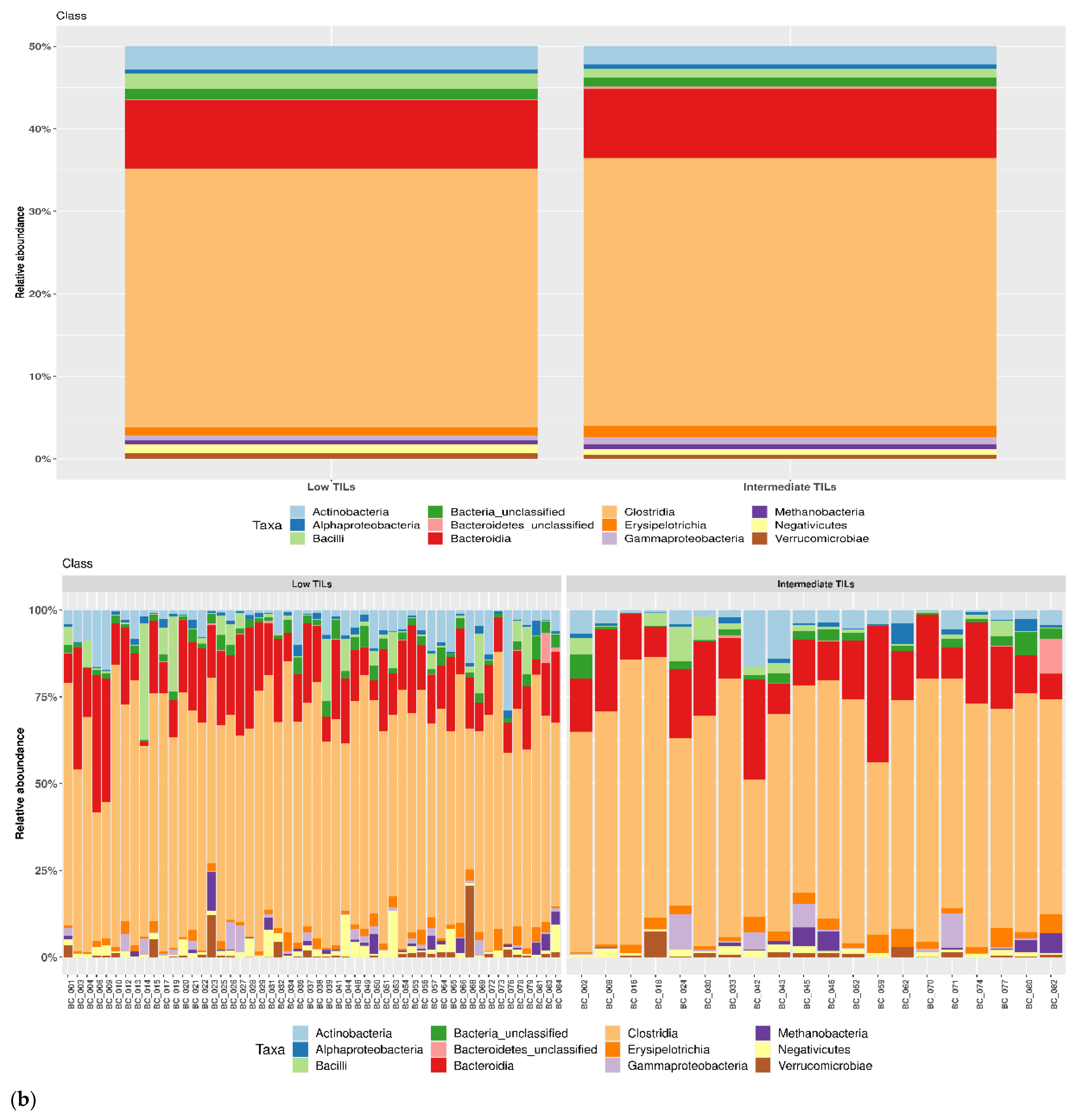
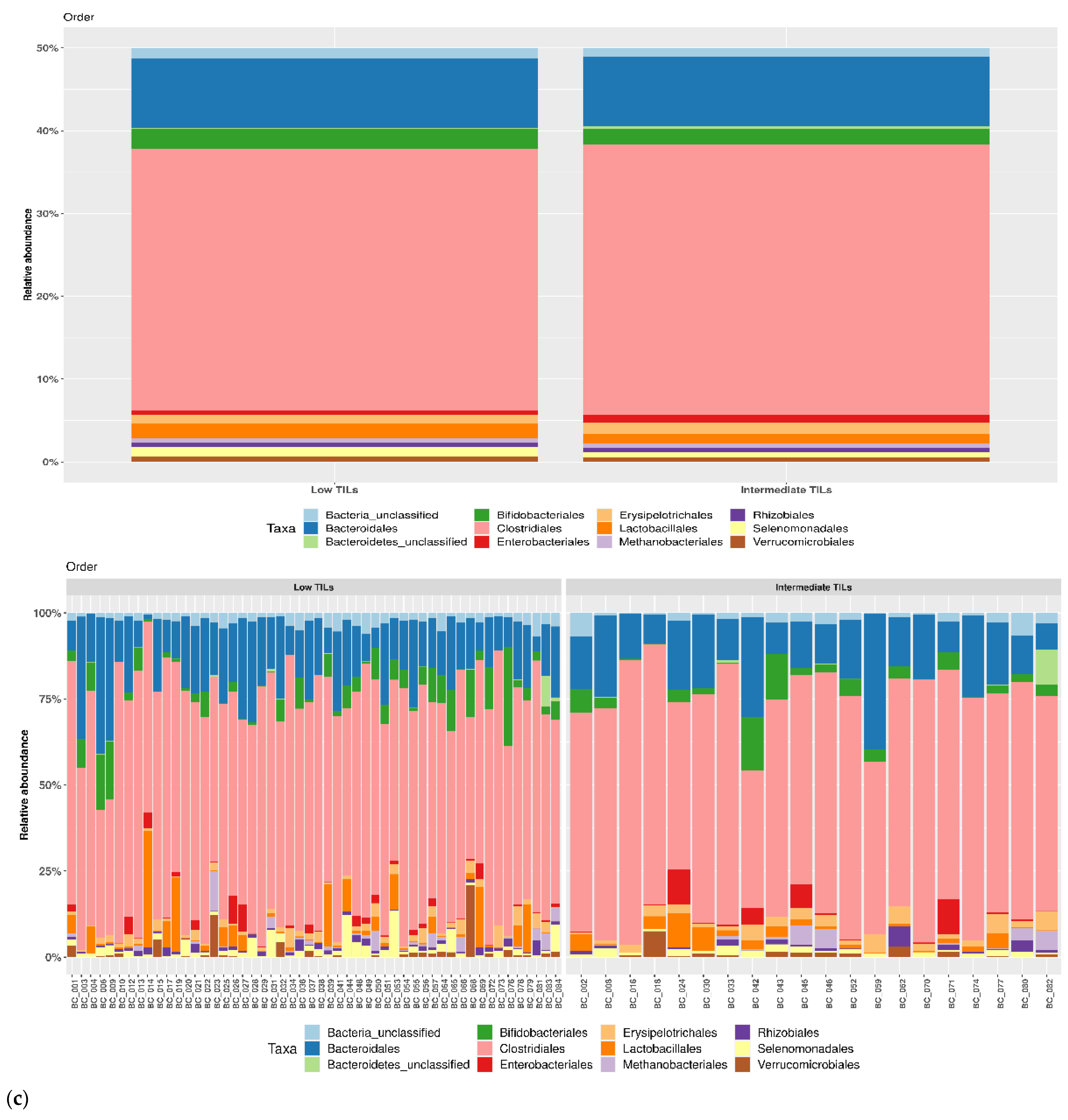
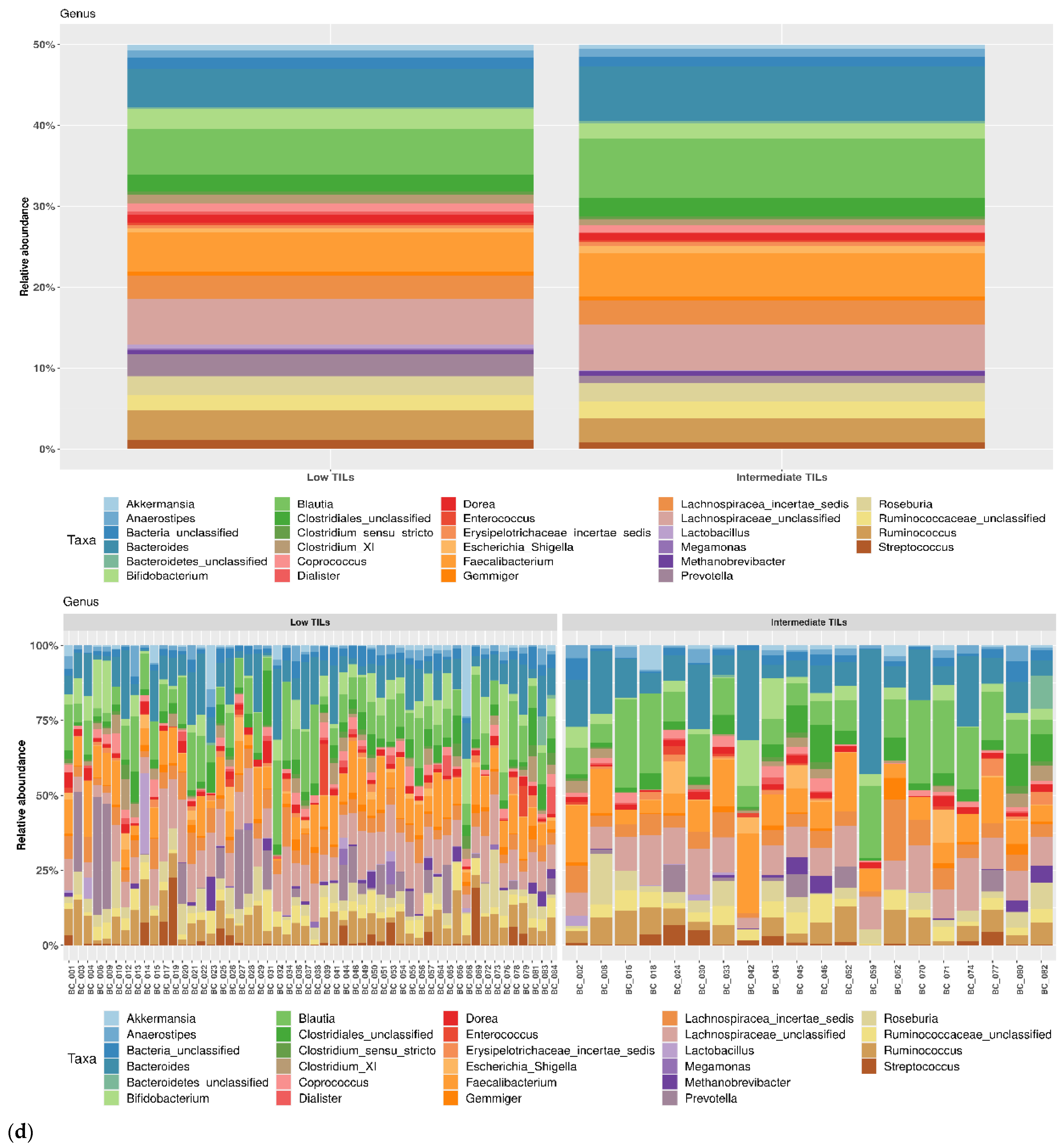
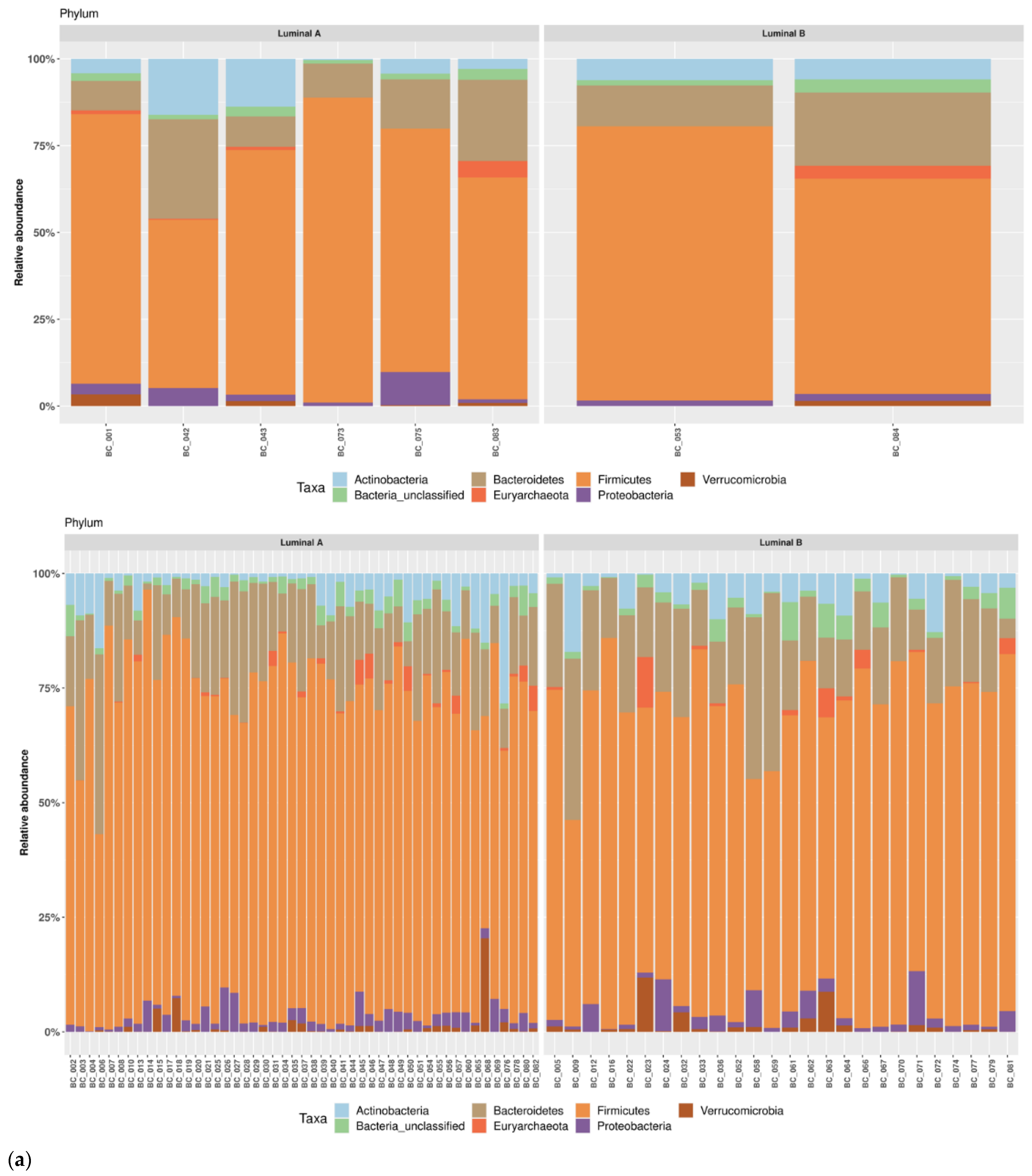
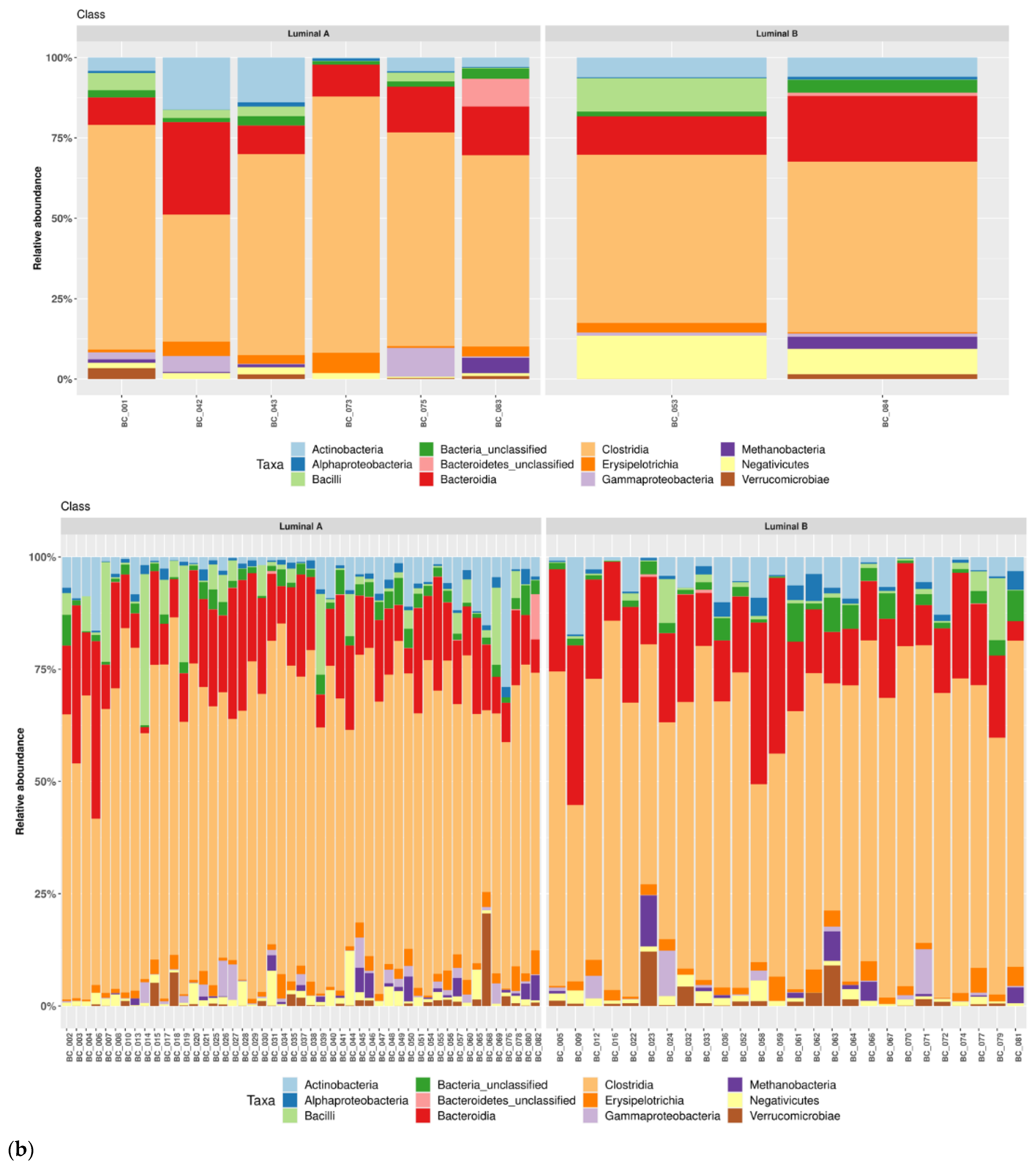
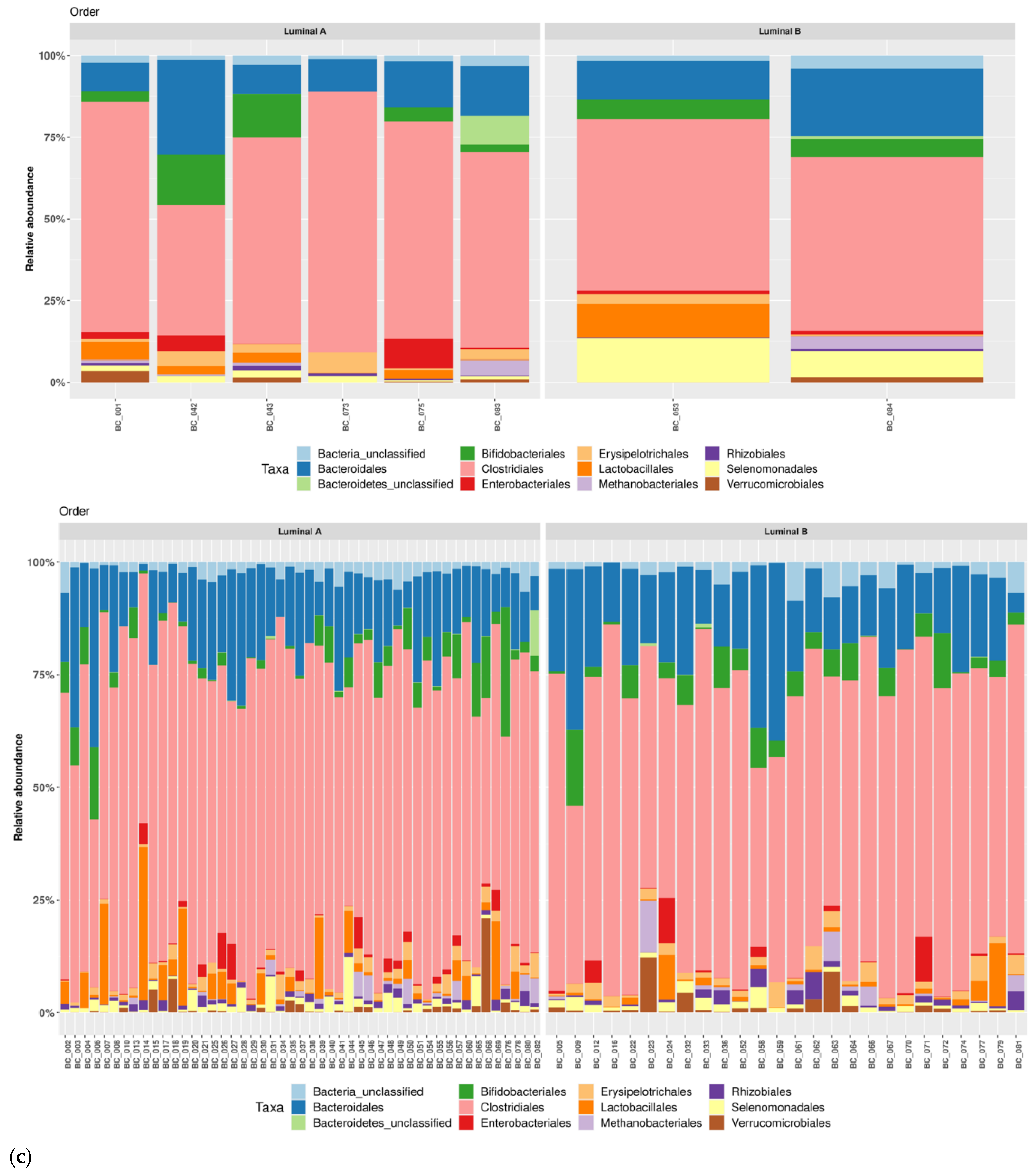
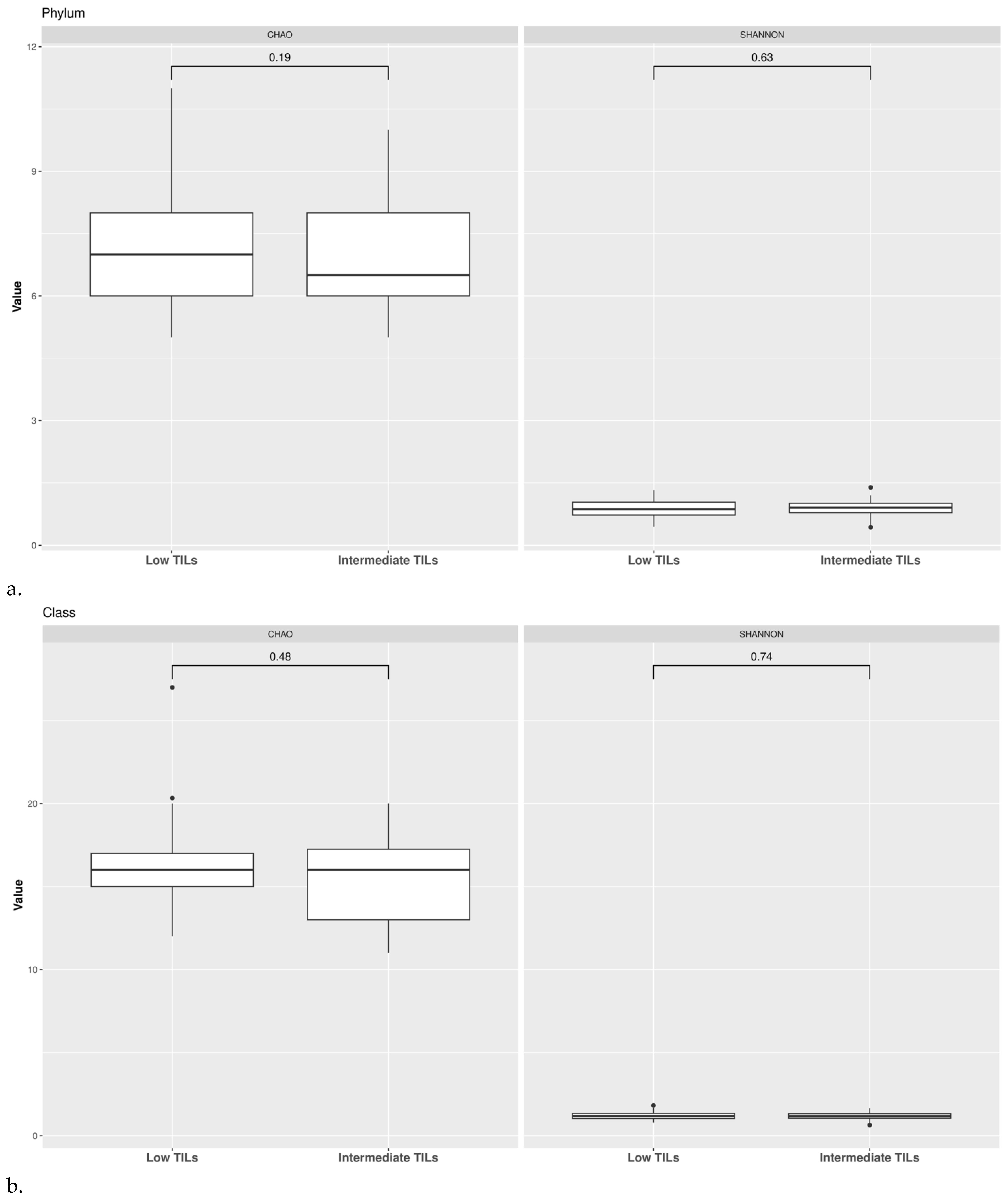
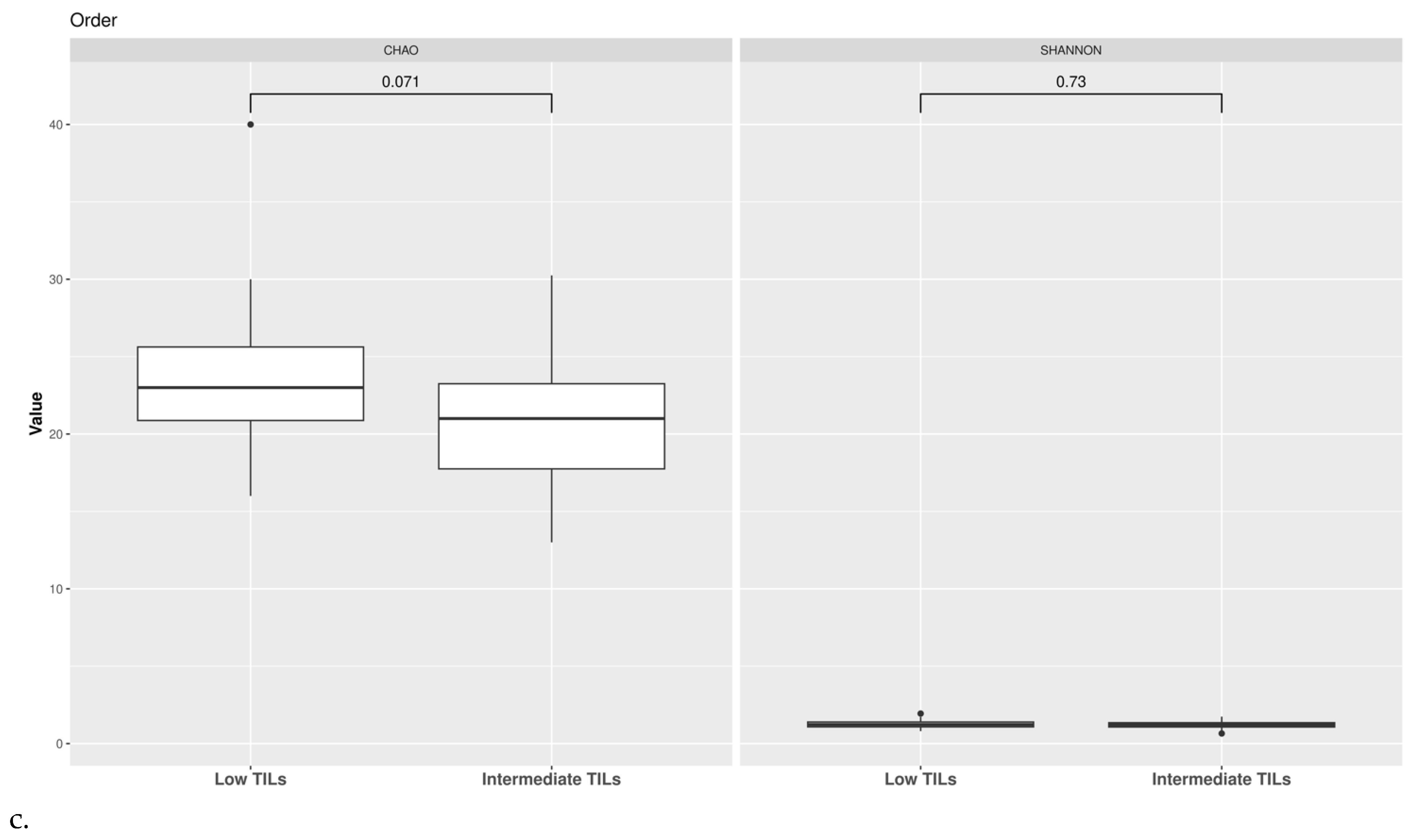
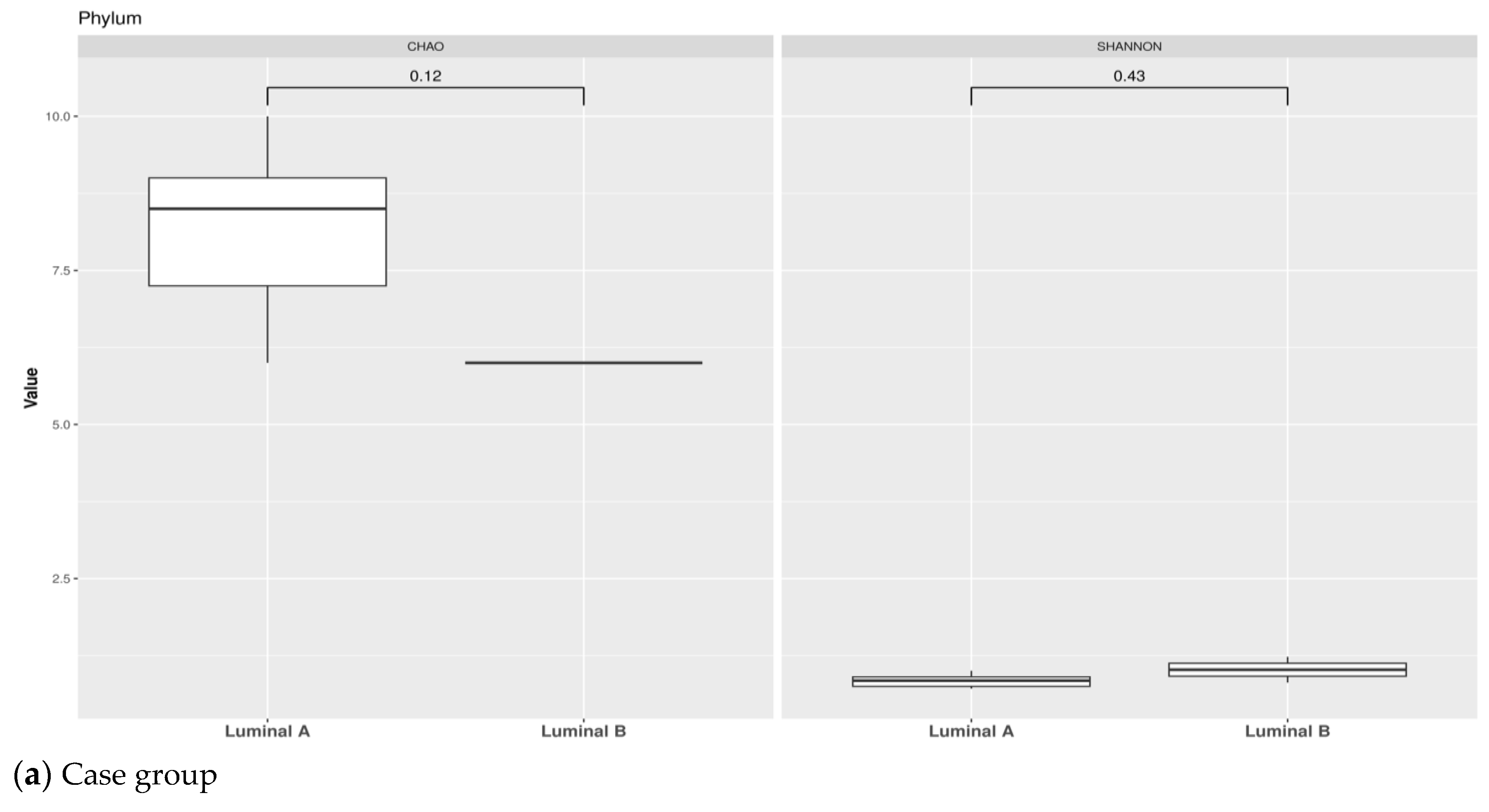
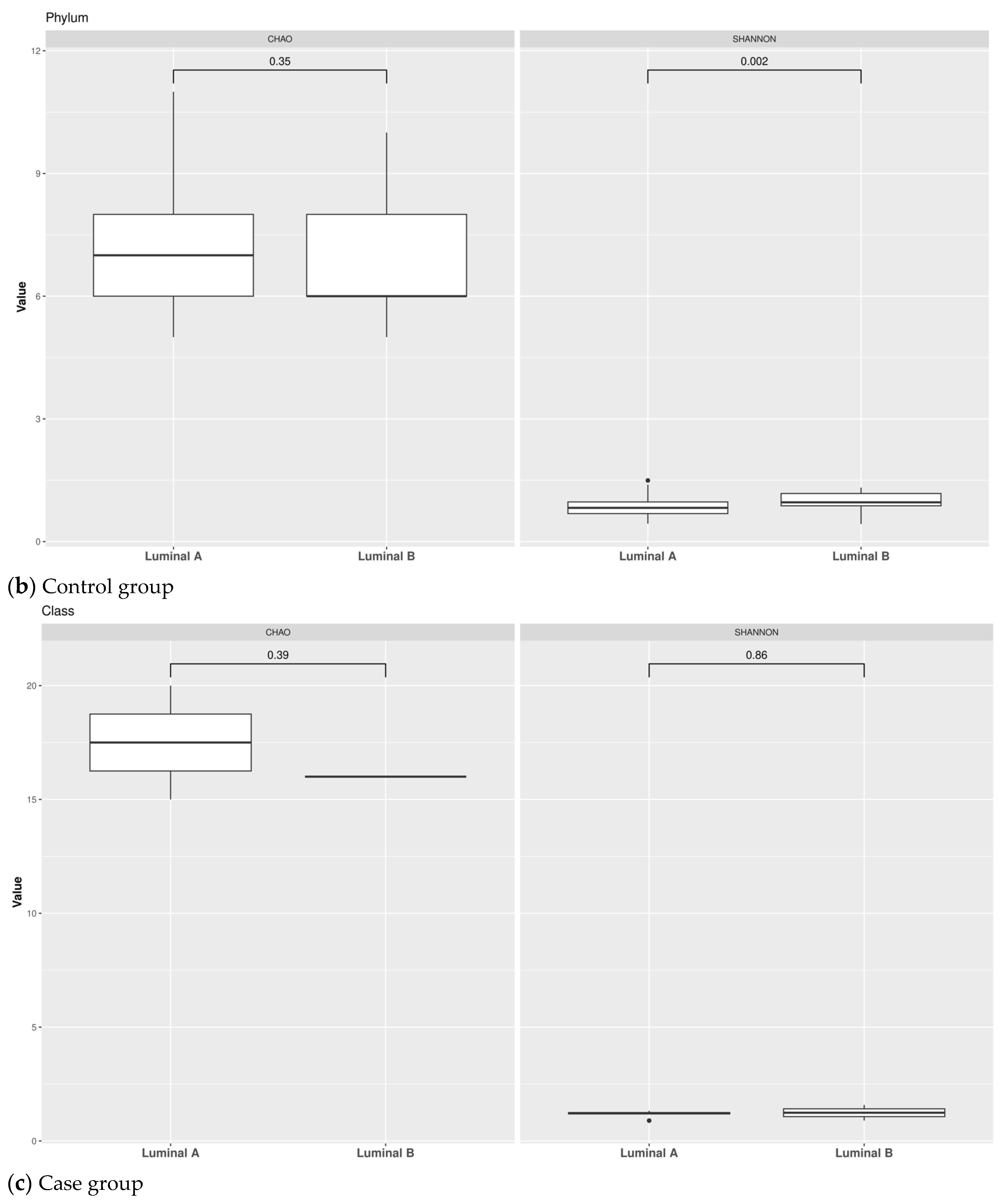
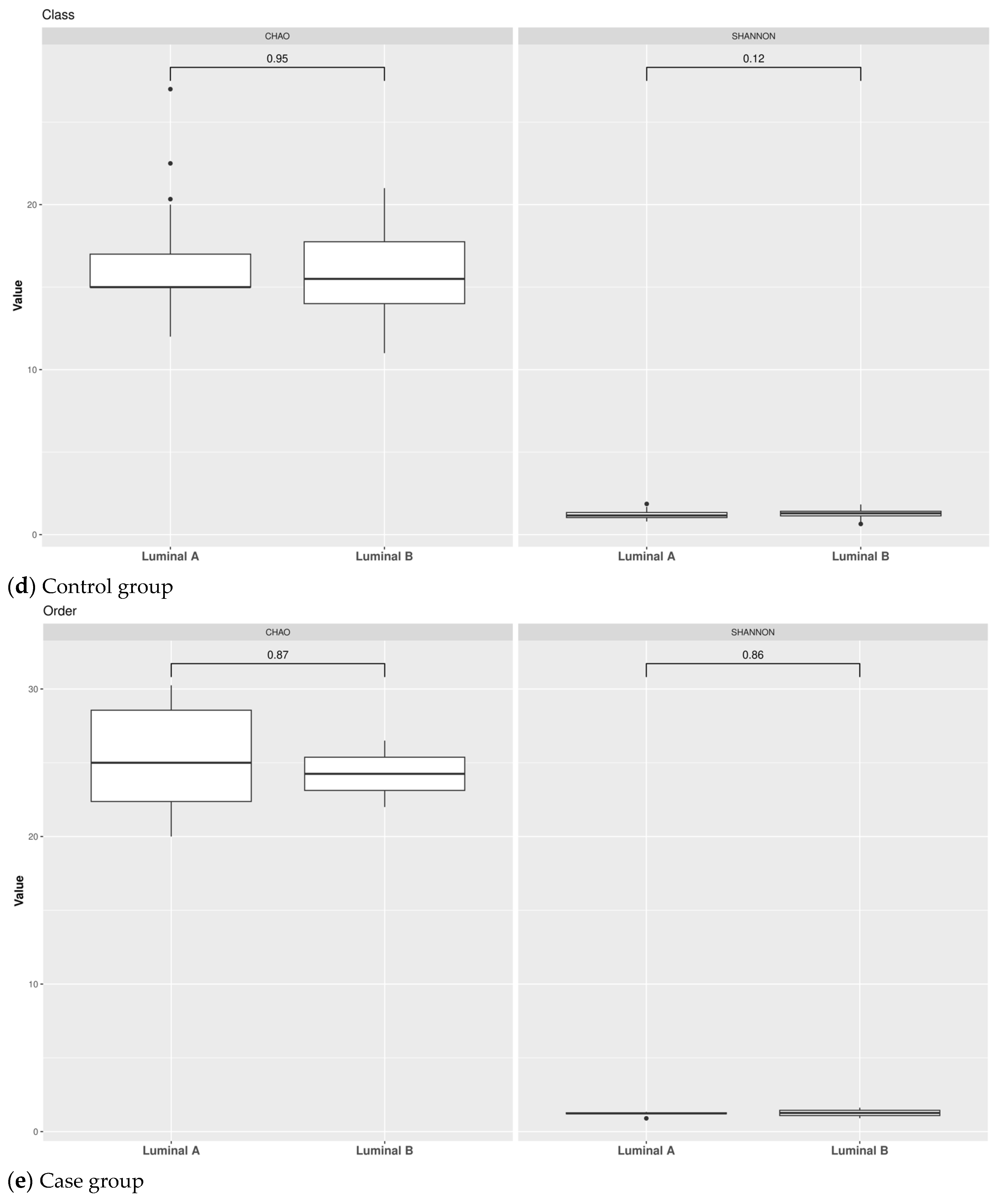
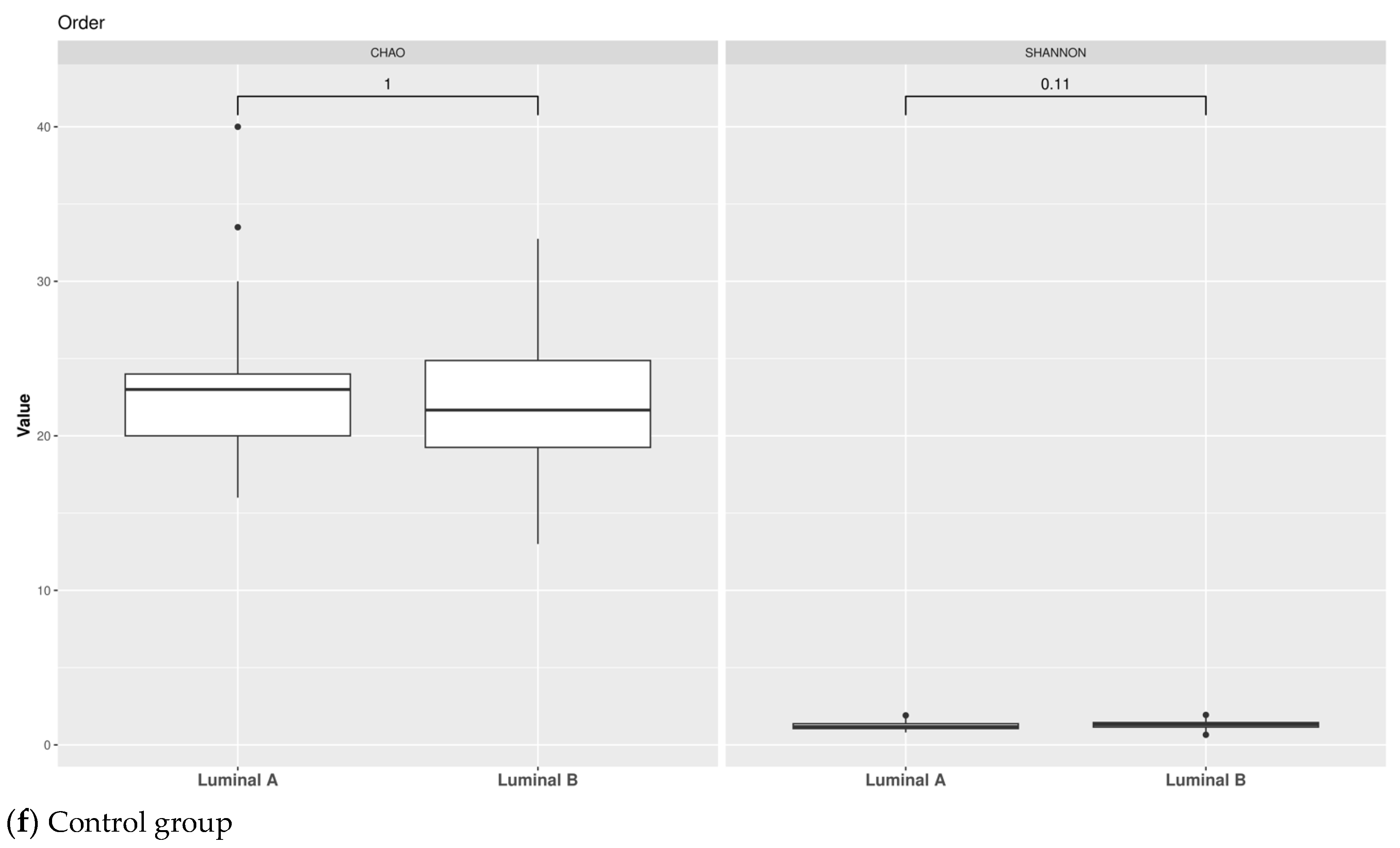
3.1. Taxonomic Structure of Fecal Bacterial Communities in Cases and Controls
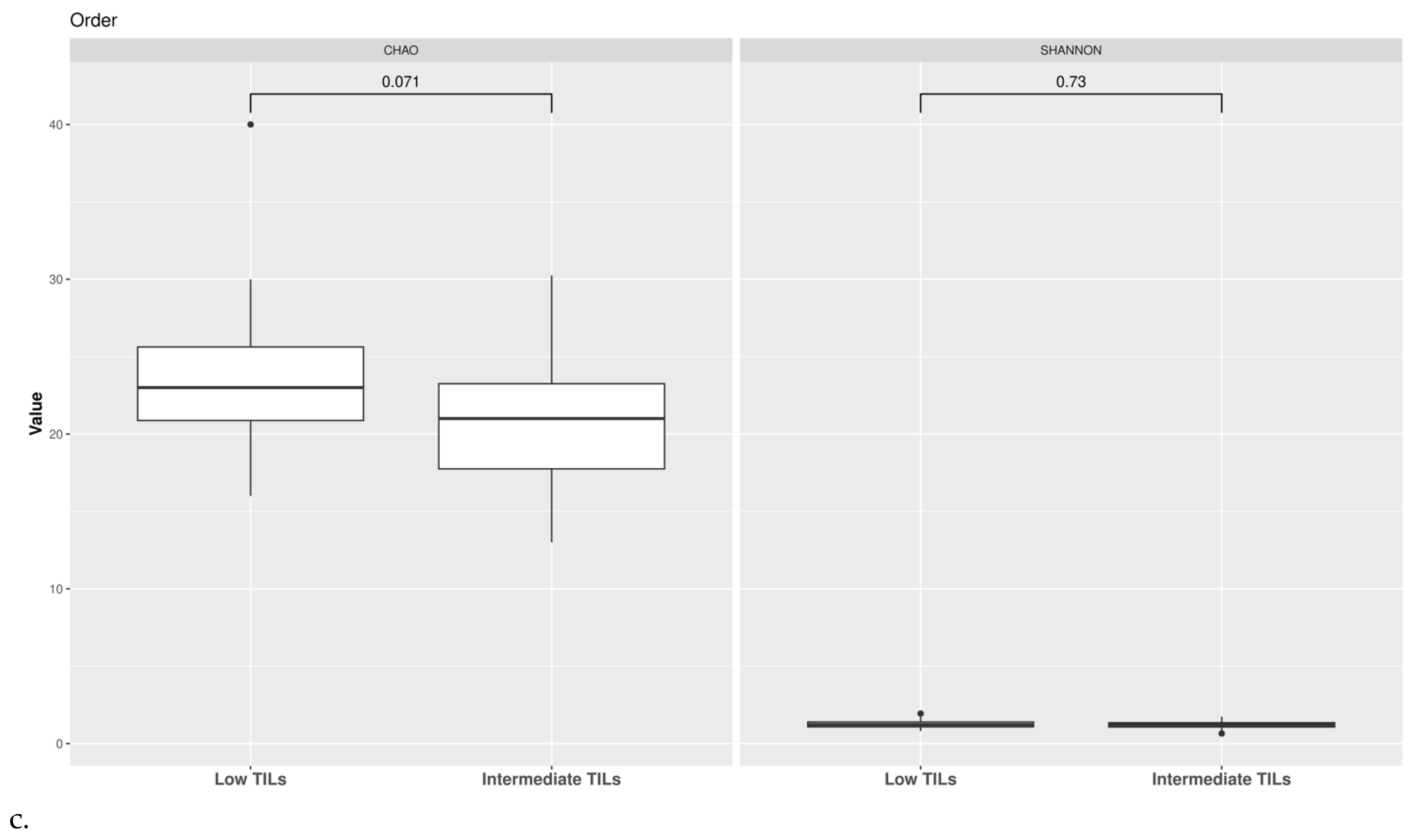
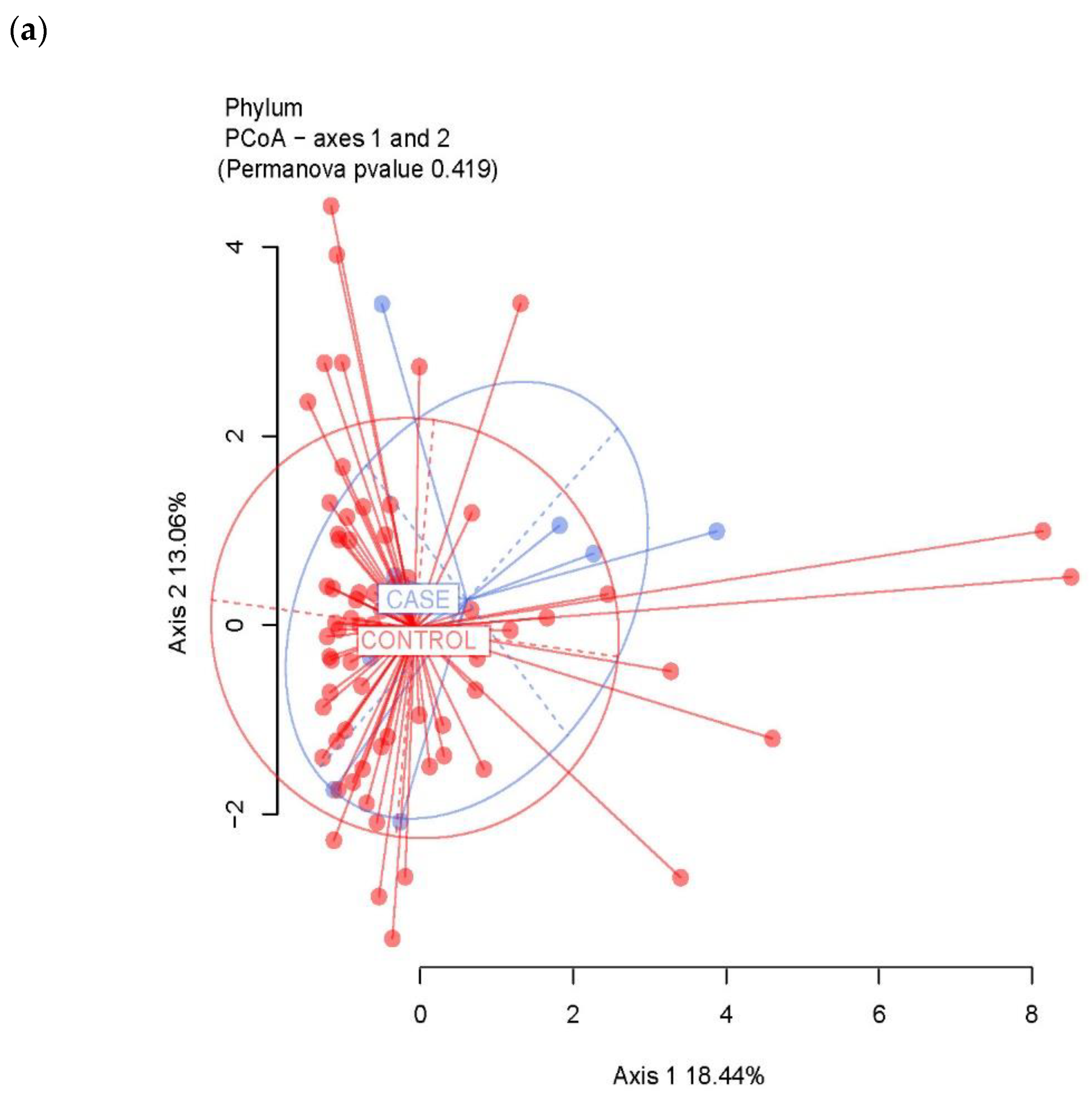
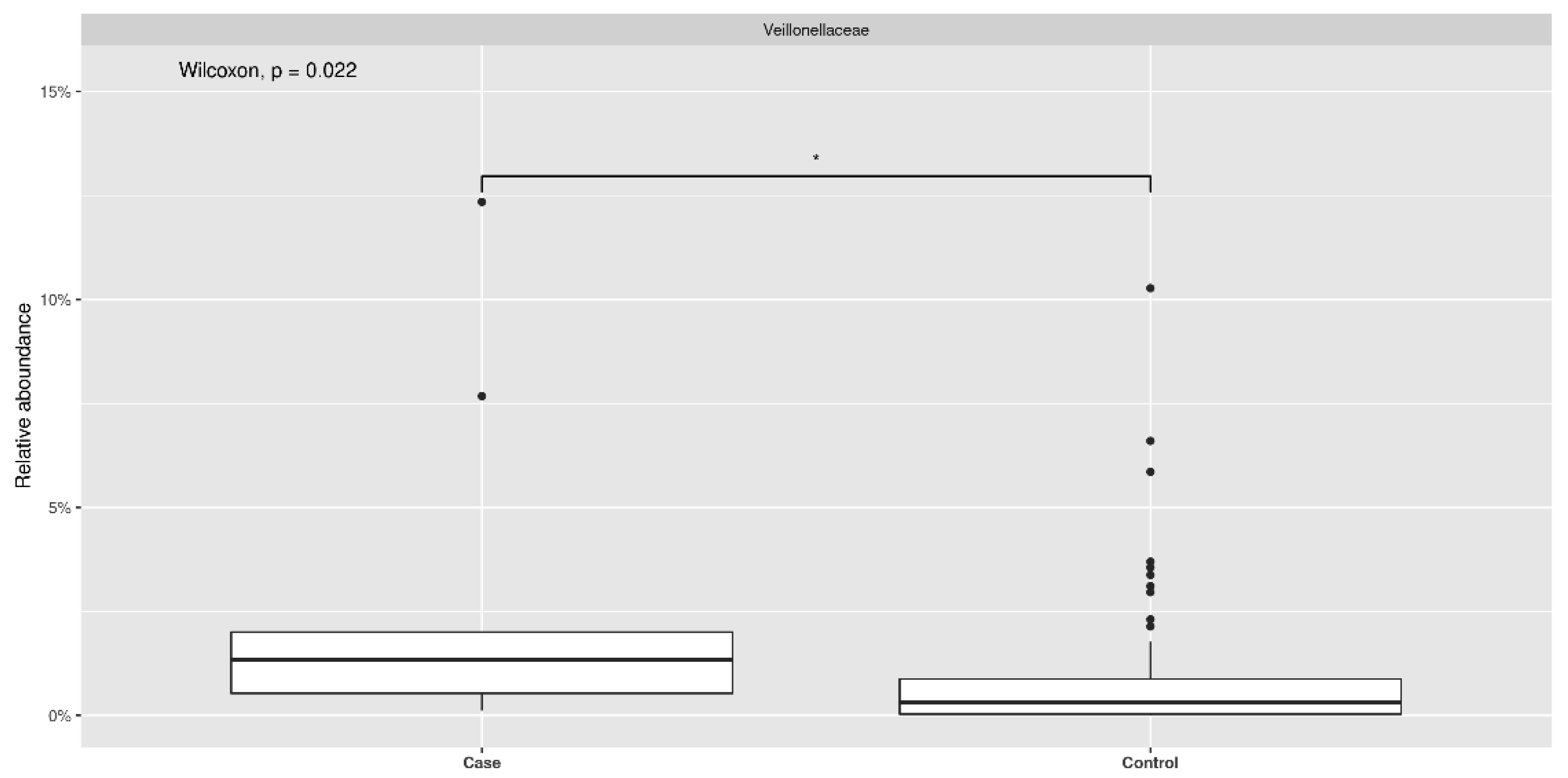
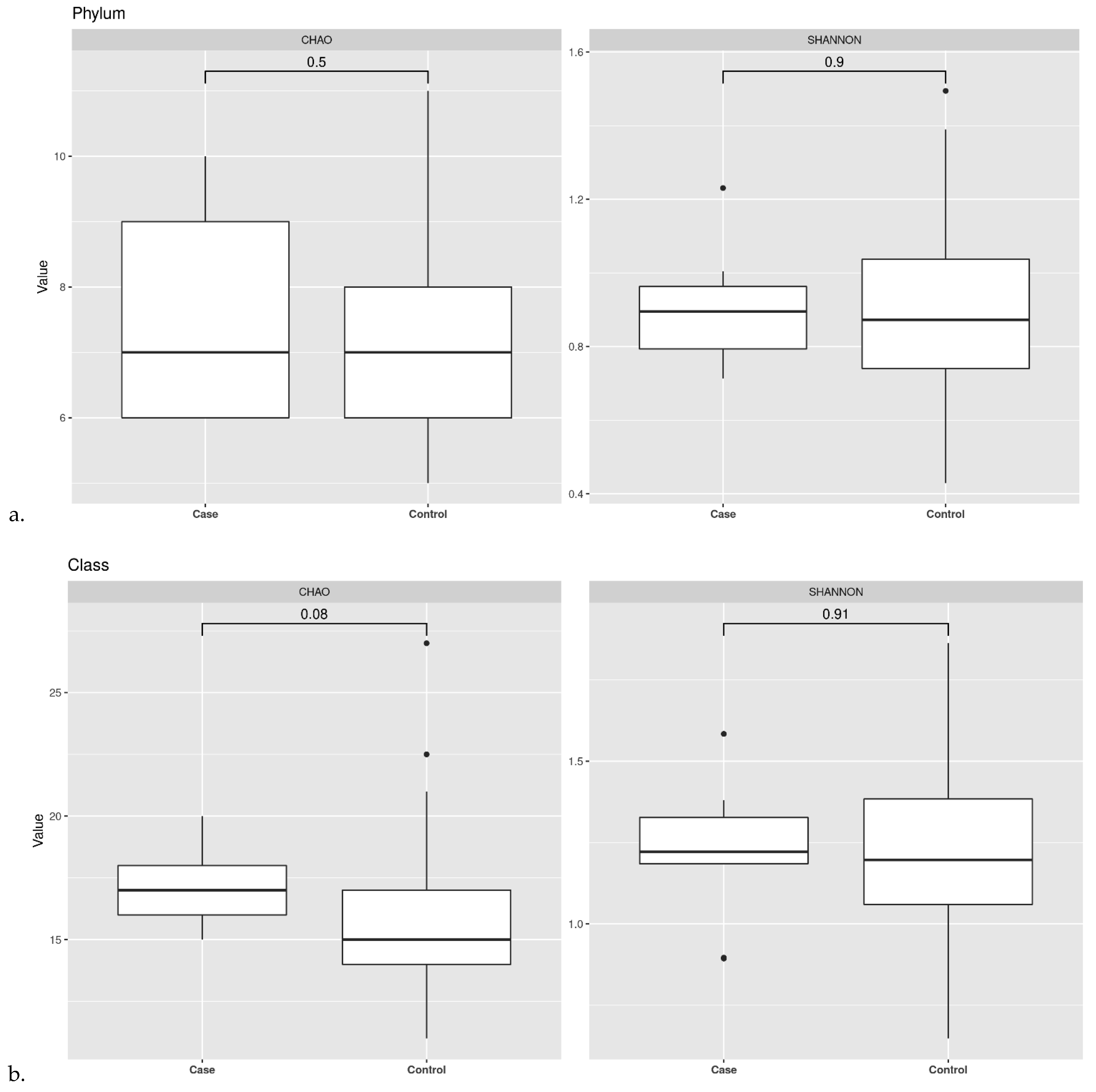
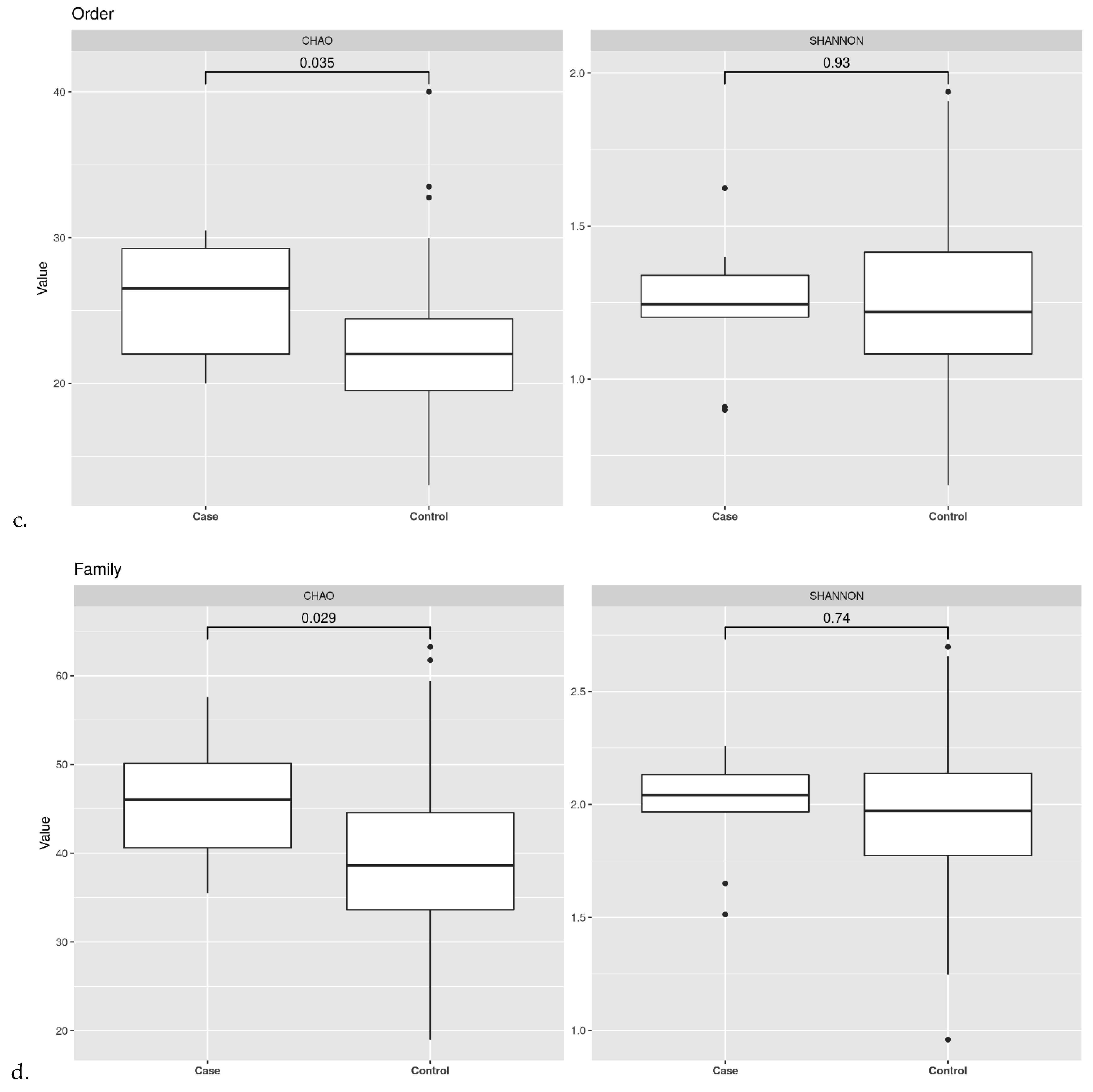
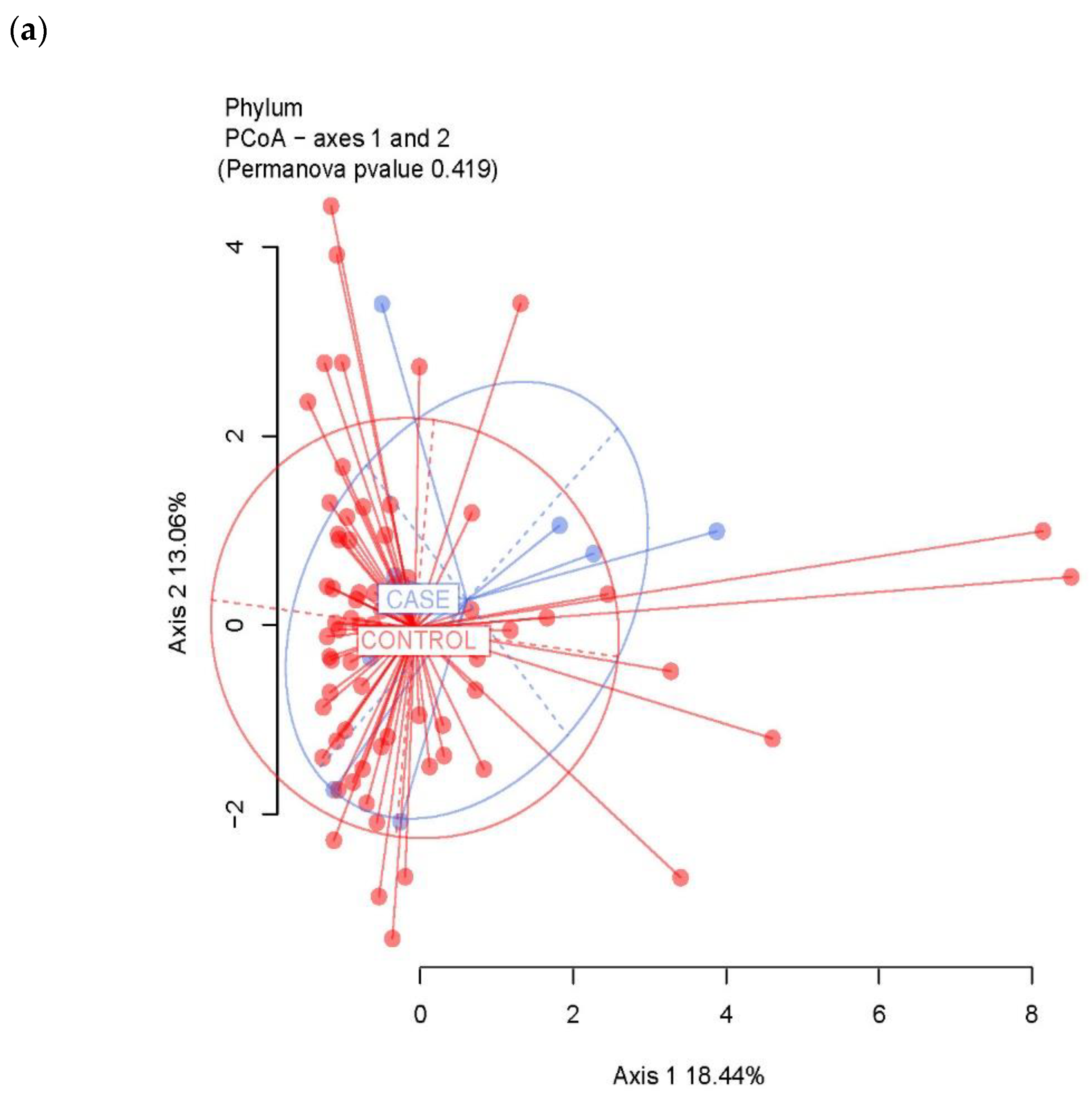
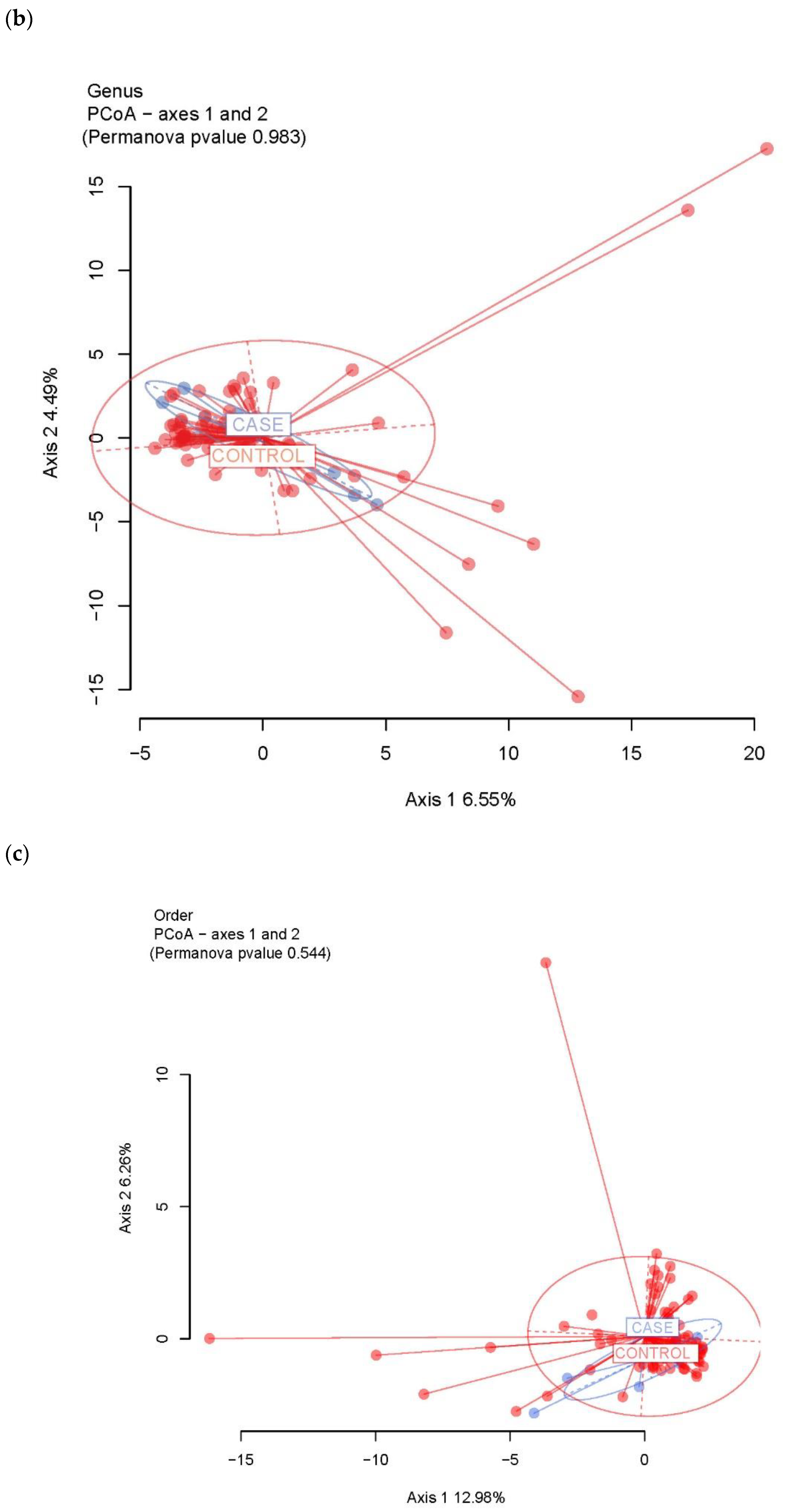
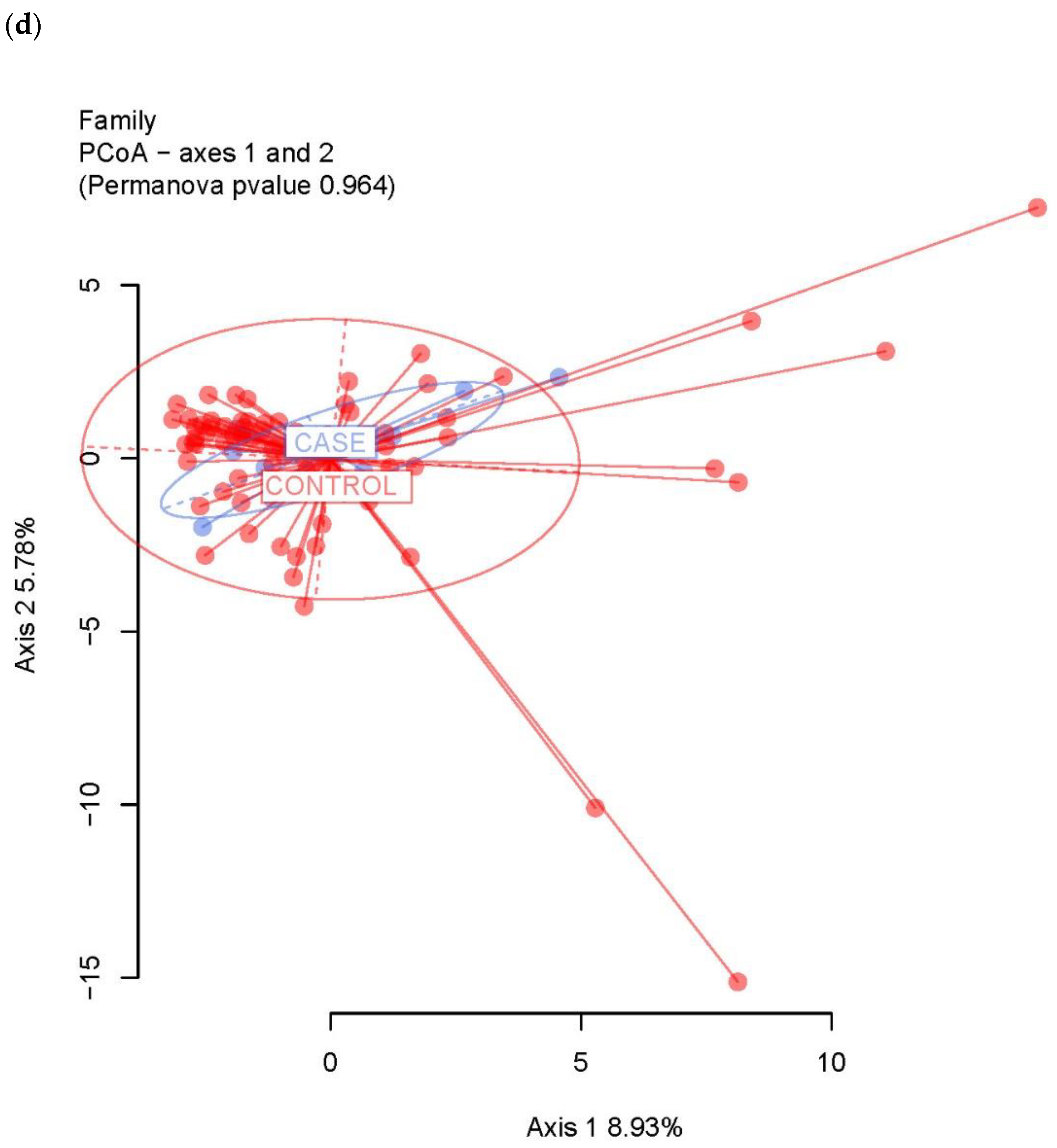
3.2. Ecological Analyses of Fecal Communities in Cases and Controls
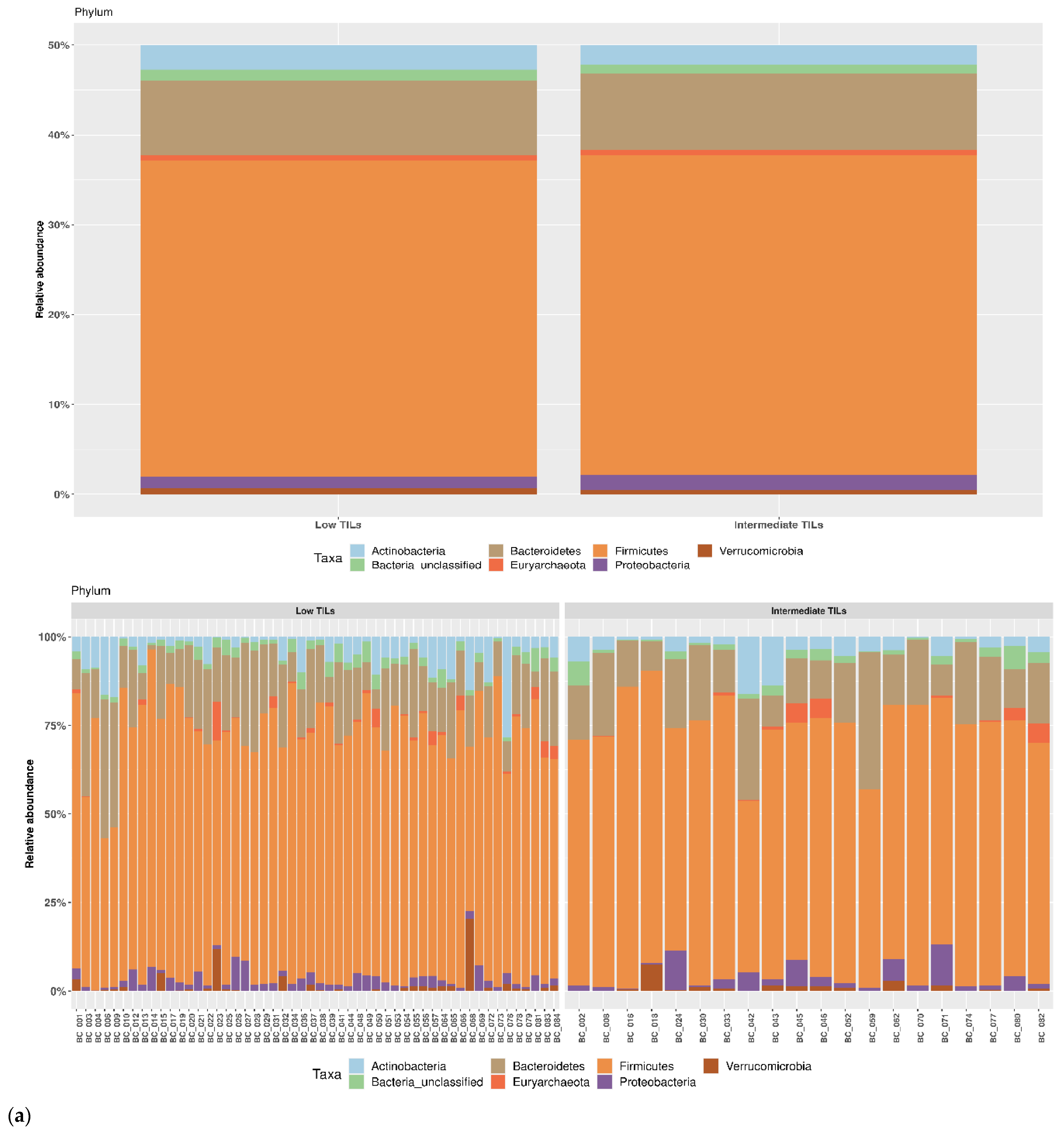
3.3. TILs Levels and Cytokine Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GLOBOCAN. 2020. Available online: https://gco.iarc.fr (accessed on 4 July 2022).
- Brown, S.B.; Hankinson, S.E. Endogenous estrogens and the risk of breast, endometrial, and ovarian cancers. Steroids 2015, 99, 8–10. [Google Scholar] [CrossRef] [PubMed]
- Arrieta, M.C.; Stiemsma, L.T.; Amenyogbe, N.; Brown, E.M.; Finlay, B. The intestinal microbiome in early life: Health and disease. Front. Immunol. 2014, 5, 427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwabe, R.; Jobin, C. The microbiome and cancer. Nat. Rev. Cancer. 2013, 13, 800–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rea, D.; Coppola, G.; Palma, G.; Barbieri, A.; Luciano, A.; Del Prete, P.; Rossetti, S.; Berretta, M.; Facchini, G.; Perdonà, S.; et al. Microbiota effects on cancer: From risks to therapies. Oncotarget 2018, 9, 17915–17927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plottel, C.S.; Blaser, M.J. Microbiome and malignancy. Cell Host Microbe 2011, 10, 324–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ervin, S.M.; Li, H.; Lim, L.; Roberts, L.R.; Liang, X.; Mani, S.; Redinbo, M.R. Gut microbial β-glucuronidases reactivate estrogens as components of the estrobolome that reactivate estrogens. J. Biol. Chem. 2019, 294, 18586–18599. [Google Scholar] [CrossRef]
- An, J.; Kwon, H.; Lim, W.; Moon, B.I. Staphylococcus aureus-Derived Extracellular Vesicles Enhance the Efficacy of Endocrine Therapy in Breast Cancer Cells. J. Clin. Med. 2022, 11, 2030. [Google Scholar] [CrossRef]
- Kwa, M.; Plottel, C.S.; Blaser, M.J.; Adams, S. The Intestinal Microbiome and Estrogen Receptor-Positive Female Breast Cancer. J. Natl. Cancer Inst. 2016, 108, djw029. [Google Scholar]
- Augusto, T.V.; Correia-da-Silva, G.; Rodrigues, C.M.P.; Teixeira, N.; Amaral, C. Acquired resistance to aromatase inhibitors: Where we stand! Endocr. Relat. Cancer 2018, 25, 283–301. [Google Scholar] [CrossRef] [Green Version]
- Fernández, M.F.; Reina-Pérez, I.; Astorga, J.M.; Rodríguez-Carrillo, A.; Plaza-Díaz, J.; Fontana, L. Breast cancer and its relationship with the microbiota. Int. J. Environ. Res. Public Health 2018, 15, 1747. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Liao, M.; Yao, Z.; Liang, W.; Li, Q.; Liu, J.; Yang, H.; Ji, Y.; Wei, W.; Tan, A.; et al. Breast cancer in postmenopausal women is associated with an altered gut metagenome. Microbiome 2018, 6, 36. [Google Scholar] [CrossRef] [PubMed]
- Buchta Rosean, C.; Bostic, R.R.; Ferey, J.C.M.; Feng, T.Y.; Azar, F.N.; Tung, K.S.; Dozmorov, M.G.; Smirnova, E.; Bos, P.D.; Rutkowski, M.R. Preexisting commensal dysbiosis is a host-intrinsic regulator of tissue inflammation and tumor cell dissemination in hormone receptor-positive breast cancer. Cancer Res. 2019, 79, 3662–3675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larkins, T.L.; Nowell, M.; Singh, S.; Sanford, G.L. Inhibition of cyclooxygenase-2 decreases breast cancer cell motility, invasion and matrix metalloproteinase expression. BMC Cancer 2006, 6, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Dubois, R.N. Cyclooxygenase-2: A potential target in breast cancer. Semin. Oncol. 2004, 31, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Geng, C.; Sang, M.; Gao, W.; Li, S.; Yang, S.; Li, Z. Effect of gastrointestinal microbiome and its diversity on the expression of tumor infiltrating lymphocytes in breast cancer. Oncol. Lett. 2019, 17, 5050–5056. [Google Scholar] [PubMed] [Green Version]
- Miyoshi, Y.; Shien, T.; Ogiya, A.; Ishida, N.; Yamazaki, K.; Horii, R.; Horimoto, Y.; Masuda, N.; Yasojima, H.; Inao, T.; et al. Associations in tumor infiltrating lymphocytes between clinicopathological factors and clinical outcomes in estrogen receptor positive/human epidermal growth factor receptor type 2 negative breast cancer. Oncol. Lett. 2019, 17, 2177–2186. [Google Scholar] [CrossRef] [Green Version]
- Cuschieri, S. The STROBE guidelines. Saudi J. Anaesth. 2019, 13 (Suppl. 1), S31–S34. [Google Scholar] [CrossRef]
- Salgado, R.; Denkert, C.; Demaria, S.; Sirtaine, N.; Klauschen, F.; Pruneri, G.; Wienert, S.; Van den Eynden, G.; Baehner, F.L.; Penault-Llorca, F.; et al. The evaluation of tumor-infiltrating lymphocytes (TILs) in breast cancer: Recommendations by an International TILs Working Group 2014. Ann. Oncol. 2015, 26, 259–271. [Google Scholar] [CrossRef]
- Demsar, J.; Curk, T.; Erjavec, A.; Gorup, C.; Hocevar, T.; Milutinovic, M.; Mozina, M.; Polajnar, M.; Toplak, M.; Staric, A.; et al. Orange: Data Mining Toolbox in Python. J. Mach. Learn. Res. 2013, 14, 2349–2353. [Google Scholar]
- Goldhirsch, A.; Winer, E.P.; Coates, A.S.; Gelber, R.D.; Piccart-Gebhart, M.; Thürlimann, B.; Senn, H.J.; Panel members. Personalizing the treatment of women with early breast cancer: Highlights of the St Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2013. Ann. Oncol. 2013, 24, 2206–2223. [Google Scholar] [CrossRef]
- Goedert, J.J.; Jones, G.; Hua, X.; Xu, X.; Yu, G.; Flores, R.; Falk, R.T.; Gail, M.H.; Shi, J.; Ravel, J.; et al. Investigation of the association between the fecal microbiota and breast cancer in postmenopausal women: A population-based case-control pilot study. J. Natl. Cancer Inst. 2015, 107, djv147. [Google Scholar] [CrossRef] [PubMed]
- Parida, S.; Sharma, D. Microbial Alterations and Risk Factors of Breast Cancer: Connections and Mechanistic Insights. Cells 2020, 9, 1091. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.P.; Xin, H.Y.; Zhang, Z.W.; Liu, C.G.; Yang, Z.; You, H.; Xin, H.W. Gut microbiota homeostasis restoration may become a novel therapy for breast cancer. Investig. New Drugs 2021, 39, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Sui, Y.; Wu, J.; Chen, J. The role of gut microbial beta-glucuronidase in estrogen reactivation and breast cancer. Front. Cell Dev. Biol. 2021, 9, 631552. [Google Scholar] [CrossRef] [PubMed]
- Laborda-Illanes, A.; Sanchez-Alcoholado, L.; Dominguez-Recio, M.E.; Jimenez-Rodriguez, B.; Lavado, R.; Comino-Méndez, I.; Alba, E.; Queipo-Ortuño, M.I. Breast and Gut Microbiota Action Mechanisms in Breast Cancer Pathogenesis and Treatment. Cancers 2020, 12, 2465. [Google Scholar] [CrossRef]
- Ubachs, J.; Ziemons, J.; Soons, Z.; Aarnoutse, R.; van Dijk, D.P.J.; Penders, J.; van Helvoort, A.; Smidt, M.L.; Kruitwagen, R.F.P.M.; Baade-Corpelijn, L.; et al. Gut microbiota and short-chain fatty acid alterations in cachectic cancer patients. J. Cachexia Sarcopenia Muscle 2021, 12, 2007–2021. [Google Scholar] [CrossRef]
- Sampsell, K.; Hao, D.; Reimer, R.A. The Gut Microbiota: A Potential Gateway to Improved Health Outcomes in Breast Cancer Treatment and Survivorship. Int. J. Mol. Sci. 2020, 21, 9239. [Google Scholar] [CrossRef]
- Available online: https://clinicaltrials.gov/ (accessed on 14 October 2022).
- Lasagna, A.; Zuccaro, V.; Ferraris, E.; Corbella, M.; Bruno, R.; Pedrazzoli, P. COVID-19 and breast cancer: May the microbiome be the issue? Future Oncol. 2021, 17, 123–126. [Google Scholar] [CrossRef]
- Mombelli, S.; Cochaud, S.; Merrouche, Y.; Garbar, C.; Antonicelli, F.; Laprevotte, E.; Alberici, G.; Bonnefoy, N.; Eliaou, J.F.; Bastid, J.; et al. IL-17A and its homologs IL-25/IL-17E recruit the c-RAF/S6 kinase pathway and the generation of pro-oncogenic LMW-E in breast cancer cells. Sci. Rep. 2015, 5, 11874. [Google Scholar] [CrossRef] [Green Version]
- Cochaud, S.; Giustiniani, J.; Thomas, C.; Laprevotte, E.; Garbar, C.; Savoye, A.M.; Curé, H.; Mascaux, C.; Alberici, G.; Bonnefoy, N.; et al. IL-17A is produced by breast cancer TILs and promotes chemoresistance and proliferation through ERK1/2. Sci. Rep. 2013, 3, 3456. [Google Scholar] [CrossRef] [Green Version]
- Savas, P.; Salgado, R.; Denkert, C.; Sotiriou, C.; Darcy, P.K.; Smyth, M.J.; Loi, S. Clinical relevance of host immunity in breast cancer: From TILs to the clinic. Nat. Rev. Clin. Oncol. 2016, 13, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Song, Y.; Zhang, Y.; Wu, H.; Chen, L.; Pang, J.; Zhou, L.; Shen, S.; Liang, Z. Prognostic significance of different molecular typing methods and immune status based on RNA sequencing in HR-positive and HER2-negative early-stage breast cancer. BMC Cancer 2022, 22, 548. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.R.; Provenzano, E.; Dawson, S.J.; Blows, F.M.; Liu, B.; Shah, M.; Earl, H.M.; Poole, C.J.; Hiller, L.; Dunn, J.A.; et al. Association between CD8+ T-cell infiltration and breast cancer survival in 12,439 patients. Ann. Oncol. 2014, 25, 1536–1543. [Google Scholar] [CrossRef] [PubMed]
- Bense, R.D.; Sotiriou, C.; Piccart-Gebhart, M.J.; Haanen, J.B.A.G.; van Vugt, M.A.T.M.; de Vries, E.G.E.; Schroder, C.P.; Fehrmann, R.S.N. Relevance of tumor-infiltrating immune cell composition and functionality for disease outcome in breast cancer. J. Natl. Cancer Inst. 2016, 109, djw192. [Google Scholar] [CrossRef]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cases (n = 8) | Controls (n = 76) | |
|---|---|---|
| Tumor histology | ||
| IDC | 8 (100%) | 63 (83%) |
| ILC | 0 | 12 (16%) |
| Other | 0 | 1 (1%) |
| Tumor grade | ||
| G1 | 0 | 9 (12%) |
| G2 | 6 (75%) | 50 (66%) |
| G3 | 2 (25%) | 17 (22%) |
| Estrogen receptor | ||
| Positive | 8 (100%) | 76 (100%) |
| Negative | 0 | 0 |
| Progesterone receptor | ||
| Positive | 8 (100%) | 76 (100%) |
| Negative | 0 | 0 |
| Ki-67 | ||
| Low | 6 (75%) | 60 (79%) |
| High | 2 (25%) | 16 (21%) |
| Molecular subtype | ||
| Luminal A | 6 (75%) | 49 (65%) |
| Luminal B | 2 (25%) | 27 (35%) |
| TILs | ||
| Low TILs | 5 (63%) | 48 (63%) |
| Intermediate TILs | 2 (25%) | 18 (24%) |
| Lymphocyte-predominant | 0 | 0 |
| nv | 1 (12%) | 10 (13%) |
| Adjuvant endocrine therapy | ||
| Anastrozole | 2 (25%) | 74 (97%) |
| Letrozole | 6 (75%) | 2 (3%) |
| Exemestane | 0 | 0 |
| Adjuvant chemotherapy | ||
| Yes | 3 (37%) | 19 (25%) |
| No | 5 (63%) | 57 (75%) |
| Comorbidities | ||
| Diabetes mellitus | 2 (25%) | 8 (10%) |
| Autoimmune disorders | 1 (12%) | 2 (3%) |
| HCV | 0 | 5 (6%) |
| HBV | 0 | 1 (1%) |
| BMI | ||
| 25–29.9 kg/m2 | 4 (50%) | 36 (47%) |
| 18.5–24.9 kg/m2 | 4 (50%) | 36 (47%) |
| >18.5 Kg/m2 | 0 | 4 (6%) |
| TILs’ Levels—All Patients (n = 84) (Median [IQR]) | TILs’ Levels—Low (n = 53) (Median [IQR]) | TILs’ Levels—Intermediate (n = 20) (Median [IQR]) | p-Value (Low vs. Intermediate) | |
|---|---|---|---|---|
| IL17 | 0.11 [0.04–0.21] | 0.15 [0.04–0.24] | 0.08 [0.02–0.11] | 0.04 |
| CD14 | 1828.50 [1081.1–2621.04] | 1894.76 [1139.61—2552.24] | 1433.70 [1049.86—2573.24] | 0.69 |
| PGE | 1207.93 [672.33–1633.71] | 1162.52 [672.33—1633.71] | 1295.24 [599.40—1633.15] | 0.93 |
| TILs Levels—All Patients (n = 84) (Median [IQR]) | TILs Levels—Controls (n = 76) (Median [IQR]) | TILs Levels—Cases (n = 8) (Median [IQR]) | p-Value (Cases vs. Controls) | |
|---|---|---|---|---|
| IL17 | 0.11 [0.04–0.21] | 0.10 [0.03–0.21] | 0.16 [0.07–0.28] | 0.38 |
| CD14 | 1828.50 [1081.1–2621.04] | 1828.50 [1085.25–2664.73] | 1947.49 [817.03–2232.15] | 0.67 |
| PGE | 1207.93 [672.33–1633.71] | 1221.90 [741.60–741.60] | 591.46 [244.85—2600.24] | 0.41 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lasagna, A.; De Amici, M.; Rossi, C.; Zuccaro, V.; Corbella, M.; Petazzoni, G.; Comandatore, F.; Sacchi, L.; Testa, G.; Ferraris, E.; et al. The Bio-Diversity and the Role of Gut Microbiota in Postmenopausal Women with Luminal Breast Cancer Treated with Aromatase Inhibitors: An Observational Cohort Study. Pathogens 2022, 11, 1421. https://doi.org/10.3390/pathogens11121421
Lasagna A, De Amici M, Rossi C, Zuccaro V, Corbella M, Petazzoni G, Comandatore F, Sacchi L, Testa G, Ferraris E, et al. The Bio-Diversity and the Role of Gut Microbiota in Postmenopausal Women with Luminal Breast Cancer Treated with Aromatase Inhibitors: An Observational Cohort Study. Pathogens. 2022; 11(12):1421. https://doi.org/10.3390/pathogens11121421
Chicago/Turabian StyleLasagna, Angioletta, Mara De Amici, Chiara Rossi, Valentina Zuccaro, Marta Corbella, Greta Petazzoni, Francesco Comandatore, Lucia Sacchi, Giorgia Testa, Elisa Ferraris, and et al. 2022. "The Bio-Diversity and the Role of Gut Microbiota in Postmenopausal Women with Luminal Breast Cancer Treated with Aromatase Inhibitors: An Observational Cohort Study" Pathogens 11, no. 12: 1421. https://doi.org/10.3390/pathogens11121421