1. Introduction
The genus
Acinetobacter represents Gram-negative rods or coccobacilli with a tendency to resist destaining, non-motility in liquid media (while surface-bound motility may be observed), catalase-positive, oxidase-negative, carbohydrate non-fermentative, chemo-organotrophic, and aerobic metabolism [
1].
A. calcoaceticus,
A. baumannii,
Acinetobacter genomic species 3 (later
A. pittii [
2]), and
Acinetobacter genomic species 13
sensu Tjernberg and Ursing “TU”, 1989 (later
A. nosocomialis [
3]), are very closely related and difficult to distinguish from each other by phenotypic properties, while biochemical differentiation from other members of the genus is possible [
4]. It has been proposed by Gerner–Smidt et al. to refer to these species as the
A. calcoaceticus-A. baumannii complex (ACB complex) [
5,
6]. Genetic studies support that the ACB complex represents a phylogenetically well-defined subgroup (clade) within the genus (cited in [
7]). Since the recognition of the ACB complex, many new
Acinetobacter species have been described, among which the clinically relevant and phylogenetically closely related
Acinetobacter seifertii [
7] and
Acinetobacter lactucae [
8] (heterotypic syn.
Acinetobacter dijkshoorniae [
9]) have been included in the complex.
As
A. calcoaceticus—in contrast to the other members of the taxonomically valid ACB complex—very rarely, if ever, causes clinically relevant infections, the clinically oriented
Acinetobacter baumannii group (ACB group) has been coined, excluding
A. calcoaceticus [
10,
11,
12].
The clinical role of the members of the ACB group resemble each other; however,
A. baumannii is more often associated with multidrug resistance and endemic spread in hospitals and with higher mortality among patients with systemic infections in case of carbapenem resistance [
13,
14]. In the United States in 2017, 8500 estimated cases and 700 estimated deaths occurred due to carbapenem-resistant
A. baumannii (CRAB) [
15]. Rice [
16] reported
A. baumannii in line with
Enterococcus faecium,
Staphylococcus aureus,
Klebsiella pneumoniae, and
Pseudomonas aeruginosa as the “ESKAPE” pathogens to emphasize that they effectively “escape” the effects of antibacterial drugs. The infection frequency is similar across Europe and Central America [
17,
18,
19,
20,
21,
22], while being even higher in Southeast Asia, India, and some countries in South America [
23,
24,
25,
26,
27,
28]. Accordingly, the WHO published a list of bacteria for which new antibiotics are urgently needed, and listed CRAB as critical and of highest priority [
29].
The general resistance of
Acinetobacter spp. to antibiotics stems in part from the very small number and permeability of porins in its outer membrane and in addition from multidrug efflux pumps [
30]. Moreover, these bacteria have the capacity to rapidly acquire additional genetic entities for resistance even from other bacterial species [
31,
32].
Acinetobacter spp. isolates have shown a complex interaction of multiple mechanisms of resistance to carbapenems, with the production of naturally occurring oxacillinases (carbapenem-hydrolyzing class D ß-lactamases, CHDLs), grouped into closely related enzymes [
33]. While
A. baumannii harbors OXA-51 (OXA-51-Like Group), a chromosome-based enzyme intrinsic to the species, other CHDLs are acquired. The predominant CHDLs (OXA-23-Like Group, OXA-24 (syn. OXA-40)-Like Group, OXA-51-Like Group, OXA-58- Like Group, OXA-143-Like Group, and OXA-235-Like Group) are responsible for the majority of phenotypic resistance to carbapenems [
12,
33,
34,
35,
36,
37]. These Ambler-class D ß-lactamases are not very active carbapenemases, and some hydrolyze imipenem and ertapenem more actively than meropenem [
38,
39,
40,
41,
42]. The presence of an insertion sequence (IS) element, such as IS
AbaI and IS
Aba9, increases the expression of the carbapenemase significantly, resulting in clinical carbapenem resistance [
43,
44,
45]. Although the CHDLs listed above are described mainly in
A. baumannii, they do spread to other
Acinetobacter species [
35,
46,
47].
Acinetobacter sp. can also acquire Ambler classes A and B carbapenemases (recently summarized in [
48]).
Based upon the prevalence, morbidity and mortality impact of CRAB infections, several organizations, e.g., the German Commission for Hospital Hygiene and Infectious Disease Prevention (KRINKO) at the Robert Koch–Institute (RKI) [
49,
50,
51] and the WHO [
52], have established guidelines for the prevention, containment, and detection of nosocomial spread. Testing for carbapenemases should be considered as a routine in all microbiology laboratories as well as the reporting to health authorities, which is explicitly recommended [
49,
53].
With the availability of new antibiotics effective against bacteria producing carbapenemases of distinct Ambler classes, it is of particular importance to rapidly inform the clinicians about the presence and Ambler class of the detected carbapenemase [
48,
54,
55,
56,
57]. In contrast to
Enterobacterales or
P. aeruginosa, carbapenemase-producing
Acinetobacer spp. almost always harbor specific CHDLs. Simply knowing about carbapenemase production may therefore be sufficient. In turn, a positive result of genetic analysis without phenotypic testing may be problematic, especially in
A. baumannii, as intrinsic OXA-51 ß-lactamases without insertion sequence-associated overexpression and mutations altering the substrate-specificity have only minimal effect on carbapenem susceptibility [
3,
12,
58].
Criteria for laboratory selection of a carbapenemase detection test on isolates or clinical samples encompass epidemiology, diagnostic performance, labor intensity, complexity, and cost. Short turnaround times are important in patient management [
55]. Assays divide into phenotypic and genotypic ones.
Nucleic acid-based assays are sensitive, specific, and may have same-day turnaround time, but (except for the remaining demand of whole genome sequencing) would not detect the emergence of new or previously uncommon carbapenemase genes and are typically associated with high costs [
59,
60].
Phenotypic tests reveal carbapenemase activity either by the uninhibited growth of a carbapenem-sensitive reporter strain exposed to a potentially degraded carbapenem, by visualized carbapenem hydrolysis, or by mass changes detected by matrix assisted-time of flight (MALDI-TOF) mass spectrometry (MS). There are currently no specific inhibitors of the prevalent Acinetobacter-associated CHDLs.
The 2017 European Committee on Antimicrobial Susceptibility Testing (EUCAST) guideline for the detection of resistance mechanisms, however, states that none of the existing phenotypic methods yield satisfactory results in the ACB group [
53]. For more advanced methods such as MALDI-TOF MS, widespread implementation seems unfeasible due to complex processing and interpretation difficulties [
61]. In
Enterobacterales, the carbapenem inactivation method (CIM) [
62,
63] has been considered favorable in a modified version (mCIM) [
64] with respect to sensitivity (97%) and specificity (99%), and was added to the Clinical and Laboratory Standards Institute M100 document [
65]. In contrast, in the corresponding EUCAST document the CIM is mentioned as a possible alternative in
Enterobacterales; however, with the accompanying information on the uncertain negative predictive value and the “disadvantage” of at least 18 h of time-to-result [
53].
Because of the lack of accurate and rapid phenotypic methods for carbapenemase detection in
Acinetobacter, variations of the classical CIM method [
62] have been tested [
64,
66,
67,
68,
69,
70,
71]. Performance characteristics of which are summarized in
Table 1.
The comparability of results is complicated by different definitions applied by various studies of what exactly is intended to be revealed by the CIM. These definitions span a spectrum from the demonstration of the activity of acquired carbapenemases (van der Zwaluw et al., 2015, Uechi et al., 2017, Simner et al., 2018, Jing et al., 2018, Vu et al., 2020, Howard et al., 2020, Cui et al., 2020) to the demonstration of the activity of chromosomal OXA-51-family, with (Liu et al., 2018, Uechi et al., 2019) or without associated insertion sequence (in Yamada et al., 2020). Since the objective of CIM was to reveal information relevant to infection control, choice of therapy, and epidemiology, supplementing the results of standard susceptibility testing, the authors of the present study retain the original intention [
62,
66] to demonstrate the presence of acquired carbapenemases and provide the accordingly transformed results of other studies in
Table 1 (bold letters), where appropriate.
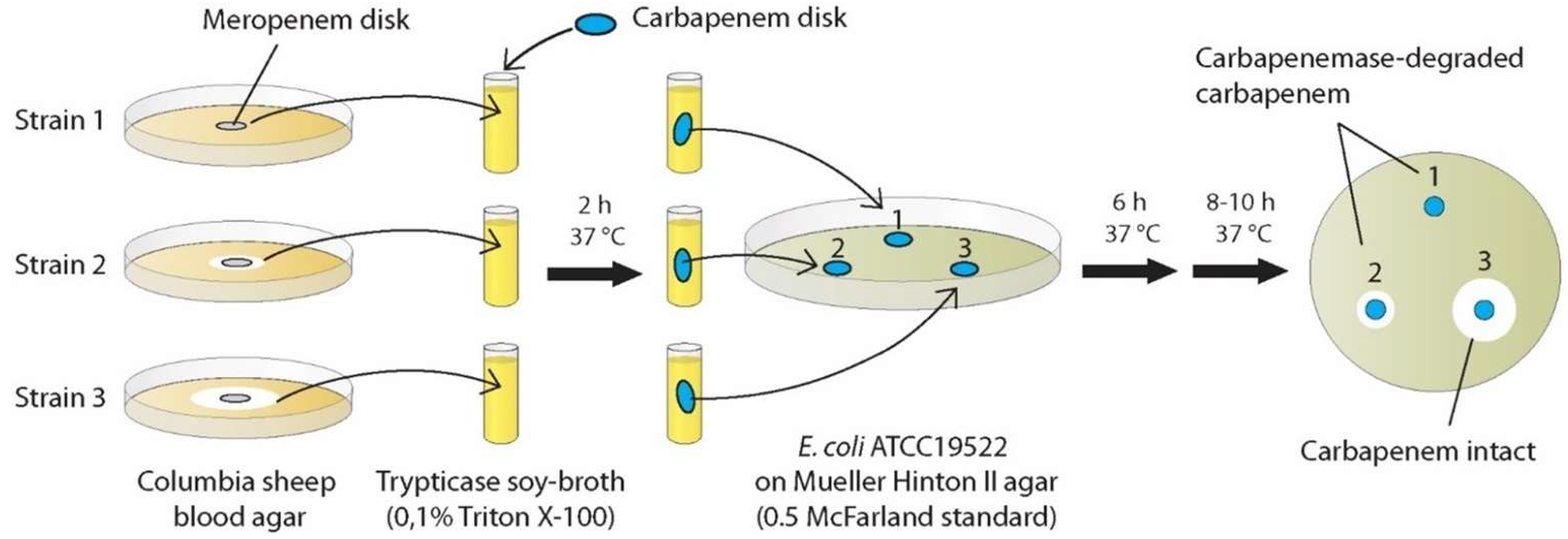
Due to the promising but still not standardized CIM methods applied so far, we conducted experiments applying another modified CIM method with the question of whether this procedure results in an early (~8 h) detection of carbapenemase production with high sensitivity and high reproducibility. This novel method consisted of advantageous features of existing protocols, which we combined and complemented with previously untested approaches: rCIM-A (
Figure 1).
3. Discussion
Given the advent of last generation anti-infective drugs, which require knowledge of resistance mechanisms of the causative organisms [
48,
54,
57] as well as the development of effective worldwide strategies against nosocomial transmission of resistant pathogens [
52], the need for rapid and accurate laboratory methods for the identification of such isolates, even in low-resource settings, has been recognized as a priority in the last years.
Existing protocols never resulted in simple methods of good performance with time-to-result shorter than 16–18 h (cf.
Table 1).
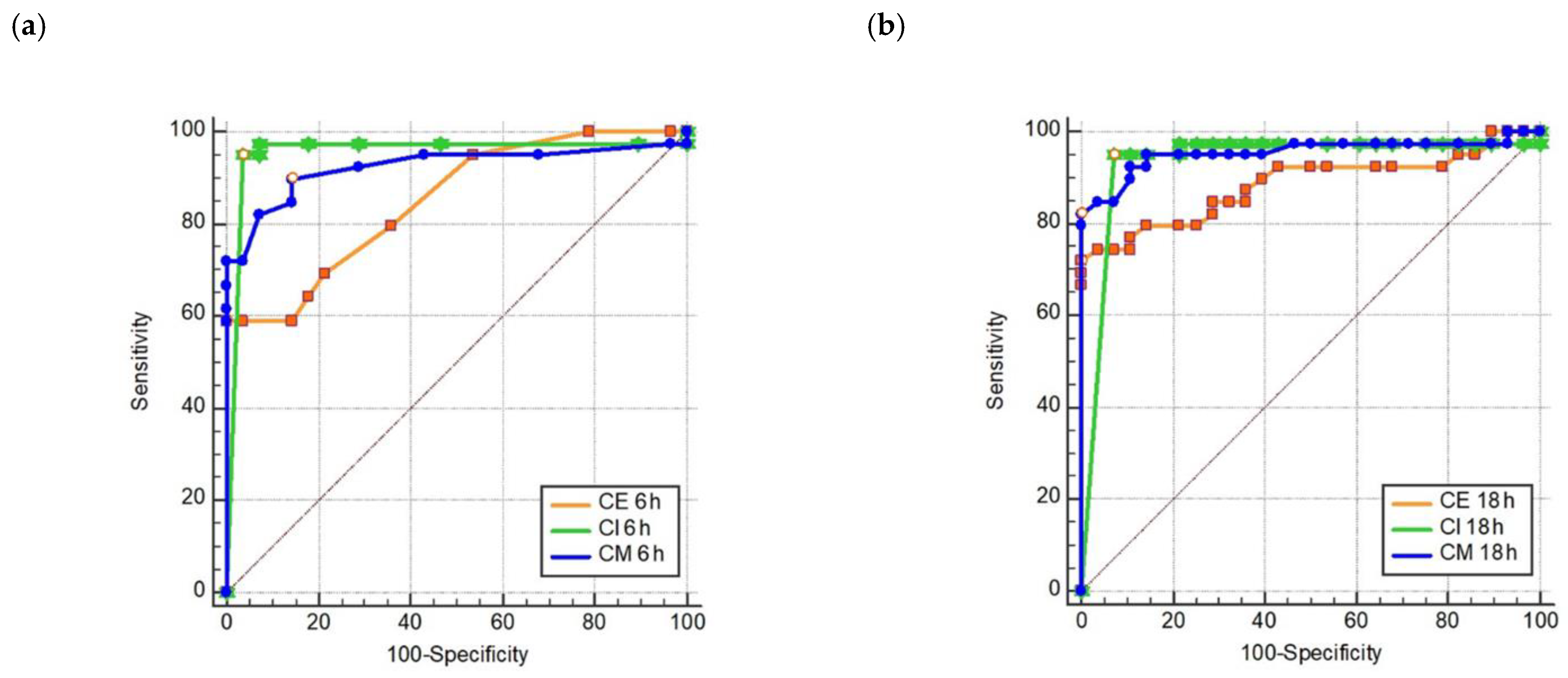
In the present study, we developed the first 8 h/“within-a-day” CIM method for routine-diagnostics application, called rCIM-A, enabling correct results within this shortened turn-around time in up to 96% of cases. Ertapenem, meropenem, and imipenem disks can be used simultaneously, as all reagents needed are together below 1 Euro per test and the processing is very simple with minimal hands-on time. It could be expected that the assay performance would not be strongly negatively affected if the three potency disks were incubated in one and the same strain suspension. The evaluation of all disks after 6 h of incubation resulted in a sensitivity near 100%. If there was no inhibition zone around the ertapenem disk after 6 h, the specificity of the positive result was 100%. This represented the main value of ertapenem disk inclusion. The highest sensitivity at 6 h was provided by the imipenem disk (97.4%) and continuing the incubation of any disk added nothing to sensitivity. If there was an inhibition zone around the ertapenem disk at 6 h, checking the ertapenem or meropenem inhibition zones (≤26 or ≤25.5 mm for positive results, respectively) at 16–18 h confirmed a negative result at 100% specificity.
In the following, we compiled an interpretative overview of CIM history and variations and compared the design of the rCIM-A, subsequently enabling the performance characteristics summarized above.
Van der Zwaluw et al. [
62], in their seminal work, used a methodological principle introduced by Masuda et al. in the year 1976 [
76] to demonstrate the existence of diffusable ß-lactamases of Gram-negative bacteria. Since this screening method for the detection of ß-lactamase activity represented a laborious diagnostic procedure, it was later simplified by the so-called Hodge test [
77]. Thereby, the principle was maintained to reveal the production of a hydrolyzing enzyme, released by a test strain into the growth agar, by the growth of a sensitive indicator strain towards a centered antibiotics disk. As the Hodge test suffered from the subjective interpretation of results and performance issues, it was in recent times no longer recommended [
53,
78]. In contrast, the novel CIM by van der Zwaluw et al. moved the hydrolysis step from growth agar to a preceding incubation step of a carbapenem disk in a suspension of the test strain. In the case of a carbapenemase-producing test strain, the potency loss of the disk was again read-out by the growth of a sensitive indicator strain [
62]. The characteristic features of this original method were suspending a 10 µL inoculation loop of culture in 400 µL of water and incubating a 10 µg meropenem disk in preparation for later biological analysis of the remaining potency. The authors concluded, with respect to
A. baumannii, which difficulties detecting low-level carbapenemase activity represented a limitation of the CIM.
In order to overcome the low sensitivity of CIM in
Acinetobacter spp. due to the low-level carbapenemase activity and low membrane permeability [
66] Uechi et al., 2017 used Tris-HCl buffer instead of trypticase soy broth (TSB) to extract carbapenemases. At a typically long time-to-result and acceptable sensitivity, the specificity let room for improvement [
72]. Tris-HCl was later shown by others to degrade meropenem rather quickly [
69].
According to the modified (m)CIM, working fine for
Enterobacterales [
64], which was added later as a CLSI method in this application [
65], Simner et al. incubated a 10 µg meropenem disk in TSB, but found—instead of a 1 µL— only a 10 µL inoculation loop of culture bringing about a modest increase in sensitivity in
A. baumannii strains. Due to the, therefore, lower specificity in this approach, the performance was described as less than ideal for
A. baumannii [
66].
Liu et al. increased sensitivity by the permeabilization of
A. baumannii complex with 0.1 % (vol/vol) Triton
TM X-100 in TSB and using a 10 µL inoculation loop of culture. The maximized performance demonstrated in this study [
79] could not be reproduced by Yamada et al. [
80] using various CHDL-producing strains. The assessed performance depends on the definition of carbapenemase-positive strains as shown in
Table 1. However, investigating the CRAB consisting of 93% OXA-23 strains and using a modification of Liu’s TCIM by incubating a 10 µg meropenem disk for 2 h in 400 µL TSB containing 0.1 % (vol/vol) Triton
TM X-100 and reading results after 16–18 h, Fan et al. recently reported sensitivity and specificity of 100%, respectively [
71].
Jing et al. deviated substantially from what was introduced as CIM by van der Zwaluw et al., since they, firstly, exchanged the disk-incubation step in a suspension of the test strain by directly smearing 1–3 colonies of the test strain onto the carbapenem-disk and, secondly, shifted the incubation phase of test strain and disk to the 16–18-h agar diffusion phase with the indicator strain (inoculated at a 1:10 dilution of 0.5 McFarland standard suspension). As they did not report on test repetitions, it is currently unknown if the direct smear method of colonies reaches the high robustness and reproducibility of the original CIM [
62]. In contrast to other approaches, this group used a 10-µg imipenem disk. Again, the reported maximized performance [
73] could not be reproduced by Yamada et al. [
69], and the reduced sensitivity in the hands of this group may have been partly due to the limited number of colonies used for the direct smear, of which some could have lost genetic resistance determinants. Further, for the original validation of sCIM only OXA-23 strains and one VIM-2 strain have been used [
66], while Yamada et al. tested various CHDL producers [
69].
Uechi et al. (2019) then tried to optimize their previous CIMTris-protocol further and were successful to some extent by using a 5 µg meropenem disk incubated for 1 h in a suspension of a 5 µL loopful (described as on-third of a 10 µL loop) of culture. Again, the time-to-result remained unabbreviated, and the authors did not address the question, if the delicate skill of measuring “one-third of a 10 µL loop” may translate into a method of high intra- and interobserver reproducibility [
74]. The reversion of the evaluation criteria to the one used by this group in their previous CIM-publication demonstrates that the sensitivity gain was on cost of specificity (
Table 1).
Vu et al., interestingly, showed a dramatical increase in performance of mCIM [
66] in
A. baumannii (83% of acquired carbapenemase-genes were comprised of OXA-23 in this study) by reverting incubation time and volume (400 µL) back to the conditions introduced by van der Zwaluw et al. while revealing that keeping TSB as incubation medium is optimal over Tris-HCl and using an unusual big inoculum of two 10 µL loopfuls of bacteria. Even the long time-to-result of the CIM and mCIM protocols was kept [
68].
Howard et al. reported the same approach differing in a refinement of sensitivity by using the prolonged suspension-incubation time of the mCIM (4 h) and of specificity by reducing the suspension-inoculum to the half of a 10 µL incubation loop. The long time-to-result of 18–24 h remained again unchanged, and we are unaware of the reproducibility of the method, as well as of the number of carbapenemase-negative
A. baumannii strains included in the study [
75].
Cui et al. presented a comparatively costly approach of seemingly shortened time-to-result by a CIM-related methodology, which reminds us—with respect to the laborious in-house concept—of Masuda’s 1976 screening method [
80]. Their starting point is the overnight liquid culture of the test strain, i.e., not from the common agar diffusion susceptibility test. The rest-potency of a 10 µg imipenem disk is indicated by growth of
Bacillus stearothermophilus 7453 on an in-house chromogenic plate at 60 °C. All CHDLs in this study were comprised of OXA-23 [
70].
Yamada et al. showed that 0.5 M Tris-HCl as an incubation matrix rapidly degrades meropenem in the antibiotic disk, while no substrate was degraded using 3-morpholinopropane-1-sulfonic acid (MOPS), which was therefore chosen for LCIM. In this method, the concentration of Triton
TM X-100 was increased to lytic levels of 2% (vol/vol) in a 4-h incubation step with the disk, followed by a standard incubation time with the indicator strain. In an attempt to maximize sensitivity, this group reported on the detection of OXA-51-positive strains without IS
Aba1 [
69], which clearly misses the point as those detections essentially just reveal the presence of
A. baumannii per se and should thus be regarded as false-positives.
Characteristic similarities and differences between existing and the novel method are briefly summarized below.
To make use of potential induction of the weak
A. baumannii group carbapenemases colonies were taken from the edge of the inhibition zones [
66] around meropenem disks used for routine-AST.
As smaller volumes were shown to reduce sensitivity at better specificity in
Acinetobacter too much [
66,
67,
75], the volume contained in a 10 µL inoculation loop was used for preparing a suspension in 400 µL TSB containing 0.1% (vol/vol) Triton
TM X-100. The mildly membrane-permeabilizing 0.1% (vol/vol) Triton
TM X-100 [
67,
71] plus TSB (carbapenem-protective, [
68]) approach was chosen, since the disk incubation step in water-suspension [
62] or TSB [
66] did not result in sufficient bacterial-borne lytic activity [
62] and Tris-HCl extraction was shown to degrade the carbapenem unspecifically [
69].
Representing a new CIM modification, a 10 µg potency disk of ertapenem, meropenem, and imipenem was added to test-strain suspensions. One may ask why one should deviate from the original CIM protocol [
62,
66,
67] by using imipenem or, in particular, the clinically ineffective ertapenem in
A. baumannii-CIM. Importantly, the most prevalent carbapenemases in
A. baumannii (CHDLs) are low-activity enzymes, being comparatively difficult to detect phenotypically; to this end these enzymes should therefore be combined with substrates, they do most actively convert. Conversely, the in vitro or clinical effectivity of the carbapenemase-substrates (assay-carbapenems) on
A. baumannii have no impact on the test performance at all. The carbapenem does not have to inwardly cross the
Acinetobacter outer membrane, but awaits, on the contrary, the exit of carbapenemases. The read-out strain (
Escherichia coli ATCC 25922) only has to be susceptible. Imipenem [
72] and, more, ertapenem [
42] are most effectively hydrolyzed by CHDLs and were, for sensitivity and assay-rapidity reasons, integrated in the rCIM-A protocol.
Due to above-described optimizations a read-out was performed as early as after 6 h and after additional 8–10 h of incubation, enabling the potential shortest CIM protocol suitable for routine-use so far (
Figure 1).
Although the substrate-specific activities of the predominating Ambler class D carbapenemases in
Acinetobacter spp. have been known for a long time, the present study describes, irrespective of two quite departing variations [
70,
73], the only classical, and hence robust and straightforward, CIM method using imipenem. The inclusion of two other carbapenem potency disks in the protocol may be advantageous in case of carbapenemases of the Ambler classes A and B, which also occur in the
A. baumannii group with varying prevalence globally. To our knowledge, ertapenem has never been tested in a CIM approach previously.
Because of the high NPV revealed, the rCIM-A seems particularly appropriate in areas with lower CRAB frequencies and when PCR tests (NAATs) are not available because of high costs, required technical skills, and/or laboratory compartments. In addition, PCRs are not free of hypotheses and may not detect new or locally rare genetic variants.
The newly described phenomenon of “carryover microsatellites” may be helpful even in other variants of CIM in order not to overlook true-positive results.
Further studies may investigate the ability of included specific inhibitors of Ambler class A and B carbapenemases to discriminate these from CHDLs, in order to inform appropriate therapeutic decisions in areas where the class spectrum of carbapenemases in A. baumanii group is more diverse.
As for every diagnostic laboratory method, the validation in a clinical study addressing the clinical benefit would be desirable.


 ,
, 





{kind=link}
{kind=link}
{kind=link}