
Determinants of Virus Variation, Evolution, and Host Adaptation



Abstract
:1. Introduction
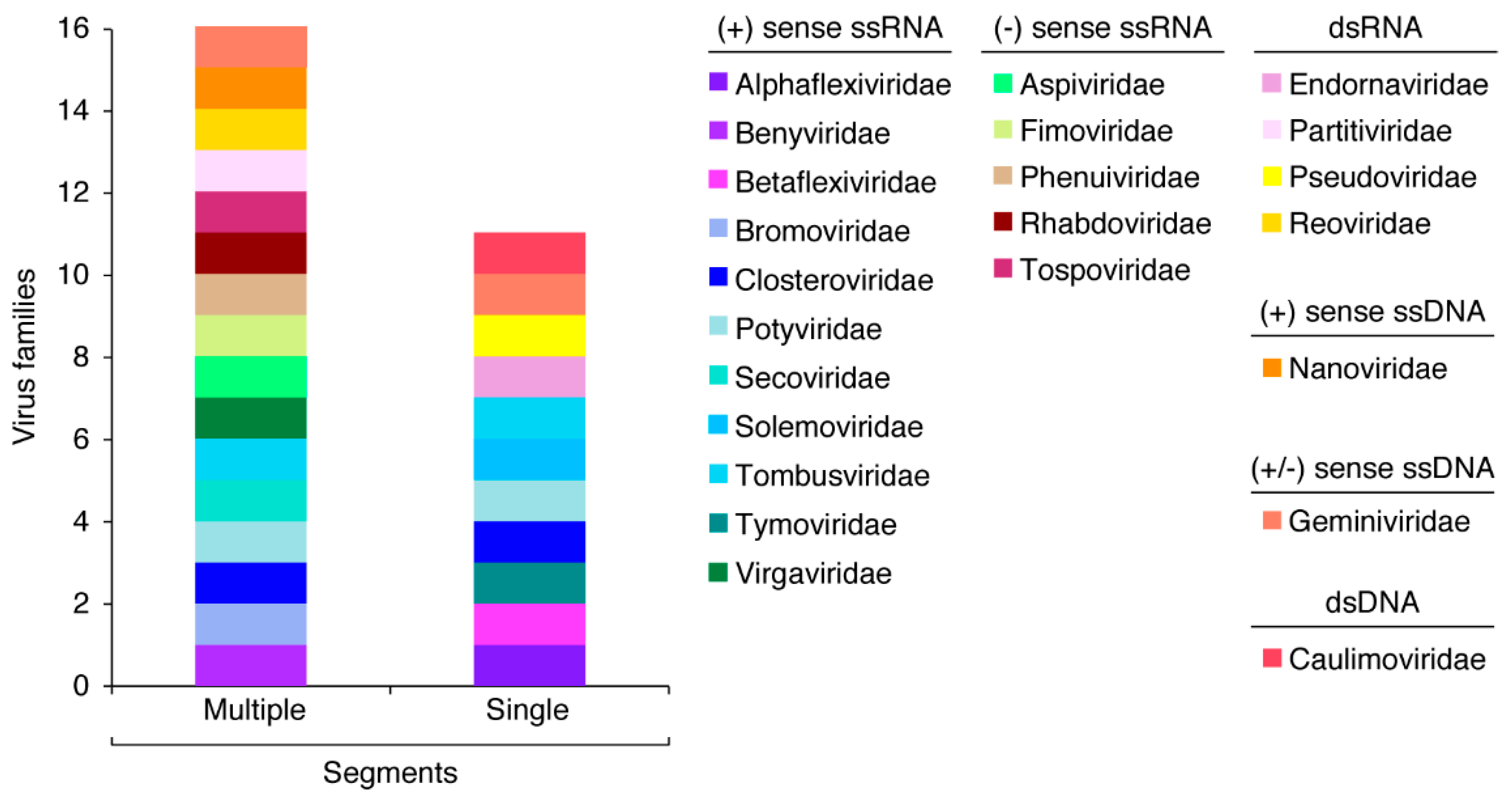
1.1. Plant Virus Genome Organization
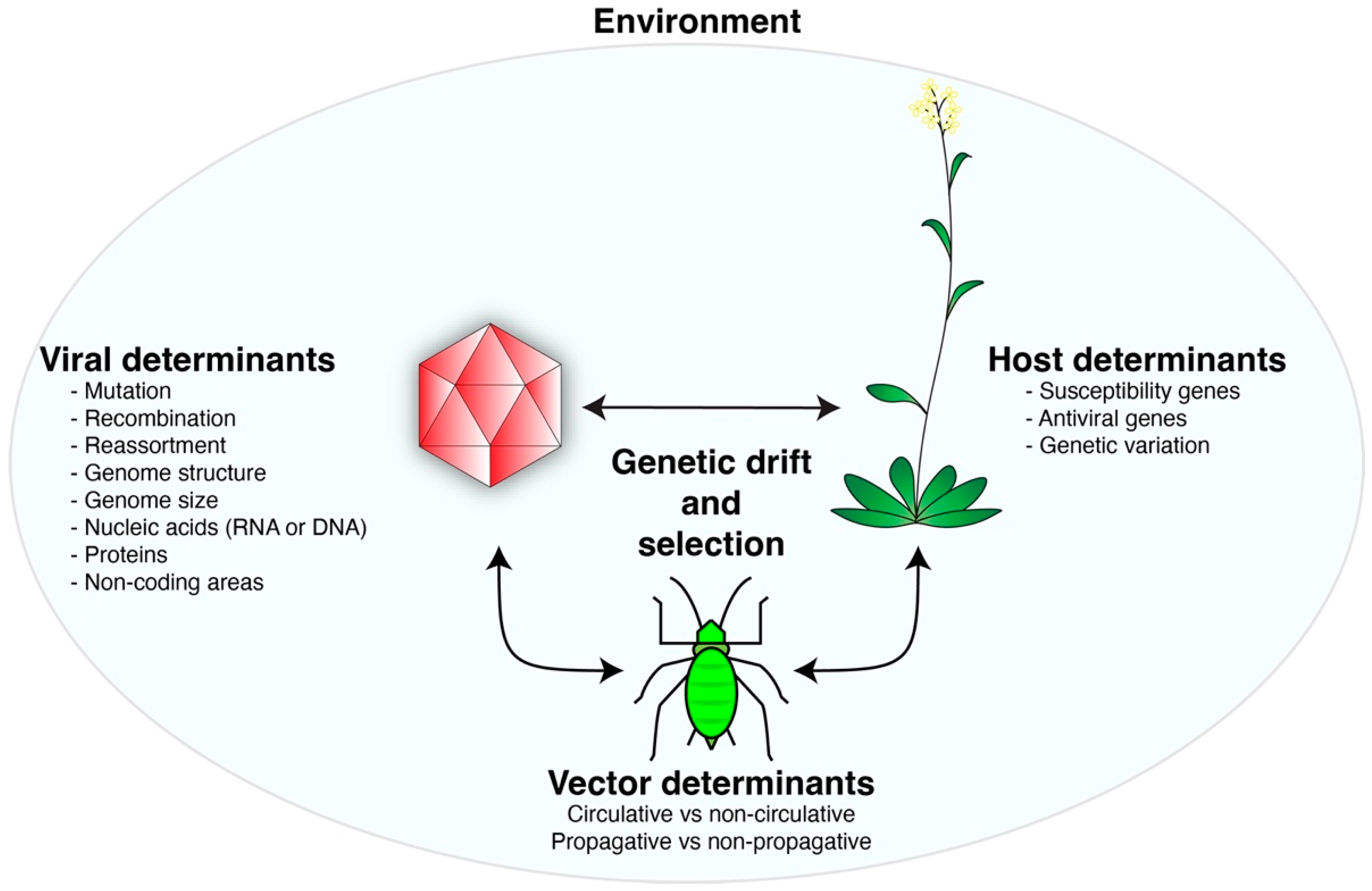
1.2. Virus-Plant Co-Evolution
1.3. Virus-Vector Co-Evolution
1.4. The Environment and Virus Evolution
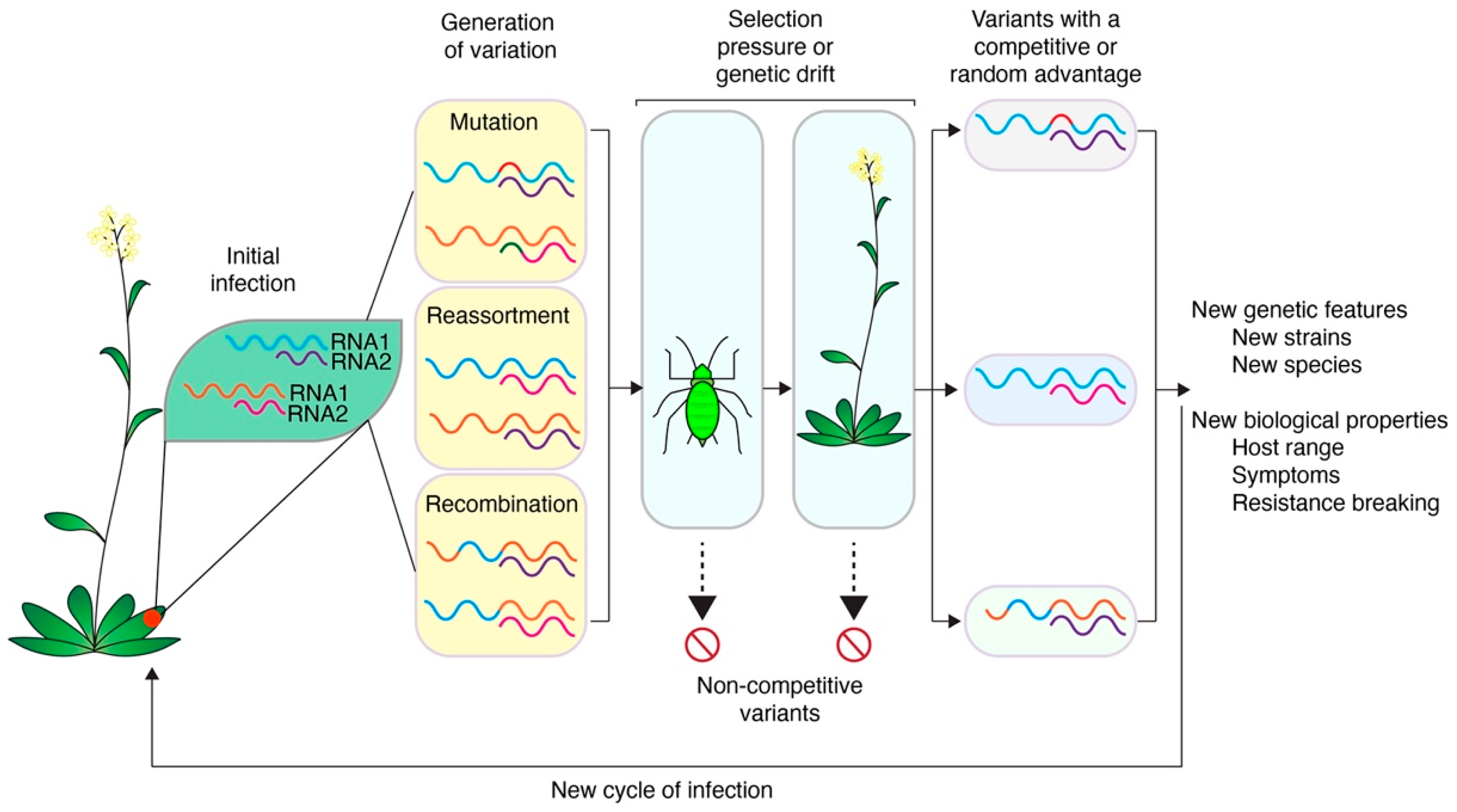
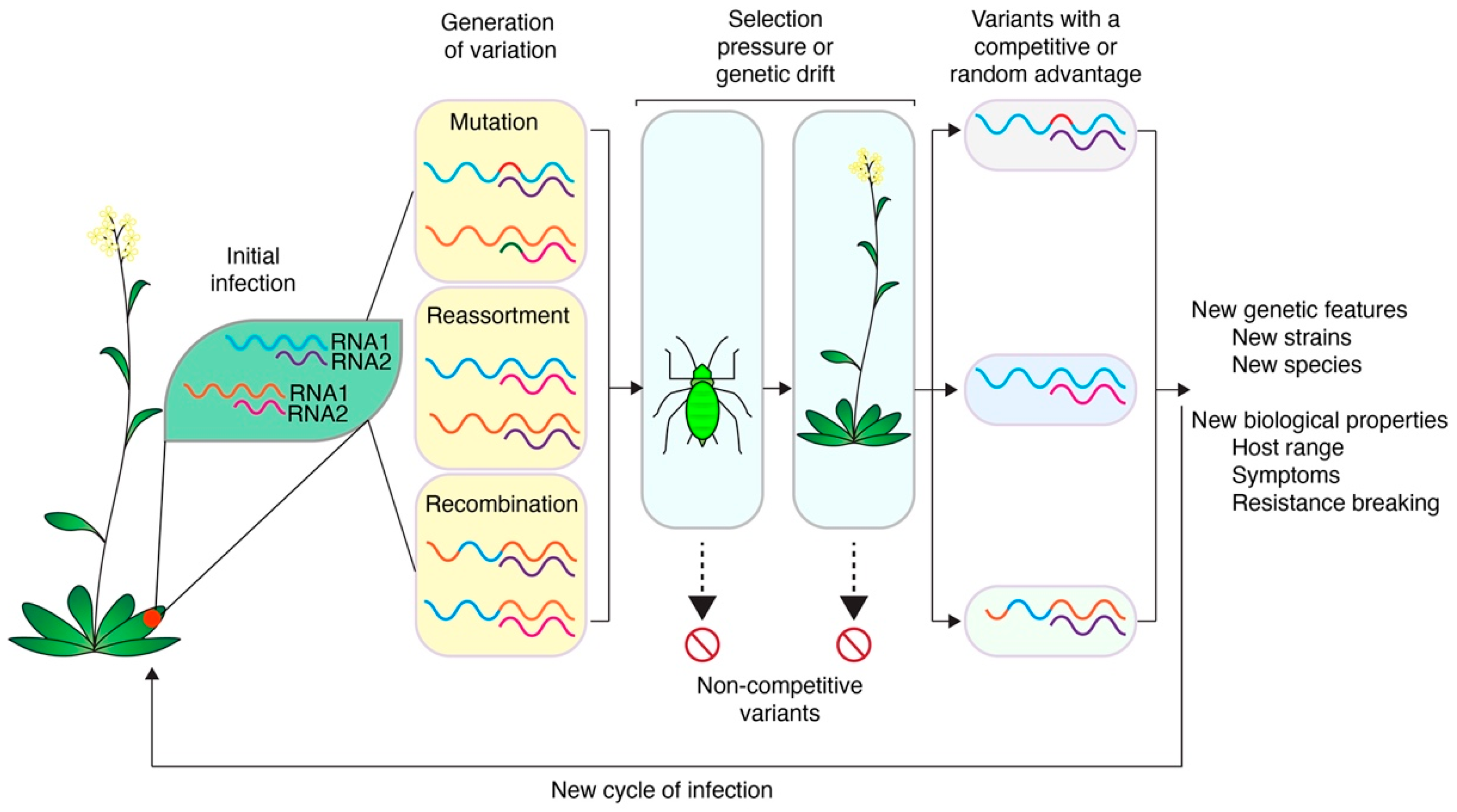
1.5. Variation, Bottlenecks, Genetic Drift, and Selection Pressure in Virus Populations
1.6. Host Adaptation
2. Mutation
3. Reassortment
4. Recombination
5. Genetically Stationary and Transient Viruses
6. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Godfray, H.C.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef]
- Anderson, P.K.; Cunningham, A.A.; Patel, N.G.; Morales, F.J.; Epstein, P.R.; Daszak, P. Emerging infectious diseases of plants: Pathogen pollution, climate change and agrotechnology drivers. Trends Ecol. Evol. 2004, 19, 535–544. [Google Scholar] [CrossRef]
- Sastry, K.S.; Zitter, T.A. Management of Virus and Viroid Diseases of Crops in the Tropics. In Plant Virus and Viroid Diseases in the Tropics: Volume 2: Epidemiology and Management; Sastry, K.S., A. Zitter, T., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 149–480. [Google Scholar]
- Jones, R.A.C.; Naidu, R.A. Global Dimensions of Plant Virus Diseases: Current Status and Future Perspectives. Annu. Rev. Virol. 2019, 6, 387–409. [Google Scholar] [CrossRef]
- Jones, R.A.C. Chapter Three—Future Scenarios for Plant Virus Pathogens as Climate Change Progresses. In Advances in Virus Research; Kielian, M., Maramorosch, K., Mettenleiter, T.C., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 95, pp. 87–147. [Google Scholar]
- Cooper, J.I.; Kuehne, T.; Polischuk, V.P. Virus Diseases and Crop Biosecurity: NATO Sponsered Workshop, Held in Kiev, Ukraine during 4–7 May 2005; Cooper, J.I., Ed.; Springer: Dordrecht, The Netherlands, 2006; p. 148. [Google Scholar]
- Trebicki, P. Climate change and plant virus epidemiology. Virus. Res. 2020, 286, 198059. [Google Scholar] [CrossRef]
- Elena, S.F.; Fraile, A.; García-Arenal, F. Chapter Three—Evolution and Emergence of Plant Viruses. In Advances in Virus Research; Maramorosch, K., Murphy, F.A., Eds.; Academic Press: Cambridge, MA, USA, 2014; Volume 88, pp. 161–191. [Google Scholar]
- Makinen, K.; Hafren, A. Intracellular coordination of potyviral RNA functions in infection. Front. Plant Sci. 2014, 5, 110. [Google Scholar] [CrossRef]
- Garcia-Ruiz, H. Susceptibility Genes to Plant Viruses. Viruses 2018, 10, 484. [Google Scholar] [CrossRef]
- Fereres, A.; Raccah, B. Plant Virus Transmission by Insects. In eLS; John, W., Sons, L., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2015; pp. 1–12. [Google Scholar]
- Heinlein, M. Plant virus replication and movement. Virology 2015, 479–480, 657–671. [Google Scholar] [CrossRef]
- Garcia-Ruiz, H. Host factors against plant viruses. Mol. Plant Pathol. 2019, 20, 1588–1601. [Google Scholar] [CrossRef]
- Hafren, A.; Ustun, S.; Hochmuth, A.; Svenning, S.; Johansen, T.; Hofius, D. Turnip Mosaic Virus Counteracts Selective Autophagy of the Viral Silencing Suppressor HCpro. Plant Physiol. 2018, 176, 649–662. [Google Scholar] [CrossRef]
- Palukaitis, P.; Yoon, J.Y. R gene mediated defense against viruses. Curr. Opin. Virol. 2020, 45, 1–7. [Google Scholar] [CrossRef]
- Bendahmane, A.; Kanyuka, K.; Baulcombe, D.C. The Rx gene from potato controls separate virus resistance and cell death responses. Plant Cell 1999, 11, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Brommonschenkel, S.H.; Frary, A.; Frary, A.; Tanksley, S.D. The broad-spectrum tospovirus resistance gene Sw-5 of tomato is a homolog of the root-knot nematode resistance gene Mi. Mol. Plant Microbe Interact. 2000, 13, 1130–1138. [Google Scholar] [CrossRef] [PubMed]
- Hajimorad, M.R.; Hill, J.H. Rsv1-mediated resistance against soybean mosaic virus-N is hypersensitive response-independent at inoculation site, but has the potential to initiate a hypersensitive response-like mechanism. Mol. Plant Microbe Interact. 2001, 14, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Heo, K.J.; Kwon, S.J.; Kim, M.K.; Kwak, H.R.; Han, S.J.; Kwon, M.J.; Rao, A.L.N.; Seo, J.K. Newly emerged resistance-breaking variants of cucumber mosaic virus represent ongoing host-interactive evolution of an RNA virus. Virus Evol. 2020, 6, veaa070. [Google Scholar] [CrossRef] [PubMed]
- Desbiez, C.; Domingo-Calap, M.L.; Pitrat, M.; Wipf-Scheibel, C.; Girardot, G.; Ferriol, I.; Lopez-Moya, J.J.; Lecoq, H. Specificity of Resistance and Tolerance to Cucumber Vein Yellowing Virus in Melon Accessions and Resistance Breaking with a Single Mutation in VPg. Phytopathology 2022, 112, 1185–1191. [Google Scholar] [CrossRef]
- Rousseau, E.; Bonneault, M.; Fabre, F.; Moury, B.; Mailleret, L.; Grognard, F. Virus epidemics, plant-controlled population bottlenecks and the durability of plant resistance. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180263. [Google Scholar] [CrossRef]
- Crespo-Bellido, A.; Hoyer, J.S.; Dubey, D.; Jeannot, R.B.; Duffy, S. Interspecies Recombination Has Driven the Macroevolution of Cassava Mosaic Begomoviruses. J. Virol. 2021, 95, e0054121. [Google Scholar] [CrossRef]
- Domingo, E.; Garcia-Crespo, C.; Perales, C. Historical Perspective on the Discovery of the Quasispecies Concept. Annu. Rev. Virol. 2021, 8, 51–72. [Google Scholar] [CrossRef]
- Domingo, E. Quasispecies: Concept and Implications for Virology, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Ali, A.; Roossinck, M.J. Analysis of quasispecies variation in single and mixed viral infection. Virus Evol. 2017, 3, vex037. [Google Scholar] [CrossRef]
- Alcaide, C.; Sardanyes, J.; Elena, S.F.; Gomez, P. Increasing temperature alters the within-host competition of viral strains and influences virus genetic variability. Virus Evol. 2021, 7, veab017. [Google Scholar] [CrossRef]
- Roossinck, M.; Ali, A. Mechanisms of plant virus evolution and identification of genetic bottlenecks: Impact on diseasemanagement. Biotechnol. Plant Dis. Manag. 2007, 109–124. [Google Scholar] [CrossRef]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Alfenas-Zerbini, P.; Davison, A.J.; Dempsey, D.M.; Dutilh, B.E.; Garcia, M.L.; et al. Changes to virus taxonomy and to the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2021). Arch. Virol. 2021, 166, 2633–2648. [Google Scholar] [CrossRef] [PubMed]
- Hull, R. Matthews’ Plant Virology; Academic Press: Cambridge, MA, USA, 2002. [Google Scholar]
- Moury, B.; Fabre, F.; Hebrard, E.; Froissart, R. Determinants of host species range in plant viruses. J. Gen. Virol. 2017, 98, 862–873. [Google Scholar] [CrossRef] [PubMed]
- Sicard, A.; Michalakis, Y.; Gutierrez, S.; Blanc, S. The Strange Lifestyle of Multipartite Viruses. PLoS Pathog. 2016, 12, e1005819. [Google Scholar] [CrossRef] [PubMed]
- Traore, O.; Pinel-Galzi, A.; Issaka, S.; Poulicard, N.; Aribi, J.; Ake, S.; Ghesquiere, A.; Sere, Y.; Konate, G.; Hebrard, E.; et al. The adaptation of Rice yellow mottle virus to the eIF(iso)4G-mediated rice resistance. Virology 2010, 408, 103–108. [Google Scholar] [CrossRef]
- Agudelo-Romero, P.; de la Iglesia, F.; Elena, S.F. The pleiotropic cost of host-specialization in Tobacco etch potyvirus. Infect. Genet. Evol. 2008, 8, 806–814. [Google Scholar] [CrossRef]
- Gong, Y.N.; Tang, R.Q.; Zhang, Y.; Peng, J.; Xian, O.; Zhang, Z.H.; Zhang, S.B.; Zhang, D.Y.; Liu, H.; Luo, X.W.; et al. The NIa-Protease Protein Encoded by the Pepper Mottle Virus Is a Pathogenicity Determinant and Releases DNA Methylation of Nicotiana benthamiana. Front. Microbiol. 2020, 11, 102. [Google Scholar] [CrossRef]
- Koonin, E.V.; Dolja, V.V. A virocentric perspective on the evolution of life. Curr. Opin. Virol. 2013, 3, 546–557. [Google Scholar] [CrossRef]
- Wilke, C.O.; Wang, J.L.; Ofria, C.; Lenski, R.E.; Adami, C. Evolution of digital organisms at high mutation rates leads to survival of the flattest. Nature 2001, 412, 331–333. [Google Scholar] [CrossRef]
- Hillung, J.; Cuevas, J.M.; Valverde, S.; Elena, S.F. Experimental evolution of an emerging plant virus in host genotypes that differ in their susceptibility to infection. Evolution 2014, 68, 2467–2480. [Google Scholar] [CrossRef] [Green Version]
- Wallis, C.M.; Stone, A.L.; Sherman, D.J.; Damsteegt, V.D.; Gildow, F.E.; Schneider, W.L. Adaptation of plum pox virus to a herbaceous host (Pisum sativum) following serial passages. J. Gen. Virol. 2007, 88, 2839–2845. [Google Scholar] [CrossRef] [PubMed]
- Obenauer, J.C.; Denson, J.; Mehta, P.K.; Su, X.; Mukatira, S.; Finkelstein, D.B.; Xu, X.; Wang, J.; Ma, J.; Fan, Y.; et al. Large-scale sequence analysis of avian influenza isolates. Science 2006, 311, 1576–1580. [Google Scholar] [CrossRef] [PubMed]
- Nigam, D.; Garcia-Ruiz, H. Variation Profile of the Orthotospovirus Genome. Pathogens 2020, 9, 521. [Google Scholar] [CrossRef] [PubMed]
- Nigam, D.; LaTourrette, K.; Souza, P.F.N.; Garcia-Ruiz, H. Genome-Wide Variation in Potyviruses. Front. Plant Sci. 2019, 10, 1439. [Google Scholar] [CrossRef] [PubMed]
- Korner, C.J.; Klauser, D.; Niehl, A.; Dominguez-Ferreras, A.; Chinchilla, D.; Boller, T.; Heinlein, M.; Hann, D.R. The immunity regulator BAK1 contributes to resistance against diverse RNA viruses. Mol. Plant Microbe Interact. 2013, 26, 1271–1280. [Google Scholar] [CrossRef]
- Hebrard, E.; Poulicard, N.; Gerard, C.; Traore, O.; Wu, H.C.; Albar, L.; Fargette, D.; Bessin, Y.; Vignols, F. Direct interaction between the Rice yellow mottle virus (RYMV) VPg and the central domain of the rice eIF(iso)4G1 factor correlates with rice susceptibility and RYMV virulence. Mol. Plant Microbe Interact. 2010, 23, 1506–1513. [Google Scholar] [CrossRef]
- Torrance, L.; Cowan, G.H.; McLean, K.; MacFarlane, S.; Al-Abedy, A.N.; Armstrong, M.; Lim, T.Y.; Hein, I.; Bryan, G.J. Natural resistance to Potato virus Y in Solanum tuberosum Group Phureja. Theor. Appl. Genet. 2020, 133, 967–980. [Google Scholar] [CrossRef]
- Jridi, C.; Martin, J.F.; Marie-Jeanne, V.; Labonne, G.; Blanc, S. Distinct viral populations differentiate and evolve independently in a single perennial host plant. J. Virol. 2006, 80, 2349–2357. [Google Scholar] [CrossRef]
- Simmonds, P.; Aiewsakun, P.; Katzourakis, A. Prisoners of war-host adaptation and its constraints on virus evolution. Nat. Rev. Microbiol. 2019, 17, 321–328. [Google Scholar] [CrossRef]
- Andika, I.B.; Wei, S.; Cao, C.; Salaipeth, L.; Kondo, H.; Sun, L. Phytopathogenic fungus hosts a plant virus: A naturally occurring cross-kingdom viral infection. Proc. Natl. Acad. Sci. USA 2017, 114, 12267–12272. [Google Scholar] [CrossRef] [Green Version]
- Elena, S.F. Local adaptation of plant viruses: Lessons from experimental evolution. Mol. Ecol. 2017, 26, 1711–1719. [Google Scholar] [CrossRef] [PubMed]
- Bedhomme, S.; Lafforgue, G.; Elena, S.F. Multihost experimental evolution of a plant RNA virus reveals local adaptation and host-specific mutations. Mol. Biol. Evol. 2012, 29, 1481–1492. [Google Scholar] [CrossRef] [PubMed]
- McLeish, M.J.; Fraile, A.; Garcia-Arenal, F. Evolution of plant-virus interactions: Host range and virus emergence. Curr. Opin. Virol. 2019, 34, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Perez, M.G.; Pagan, I.; Aragon-Caballero, L.; Caceres, F.; Fraile, A.; Garcia-Arenal, F. Ecological and genetic determinants of Pepino Mosaic Virus emergence. J. Virol. 2014, 88, 3359–3368. [Google Scholar] [CrossRef]
- Correa, R.L.; Sanz-Carbonell, A.; Kogej, Z.; Muller, S.Y.; Ambros, S.; Lopez-Gomollon, S.; Gomez, G.; Baulcombe, D.C.; Elena, S.F. Viral Fitness Determines the Magnitude of Transcriptomic and Epigenomic Reprograming of Defense Responses in Plants. Mol. Biol. Evol. 2020, 37, 1866–1881. [Google Scholar] [CrossRef]
- Martinez-Turino, S.; Calvo, M.; Bedoya, L.C.; Zhao, M.; Garcia, J.A. Virus Host Jumping Can Be Boosted by Adaptation to a Bridge Plant Species. Microorganisms 2021, 9, 805. [Google Scholar] [CrossRef]
- Novakova, S.; Subr, Z.; Kovac, A.; Fialova, I.; Beke, G.; Danchenko, M. Cucumber mosaic virus resistance: Comparative proteomics of contrasting Cucumis sativus cultivars after long-term infection. J. Proteom. 2020, 214, 103626. [Google Scholar] [CrossRef]
- Garcia-Ruiz, H.; Carbonell, A.; Hoyer, J.S.; Fahlgren, N.; Gilbert, K.B.; Takeda, A.; Giampetruzzi, A.; Garcia Ruiz, M.T.; McGinn, M.G.; Lowery, N.; et al. Roles and programming of Arabidopsis ARGONAUTE proteins during Turnip mosaic virus infection. PLoS Pathog. 2015, 11, e1004755. [Google Scholar] [CrossRef]
- Brosseau, C.; Bolaji, A.; Roussin-Leveillee, C.; Zhao, Z.; Biga, S.; Moffett, P. Natural variation in the Arabidopsis AGO2 gene is associated with susceptibility to potato virus X. New Phytol. 2020, 226, 866–878. [Google Scholar] [CrossRef]
- Dolja, V.V.; Krupovic, M.; Koonin, E.V. Deep Roots and Splendid Boughs of the Global Plant Virome. Annu. Rev. Phytopathol. 2020, 58, 23–53. [Google Scholar] [CrossRef]
- Power, A.G.; Flecker, A.S. 17. Virus Specificity in Disease Systems: Are Species Redundant? In The Importance of Species; Kareiva, P., Levin, S.A., Eds.; Princeton University Press: Princeton, NJ, USA, 2003; pp. 330–346. [Google Scholar]
- Blanc, S.; Michalakis, Y. Manipulation of hosts and vectors by plant viruses and impact of the environment. Curr. Opin. Insect. Sci. 2016, 16, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Power, A.G. Community Ecology of Plant Viruses. In Plant Virus Evolution; Roossinck, M.J., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 15–26. [Google Scholar]
- Mauck, K.E.; Chesnais, Q.; Shapiro, L.R. Chapter Seven—Evolutionary Determinants of Host and Vector Manipulation by Plant Viruses. In Advances in Virus Research; Malmstrom, C.M., Ed.; Environmental Virology and Virus Ecology; Academic Press: Cambridge, MA, USA, 2018; Volume 101, pp. 189–250. [Google Scholar]
- Lu, G.; Li, S.; Zhou, C.; Qian, X.; Xiang, Q.; Yang, T.; Wu, J.; Zhou, X.; Zhou, Y.; Ding, X.S.; et al. Tenuivirus utilizes its glycoprotein as a helper component to overcome insect midgut barriers for its circulative and propagative transmission. PLoS Pathog. 2019, 15, e1007655. [Google Scholar] [CrossRef] [PubMed]
- Lefeuvre, P.; Martin, D.P.; Elena, S.F.; Shepherd, D.N.; Roumagnac, P.; Varsani, A. Evolution and ecology of plant viruses. Nat. Rev. Microbiol. 2019, 17, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Wisler, G.C.; Duffus, J.E.; Liu, H.Y.; Li, R.H. Ecology and Epidemiology of Whitefly-Transmitted Closteroviruses. Plant Dis. 1998, 82, 270–280. [Google Scholar] [CrossRef]
- Peter, K.A.; Liang, D.; Palukaitis, P.; Gray, S.M. Small deletions in the potato leafroll virus readthrough protein affect particle morphology, aphid transmission, virus movement and accumulation. J. Gen. Virol. 2008, 89, 2037–2045. [Google Scholar] [CrossRef]
- Sin, S.H.; McNulty, B.C.; Kennedy, G.G.; Moyer, J.W. Viral genetic determinants for thrips transmission of Tomato spotted wilt virus. Proc. Natl. Acad. Sci. USA 2005, 102, 5168–5173. [Google Scholar] [CrossRef]
- Pan, L.L.; Chi, Y.; Liu, C.; Fan, Y.Y.; Liu, S.S. Mutations in the coat protein of a begomovirus result in altered transmission by different species of whitefly vectors. Virus Evol. 2020, 6, veaa014. [Google Scholar] [CrossRef]
- Ding, T.B.; Li, J.; Chen, E.H.; Niu, J.Z.; Chu, D. Transcriptome Profiling of the Whitefly Bemisia tabaci MED in Response to Single Infection of Tomato yellow leaf curl virus, Tomato chlorosis virus, and Their Co-infection. Front. Physiol. 2019, 10, 302. [Google Scholar] [CrossRef]
- Wang, Q.; Li, J.; Dang, C.; Chang, X.; Fang, Q.; Stanley, D.; Ye, G. Rice dwarf virus infection alters green rice leafhopper host preference and feeding behavior. PLoS ONE 2018, 13, e0203364. [Google Scholar] [CrossRef]
- Gadhave, K.R.; Dutta, B.; Coolong, T.; Srinivasan, R. A non-persistent aphid-transmitted Potyvirus differentially alters the vector and non-vector biology through host plant quality manipulation. Sci. Rep. 2019, 9, 2503. [Google Scholar] [CrossRef] [Green Version]
- Nachappa, P.; Challacombe, J.; Margolies, D.C.; Nechols, J.R.; Whitfield, A.E.; Rotenberg, D. Tomato Spotted Wilt Virus Benefits Its Thrips Vector by Modulating Metabolic and Plant Defense Pathways in Tomato. Front. Plant Sci. 2020, 11, 575564. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Delafuente, A.; Garzo, E.; Moreno, A.; Fereres, A. A plant virus manipulates the behavior of its whitefly vector to enhance its transmission efficiency and spread. PLoS ONE 2013, 8, e61543. [Google Scholar] [CrossRef] [PubMed]
- Adachi, S.; Honma, T.; Yasaka, R.; Ohshima, K.; Tokuda, M. Effects of infection by Turnip mosaic virus on the population growth of generalist and specialist aphid vectors on turnip plants. PLoS ONE 2018, 13, e0200784. [Google Scholar] [CrossRef] [PubMed]
- Fetters, A.M.; Cantalupo, P.G.; Wei, N.; Robles, M.T.S.; Stanley, A.; Stephens, J.D.; Pipas, J.M.; Ashman, T.L. The pollen virome of wild plants and its association with variation in floral traits and land use. Nat. Commun. 2022, 13, 523. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Nevado, C.; Gavilán, R.G.; Pagan, I. Host Abundance and Identity Determine the Epidemiology and Evolution of a Generalist Plant Virus in a Wild Ecosystem. Phytopathology 2020, 110, 94–105. [Google Scholar] [CrossRef]
- Lacroix, C.; Jolles, A.; Seabloom, E.W.; Power, A.G.; Mitchell, C.E.; Borer, E.T. Non-random biodiversity loss underlies predictable increases in viral disease prevalence. J. R. Soc. Interface 2014, 11, 20130947. [Google Scholar] [CrossRef]
- Liu, X.; Chen, L.F.; Liu, M.; Garcia-Guzman, G.; Gilbert, G.S.; Zhou, S.R. Dilution effect of plant diversity on infectious diseases: Latitudinal trend and biological context dependence. Oikos 2020, 129, 457–465. [Google Scholar] [CrossRef]
- Pagan, I.; Gonzalez-Jara, P.; Moreno-Letelier, A.; Rodelo-Urrego, M.; Fraile, A.; Pinero, D.; Garcia-Arenal, F. Effect of biodiversity changes in disease risk: Exploring disease emergence in a plant-virus system. PLoS Pathog. 2012, 8, e1002796. [Google Scholar] [CrossRef]
- Bernardo, P.; Charles-Dominique, T.; Barakat, M.; Ortet, P.; Fernandez, E.; Filloux, D.; Hartnady, P.; Rebelo, T.A.; Cousins, S.R.; Mesleard, F.; et al. Geometagenomics illuminates the impact of agriculture on the distribution and prevalence of plant viruses at the ecosystem scale. ISME J 2018, 12, 173–184. [Google Scholar] [CrossRef]
- Roossinck, M.J. The good viruses: Viral mutualistic symbioses. Nat. Rev. Microbiol. 2011, 9, 99–108. [Google Scholar] [CrossRef]
- Xu, P.; Chen, F.; Mannas, J.P.; Feldman, T.; Sumner, L.W.; Roossinck, M.J. Virus infection improves drought tolerance. New Phytol. 2008, 180, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Hily, J.M.; Poulicard, N.; Mora, M.A.; Pagan, I.; Garcia-Arenal, F. Environment and host genotype determine the outcome of a plant-virus interaction: From antagonism to mutualism. New Phytol. 2016, 209, 812–822. [Google Scholar] [CrossRef] [PubMed]
- Merleau, N.S.C.; Penisson, S.; Gerrish, P.J.; Elena, S.F.; Smerlak, M. Why are viral genomes so fragile? The bottleneck hypothesis. PLoS Comput Biol 2021, 17, e1009128. [Google Scholar] [CrossRef]
- Steinhauer, D.A.; Domingo, E.; Holland, J.J. Lack of evidence for proofreading mechanisms associated with an RNA virus polymerase. Gene 1992, 122, 281–288. [Google Scholar] [CrossRef]
- Tentchev, D.; Verdin, E.; Marchal, C.; Jacquet, M.; Aguilar, J.M.; Moury, B. Evolution and structure of Tomato spotted wilt virus populations: Evidence of extensive reassortment and insights into emergence processes. J. Gen. Virol. 2011, 92, 961–973. [Google Scholar] [CrossRef] [PubMed]
- Salem, N.; Mansour, A.; Ciuffo, M.; Falk, B.W.; Turina, M. A new tobamovirus infecting tomato crops in Jordan. Arch. Virol. 2016, 161, 503–506. [Google Scholar] [CrossRef]
- Garcia-Ruiz, H.; Diaz, A.; Ahlquist, P. Intermolecular RNA Recombination Occurs at Different Frequencies in Alternate Forms of Brome Mosaic Virus RNA Replication Compartments. Viruses 2018, 10, 131. [Google Scholar] [CrossRef]
- Schneider, W.L.; Roossinck, M.J. Genetic diversity in RNA virus quasispecies is controlled by host-virus interactions. J Virol 2001, 75, 6566–6571. [Google Scholar] [CrossRef]
- Schneider, W.L.; Roossinck, M.J. Evolutionarily related Sindbis-like plant viruses maintain different levels of population diversity in a common host. J Virol 2000, 74, 3130–3134. [Google Scholar] [CrossRef]
- Ali, A.; Roossinck, M.J. Genetic Bottlenecks. In Plant Virus Evolution; Roossinck, M.J., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 123–131. [Google Scholar]
- Garcia-Arenal, F.; Fraile, A.; Malpica, J.M. Variability and genetic structure of plant virus populations. Annu. Rev. Phytopathol. 2001, 39, 157–186. [Google Scholar] [CrossRef]
- Sun, B.J.; Sun, L.Y.; Tugume, A.K.; Adams, M.J.; Yang, J.; Xie, L.H.; Chen, J.P. Selection pressure and founder effects constrain genetic variation in differentiated populations of soilborne bymovirus Wheat yellow mosaic virus (Potyviridae) in China. Phytopathology 2013, 103, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Li, H.; Schneider, W.L.; Sherman, D.J.; Gray, S.; Smith, D.; Roossinck, M.J. Analysis of genetic bottlenecks during horizontal transmission of Cucumber mosaic virus. J. Virol. 2006, 80, 8345–8350. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Roossinck, M.J. Genetic bottlenecks reduce population variation in an experimental RNA virus population. J. Virol. 2004, 78, 10582–10587. [Google Scholar] [CrossRef] [PubMed]
- Paudel, D.B.; Sanfacon, H. Exploring the Diversity of Mechanisms Associated With Plant Tolerance to Virus Infection. Front. Plant Sci. 2018, 9, 1575. [Google Scholar] [CrossRef] [PubMed]
- Tepfer, M.; Girardot, G.; Feneant, L.; Ben Tamarzizt, H.; Verdin, E.; Moury, B.; Jacquemond, M. A genetically novel, narrow-host-range isolate of cucumber mosaic virus (CMV) from rosemary. Arch. Virol. 2016, 161, 2013–2017. [Google Scholar] [CrossRef]
- Shi, B.J.; Palukaitis, P.; Symons, R.H. Differential virulence by strains of Cucumber mosaic virus is mediated by the 2b gene. Mol. Plant Microbe Interact. 2002, 15, 947–955. [Google Scholar] [CrossRef]
- Sanjuan, R.; Domingo-Calap, P. Mechanisms of viral mutation. Cell Mol. Life Sci. 2016, 73, 4433–4448. [Google Scholar] [CrossRef]
- Duffy, S.; Shackelton, L.A.; Holmes, E.C. Rates of evolutionary change in viruses: Patterns and determinants. Nat. Rev. Genet. 2008, 9, 267–276. [Google Scholar] [CrossRef]
- Hicks, A.L.; Duffy, S. Cell tropism predicts long-term nucleotide substitution rates of mammalian RNA viruses. PLoS Pathog. 2014, 10, e1003838. [Google Scholar] [CrossRef]
- Schulte, M.B.; Draghi, J.A.; Plotkin, J.B.; Andino, R. Experimentally guided models reveal replication principles that shape the mutation distribution of RNA viruses. Elife 2015, 4, e03753. [Google Scholar] [CrossRef]
- Duffy, S.; Seah, Y.M. 98% identical, 100% wrong: Per cent nucleotide identity can lead plant virus epidemiology astray. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 1891–1897. [Google Scholar] [CrossRef]
- Calvo, M.; Malinowski, T.; Garcia, J.A. Single amino acid changes in the 6K1-CI region can promote the alternative adaptation of Prunus- and Nicotiana-propagated Plum pox virus C isolates to either host. Mol. Plant Microbe Interact. 2014, 27, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Vassilakos, N.; Simon, V.; Tzima, A.; Johansen, E.; Moury, B. Genetic Determinism and Evolutionary Reconstruction of a Host Jump in a Plant Virus. Mol. Biol. Evol. 2016, 33, 541–553. [Google Scholar] [CrossRef]
- Simmonds, P. Methods for virus classification and the challenge of incorporating metagenomic sequence data. J. Gen. Virol. 2015, 96, 1193–1206. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Pendon, J.A.; Sanchez-Campos, S.; Fortes, I.M.; Moriones, E. Tomato Yellow Leaf Curl Sardinia Virus, a Begomovirus Species Evolving by Mutation and Recombination: A Challenge for Virus Control. Viruses 2019, 11, 45. [Google Scholar] [CrossRef] [PubMed]
- Eggenberger, A.L.; Hajimorad, M.R.; Hill, J.H. Gain of virulence on Rsv1-genotype soybean by an avirulent Soybean mosaic virus requires concurrent mutations in both P3 and HC-Pro. Mol. Plant Microbe Interact. 2008, 21, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Khatabi, B.; Fajolu, O.L.; Wen, R.H.; Hajimorad, M.R. Evaluation of North American isolates of Soybean mosaic virus for gain of virulence on Rsv-genotype soybeans with special emphasis on resistance-breaking determinants on Rsv4. Mol. Plant Pathol. 2012, 13, 1077–1088. [Google Scholar] [CrossRef]
- Guiu-Aragones, C.; Diaz-Pendon, J.A.; Martin-Hernandez, A.M. Four sequence positions of the movement protein of Cucumber mosaic virus determine the virulence against cmv1-mediated resistance in melon. Mol. Plant Pathol. 2015, 16, 675–684. [Google Scholar] [CrossRef]
- Wieczorek, P.; Obrepalska-Steplowska, A. A single amino acid substitution in movement protein of tomato torrado virus influences ToTV infectivity in Solanum lycopersicum. Virus. Res. 2016, 213, 32–36. [Google Scholar] [CrossRef]
- Xu, J.; Wang, X.; Shi, L.; Zhou, Y.; Li, D.; Han, C.; Zhang, Z.; Yu, J. Two distinct sites are essential for virulent infection and support of variant satellite RNA replication in spontaneous beet black scorch virus variants. J. Gen. Virol. 2012, 93, 2718–2728. [Google Scholar] [CrossRef] [Green Version]
- Bertens, P.; Wellink, J.; Goldbach, R.; van Kammen, A. Mutational analysis of the cowpea mosaic virus movement protein. Virology 2000, 267, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Obbard, D.J.; Jiggins, F.M.; Halligan, D.L.; Little, T.J. Natural selection drives extremely rapid evolution in antiviral RNAi genes. Curr. Biol. 2006, 16, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.C.; Chiang, C.H.; Raja, J.A.; Liu, F.L.; Tai, C.H.; Yeh, S.D. A single amino acid of niapro of papaya ringspot virus determines host specificity for infection of papaya. Mol. Plant Microbe Interact. 2008, 21, 1046–1057. [Google Scholar] [CrossRef] [PubMed]
- Azzam, O.; Arboleda, M.; Umadhay, K.M.; de los Reyes, J.B.; Cruz, F.S.; Mackenzie, A.; McNally, K.L. Genetic composition and complexity of virus populations at tungro-endemic and outbreak rice sites. Arch. Virol. 2000, 145, 2643–2657. [Google Scholar] [CrossRef]
- Simon-Loriere, E.; Holmes, E.C. Why do RNA viruses recombine? Nat. Rev. Microbiol. 2011, 9, 617–626. [Google Scholar] [CrossRef]
- Yuksel Ozmen, C.; Khabbazi, S.D.; Khabbazi, A.D.; Gurel, S.; Kaya, R.; Oguz, M.C.; Turan, F.; Rezaei, F.; Kibar, U.; Gurel, E.; et al. Genome composition analysis of multipartite BNYVV reveals the occurrence of genetic re-assortment in the isolates of Asia Minor and Thrace. Sci. Rep. 2020, 10, 4129. [Google Scholar] [CrossRef]
- Codoner, F.M.; Elena, S.F. The promiscuous evolutionary history of the family Bromoviridae. J. Gen. Virol. 2008, 89, 1739–1747. [Google Scholar] [CrossRef]
- Lee, Y.J.; Kil, E.J.; Kwak, H.R.; Kim, M.; Seo, J.K.; Lee, S.; Choi, H.S. Phylogenetic Characterization of Tomato chlorosis virus Population in Korea: Evidence of Reassortment between Isolates from Different Origins. Plant Pathol. J. 2018, 34, 199–207. [Google Scholar] [CrossRef]
- Ohki, T.; Sasaya, T.; Maoka, T. Cylindrical Inclusion Protein of Wheat Yellow Mosaic Virus Is Involved in Differential Infection of Wheat Cultivars. Phytopathology 2019, 109, 1475–1480. [Google Scholar] [CrossRef]
- Gu, H.; Clark, A.J.; de Sa, P.B.; Pfeiffer, T.W.; Tolin, S.; Ghabrial, S.A. Diversity Among Isolates of Bean pod mottle virus. Phytopathology 2002, 92, 446–452. [Google Scholar] [CrossRef]
- Kalyandurg, P.; Gil, J.F.; Lukhovitskaya, N.I.; Flores, B.; Muller, G.; Chuquillanqui, C.; Palomino, L.; Monjane, A.; Barker, I.; Kreuze, J.; et al. Molecular and pathobiological characterization of 61 Potato mop-top virus full-length cDNAs reveals great variability of the virus in the centre of potato domestication, novel genotypes and evidence for recombination. Mol. Plant Pathol. 2017, 18, 864–877. [Google Scholar] [CrossRef] [PubMed]
- Thekke-Veetil, T.; Polashock, J.J.; Marn, M.V.; Plesko, I.M.; Schilder, A.C.; Keller, K.E.; Martin, R.R.; Tzanetakis, I.E. Population structure of blueberry mosaic associated virus: Evidence of reassortment in geographically distinct isolates. Virus. Res. 2015, 201, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Patil, B.L.; Dangwal, M.; Mishra, R. Variability of Emaravirus Species Associated with Sterility Mosaic Disease of Pigeonpea in India Provides Evidence of Segment Reassortment. Viruses 2017, 9, 183. [Google Scholar] [CrossRef] [PubMed]
- Bolus, S.; Braithwaite, K.S.; Grinstead, S.C.; Fuentes-Bueno, I.; Beiriger, R.; Falk, B.W.; Mollov, D. Completion of Maize Stripe Virus Genome Sequence and Analysis of Diverse Isolates. Front. Microbiol. 2021, 12, 684599. [Google Scholar] [CrossRef]
- Roy, A.; Stone, A.L.; Otero-Colina, G.; Wei, G.; Brlansky, R.H.; Ochoa, R.; Bauchan, G.; Schneider, W.L.; Nakhla, M.K.; Hartung, J.S. Reassortment of Genome Segments Creates Stable Lineages Among Strains of Orchid Fleck Virus Infecting Citrus in Mexico. Phytopathology 2020, 110, 106–120. [Google Scholar] [CrossRef]
- Kwon, S.J.; Cho, Y.E.; Kwon, O.H.; Kang, H.G.; Seo, J.K. Resistance-Breaking Tomato Spotted Wilt Virus Variant that Recently Occurred in Pepper in South Korea is a Genetic Reassortant. Plant Dis. 2021, 105, 2771–2775. [Google Scholar] [CrossRef]
- Petrzik, K. Evolutionary forces at work in partitiviruses. Virus Genes 2019, 55, 563–573. [Google Scholar] [CrossRef]
- Yin, X.; Zheng, F.Q.; Tang, W.; Zhu, Q.Q.; Li, X.D.; Zhang, G.M.; Liu, H.T.; Liu, B.S. Genetic structure of rice black-streaked dwarf virus populations in China. Arch. Virol. 2013, 158, 2505–2515. [Google Scholar] [CrossRef]
- Savory, F.R.; Ramakrishnan, U. Asymmetric patterns of reassortment and concerted evolution in Cardamom bushy dwarf virus. Infect. Genet. Evol. 2014, 24, 15–24. [Google Scholar] [CrossRef]
- Chen, L.F.; Rojas, M.; Kon, T.; Gamby, K.; Xoconostle-Cazares, B.; Gilbertson, R.L. A severe symptom phenotype in tomato in Mali is caused by a reassortant between a novel recombinant begomovirus (Tomato yellow leaf curl Mali virus) and a betasatellite. Mol. Plant Pathol. 2009, 10, 415–430. [Google Scholar] [CrossRef]
- Briese, T.; Calisher, C.H.; Higgs, S. Viruses of the family Bunyaviridae: Are all available isolates reassortants? Virology 2013, 446, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Rosa, C. Thrips as the Transmission Bottleneck for Mixed Infection of Two Orthotospoviruses. Plants 2020, 9, 509. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, S.; Michalakis, Y.; Blanc, S. Virus population bottlenecks during within-host progression and host-to-host transmission. Curr. Opin. Virol. 2012, 2, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Chao, L. Levels of selection, evolution of sex in RNA viruses, and the origin of life. J. Theor. Biol. 1991, 153, 229–246. [Google Scholar] [CrossRef]
- Sicard, A.; Yvon, M.; Timchenko, T.; Gronenborn, B.; Michalakis, Y.; Gutierrez, S.; Blanc, S. Gene copy number is differentially regulated in a multipartite virus. Nat. Commun. 2013, 4, 2248. [Google Scholar] [CrossRef]
- Wu, B.; Zwart, M.P.; Sanchez-Navarro, J.A.; Elena, S.F. Within-host Evolution of Segments Ratio for the Tripartite Genome of Alfalfa Mosaic Virus. Sci. Rep. 2017, 7, 5004. [Google Scholar] [CrossRef]
- Lowen, A.C. It’s in the mix: Reassortment of segmented viral genomes. PLoS Pathog. 2018, 14, e1007200. [Google Scholar] [CrossRef]
- Miyashita, S.; Kishino, H. Estimation of the size of genetic bottlenecks in cell-to-cell movement of soil-borne wheat mosaic virus and the possible role of the bottlenecks in speeding up selection of variations in trans-acting genes or elements. J. Virol. 2010, 84, 1828–1837. [Google Scholar] [CrossRef]
- Escriu, F.; Fraile, A.; Garcia-Arenal, F. Constraints to genetic exchange support gene coadaptation in a tripartite RNA virus. PLoS Pathog. 2007, 3, e8. [Google Scholar] [CrossRef]
- White, M.C.; Lowen, A.C. Implications of segment mismatch for influenza A virus evolution. J. Gen. Virol. 2018, 99, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Fraile, A.; Alonso-Prados, J.L.; Aranda, M.A.; Bernal, J.J.; Malpica, J.M.; Garcia-Arenal, F. Genetic exchange by recombination or reassortment is infrequent in natural populations of a tripartite RNA plant virus. J. Virol. 1997, 71, 934–940. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Arenal, F.; Fraile, A.; Malpica, J.M. Variation and evolution of plant virus populations. Int. Microbiol. 2003, 6, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Arenal, F.; McDonald, B.A. An analysis of the durability of resistance to plant viruses. Phytopathology 2003, 93, 941–952. [Google Scholar] [CrossRef]
- Adegbola, R.O.; Marshall, S.H.; Batuman, O.; Ullman, D.E.; Gilbertson, R.L.; Adkins, S.; Naidu, R.A. Sequence analysis of the medium and small RNAs of impatiens necrotic spot virus reveals segment reassortment but not recombination. Arch. Virol. 2019, 164, 2829–2836. [Google Scholar] [CrossRef]
- Roossinck, M.J. Evolutionary history of Cucumber mosaic virus deduced by phylogenetic analyses. J. Virol. 2002, 76, 3382–3387. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Hirota, K.; Maruyama, K.; Andika, I.B.; Suzuki, N. A possible occurrence of genome reassortment among bipartite rhabdoviruses. Virology 2017, 508, 18–25. [Google Scholar] [CrossRef]
- Grigoras, I.; Ginzo, A.; Martin, D.P.; Varsani, A.; Romero, J.; Mammadov, A.C.; Huseynova, I.M.; Aliyev, J.A.; Kheyr-Pour, A.; Huss, H.; et al. Genome diversity and evidence of recombination and reassortment in nanoviruses from Europe. J. Gen. Virol. 2014, 95, 1178–1191. [Google Scholar] [CrossRef]
- Gu, H.; Zhang, C.; Ghabrial, S.A. Novel Naturally Occurring Bean pod mottle virus Reassortants with Mixed Heterologous RNA1 Genomes. Phytopathology 2007, 97, 79–86. [Google Scholar] [CrossRef]
- Pita, J.S.; Fondong, V.N.; Sangare, A.; Otim-Nape, G.W.; Ogwal, S.; Fauquet, C.M. Recombination, pseudorecombination and synergism of geminiviruses are determinant keys to the epidemic of severe cassava mosaic disease in Uganda. J. Gen. Virol. 2001, 82, 655–665. [Google Scholar] [CrossRef]
- Kim, Y.I.; Si, Y.J.; Kwon, H.I.; Kim, E.H.; Park, S.J.; Robles, N.J.; Nguyen, H.D.; Yu, M.A.; Yu, K.M.; Lee, Y.J.; et al. Pathogenicity and genetic characterisation of a novel reassortant, highly pathogenic avian influenza (HPAI) H5N6 virus isolated in Korea, 2017. Euro Surveill. 2018, 23, 18–00045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Höfer, P.; Engel, M.; Jeske, H.; Frischmuth, T. Host Range Limitation of a Pseudorecombinant Virus Produced by Two Distinct Bipartite Geminiviruses. Mol. Plant Microbe Interact. 1997, 10, 1019–1022. [Google Scholar] [CrossRef]
- Idris, A.M.; Mills-Lujan, K.; Martin, K.; Brown, J.K. Melon chlorotic leaf curl virus: Characterization and differential reassortment with closest relatives reveal adaptive virulence in the squash leaf curl virus clade and host shifting by the host-restricted bean calico mosaic virus. J. Virol. 2008, 82, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Bergua, M.; Luis-Arteaga, M.; Escriu, F. Genetic Diversity, Reassortment, and Recombination in Alfalfa mosaic virus Population in Spain. Phytopathology 2014, 104, 1241–1250. [Google Scholar] [CrossRef]
- Lian, S.; Lee, J.S.; Cho, W.K.; Yu, J.; Kim, M.K.; Choi, H.S.; Kim, K.H. Phylogenetic and recombination analysis of tomato spotted wilt virus. PLoS ONE 2013, 8, e63380. [Google Scholar] [CrossRef]
- Rodelo-Urrego, M.; Garcia-Arenal, F.; Pagan, I. The effect of ecosystem biodiversity on virus genetic diversity depends on virus species: A study of chiltepin-infecting begomoviruses in Mexico. Virus Evol. 2015, 1, vev004. [Google Scholar] [CrossRef]
- Kraberger, S.; Kumari, S.G.; Najar, A.; Stainton, D.; Martin, D.P.; Varsani, A. Molecular characterization of faba bean necrotic yellows viruses in Tunisia. Arch. Virol. 2018, 163, 687–694. [Google Scholar] [CrossRef]
- Ferriol, I.; Ferrer, R.M.; Luis-Arteaga, M.; Guerri, J.; Moreno, P.; Rubio, L. Genetic variability and evolution of broad bean wilt virus 1: Role of recombination, selection and gene flow. Arch. Virol. 2014, 159, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Kwak, H.R.; Kim, M.K.; Lee, Y.J.; Seo, J.K.; Kim, J.S.; Kim, K.H.; Cha, B.; Choi, H.S. Molecular Characterization and Variation of the Broad bean wilt virus 2 Isolates Based on Analyses of Complete Genome Sequences. Plant Pathol. J. 2013, 29, 397–409. [Google Scholar] [CrossRef]
- Galvino-Costa, S.B.F.; Figueira, A.D.; Camargos, V.V.; Geraldino, P.S.; Hu, X.J.; Nikolaeva, O.V.; Kerlan, C.; Karasev, A.V. A novel type of Potato virus Y recombinant genome, determined for the genetic strain PVYE. Plant Pathol. 2012, 61, 388–398. [Google Scholar] [CrossRef]
- Chare, E.R.; Holmes, E.C. A phylogenetic survey of recombination frequency in plant RNA viruses. Arch. Virol. 2006, 151, 933–946. [Google Scholar] [CrossRef]
- Nigam, D.; LaTourrette, K.; Garcia-Ruiz, H. Mutations in virus-derived small RNAs. Sci. Rep. 2020, 10, 9540. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, H.; Ahlquist, P. Inducible yeast system for Viral RNA recombination reveals requirement for an RNA replication signal on both parental RNAs. J. Virol. 2006, 80, 8316–8328. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Andres, S.; Monci, F.; Navas-Castillo, J.; Moriones, E. Begomovirus genetic diversity in the native plant reservoir Solanum nigrum: Evidence for the presence of a new virus species of recombinant nature. Virology 2006, 350, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Lefeuvre, P.; Moriones, E. Recombination as a motor of host switches and virus emergence: Geminiviruses as case studies. Curr. Opin. Virol. 2015, 10, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Heinze, C.; Wobbe, V.; Lesemann, D.E.; Zhang, D.Y.; Willingmann, P.; Adam, G. Pelargonium necrotic spot virus: A new member of the genus Tombusvirus. Arch. Virol. 2004, 149, 1527–1539. [Google Scholar] [CrossRef]
- Hasiów-Jaroszewska, B.; Kuzniar, A.; Peters, S.A.; Leunissen, J.A.M.; Pospieszny, H. Evidence for RNA recombination between distinct isolates of Pepino mosaic virus. Acta Biochim. Pol. 2010, 57, 385–388. [Google Scholar] [CrossRef]
- Bagayoko, I.; Celli, M.G.; Romay, G.; Poulicard, N.; Pinel-Galzi, A.; Julian, C.; Filloux, D.; Roumagnac, P.; Sereme, D.; Bragard, C.; et al. Genetic Diversity of Rice stripe necrosis virus and New Insights into Evolution of the Genus Benyvirus. Viruses 2021, 13, 737. [Google Scholar] [CrossRef]
- Veerakone, S.; Liefting, L.W.; Khan, S.; Pal, C.; Tang, J.; Ward, L.I. Partial biological and molecular characterization of a novel citrivirus from Nandina domestica. Arch. Virol. 2021, 166, 1395–1399. [Google Scholar] [CrossRef]
- Jacquemond, M. Chapter 13—Cucumber Mosaic Virus. In Advances in Virus Research; Loebenstein, G., Lecoq, H., Eds.; Viruses and Virus Diseases of Vegetables in the Mediterranean Basin; Academic Press: Cambridge, MA, USA, 2012; Volume 84, pp. 439–504. [Google Scholar]
- Harper, S.J.; Dawson, T.E.; Pearson, M.N. Isolates of Citrus tristeza virus that overcome Poncirus trifoliata resistance comprise a novel strain. Arch. Virol. 2010, 155, 471–480. [Google Scholar] [CrossRef]
- Desbiez, C.; Wipf-Scheibel, C.; Millot, P.; Verdin, E.; Dafalla, G.; Lecoq, H. New species in the papaya ringspot virus cluster: Insights into the evolution of the PRSV lineage. Virus. Res. 2017, 241, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Oliver, J.E.; Vigne, E.; Fuchs, M. Genetic structure and molecular variability of Grapevine fanleaf virus populations. Virus. Res. 2010, 152, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Moonan, F.; Molina, J.; Mirkov, T.E. Sugarcane yellow leaf virus: An emerging virus that has evolved by recombination between luteoviral and poleroviral ancestors. Virology 2000, 269, 156–171. [Google Scholar] [CrossRef] [PubMed]
- Tzanetakis, I.E.; Tsai, C.H.; Martin, R.R.; Dreher, T.W. A tymovirus with an atypical 3′-UTR illuminates the possibilities for 3′-UTR evolution. Virology 2009, 392, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Chavan, R.R.; Pearson, M.N. Molecular characterisation of a novel recombinant Ribgrass mosaic virus strain FSHS. Virol. J. 2016, 13, 29. [Google Scholar] [CrossRef]
- He, M.; Guan, S.Y.; He, C.Q. Evolution of rice stripe virus. Mol. Phylogenet. Evol. 2017, 109, 343–350. [Google Scholar] [CrossRef]
- Pappi, P.G.; Maliogka, V.I.; Amoutzias, G.D.; Katis, N.I. Genetic variation of eggplant mottled dwarf virus from annual and perennial plant hosts. Arch. Virol. 2016, 161, 631–639. [Google Scholar] [CrossRef]
- Iftikhar, R.; Ramesh, S.V.; Bag, S.; Ashfaq, M.; Pappu, H.R. Global analysis of population structure, spatial and temporal dynamics of genetic diversity, and evolutionary lineages of Iris yellow spot virus (Tospovirus: Bunyaviridae). Gene 2014, 547, 111–118. [Google Scholar] [CrossRef]
- Li, Y.; Xia, Z.; Peng, J.; Zhou, T.; Fan, Z. Evidence of recombination and genetic diversity in southern rice black-streaked dwarf virus. Arch. Virol. 2013, 158, 2147–2151. [Google Scholar] [CrossRef]
- Hyder, M.Z.; Shah, S.H.; Hameed, S.; Naqvi, S.M. Evidence of recombination in the Banana bunchy top virus genome. Infect. Genet. Evol. 2011, 11, 1293–1300. [Google Scholar] [CrossRef]
- Scussel, S.; Claverie, S.; Hoareau, M.; Moustache, R.; Delatte, H.; Lefeuvre, P.; Lett, J.M. Tomato leaf curl Mahe virus: A novel tomato-infecting monopartite begomovirus from the Seychelles. Arch. Virol. 2018, 163, 3451–3453. [Google Scholar] [CrossRef]
- Yasaka, R.; Nguyen, H.D.; Ho, S.Y.; Duchene, S.; Korkmaz, S.; Katis, N.; Takahashi, H.; Gibbs, A.J.; Ohshima, K. The temporal evolution and global spread of Cauliflower mosaic virus, a plant pararetrovirus. PLoS ONE 2014, 9, e85641. [Google Scholar] [CrossRef] [PubMed]
- Kraberger, S.; Kumari, S.G.; Hamed, A.A.; Gronenborn, B.; Thomas, J.E.; Sharman, M.; Harkins, G.W.; Muhire, B.M.; Martin, D.P.; Varsani, A. Molecular diversity of Chickpea chlorotic dwarf virus in Sudan: High rates of intra-species recombination—A driving force in the emergence of new strains. Infect. Genet. Evol. 2015, 29, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, L.; Simon, A.; Garcia, C.; Velasco, L.; Janssen, D. First natural crossover recombination between two distinct species of the family Closteroviridae leads to the emergence of a new disease. PLoS ONE 2018, 13, e0198228. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lin, J.; Zheng, G.; Zhang, M.; Zhi, H. Recombinant soybean mosaic virus is prevalent in Chinese soybean fields. Arch. Virol. 2014, 159, 1793–1796. [Google Scholar] [CrossRef]
- Silva, T.F.; Correa, R.L.; Castilho, Y.; Silvie, P.; Belot, J.L.; Vaslin, M.F. Widespread distribution and a new recombinant species of Brazilian virus associated with cotton blue disease. Virol. J. 2008, 5, 123. [Google Scholar] [CrossRef]
- Miras, M.; Sempere, R.N.; Kraft, J.J.; Miller, W.A.; Aranda, M.A.; Truniger, V. Interfamilial recombination between viruses led to acquisition of a novel translation-enhancing RNA element that allows resistance breaking. New Phytol. 2014, 202, 233–246. [Google Scholar] [CrossRef]
- Legg, J.P.; Thresh, J.M. Cassava mosaic virus disease in East Africa: A dynamic disease in a changing environment. Virus. Res. 2000, 71, 135–149. [Google Scholar] [CrossRef]
- Green, K.J.; Funke, C.N.; Chojnacky, J.; Alvarez-Quinto, R.A.; Ochoa, J.B.; Quito-Avila, D.F.; Karasev, A.V. Potato Virus Y (PVY) Isolates from Solanum betaceum Represent Three Novel Recombinants Within the PVY(N) Strain Group and Are Unable to Systemically Spread in Potato. Phytopathology 2020, 110, 1588–1596. [Google Scholar] [CrossRef]
- Ouedraogo, R.S.; Pita, J.S.; Somda, I.P.; Traore, O.; Roossinck, M.J. Impact of Cultivated Hosts on the Recombination of Cucumber Mosaic Virus. J. Virol. 2019, 93, e01770-18. [Google Scholar] [CrossRef]
- Hou, Y.M.; Gilbertson, R.L. Increased pathogenicity in a pseudorecombinant bipartite geminivirus correlates with intermolecular recombination. J. Virol. 1996, 70, 5430–5436. [Google Scholar] [CrossRef] [Green Version]
- Monci, F.; Sanchez-Campos, S.; Navas-Castillo, J.; Moriones, E. A natural recombinant between the geminiviruses Tomato yellow leaf curl Sardinia virus and Tomato yellow leaf curl virus exhibits a novel pathogenic phenotype and is becoming prevalent in Spanish populations. Virology 2002, 303, 317–326. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, A.S.; Melo, F.L.; Inoue-Nagata, A.K.; Nagata, T.; Kitajima, E.W.; Resende, R.O. Characterization of bean necrotic mosaic virus: A member of a novel evolutionary lineage within the Genus Tospovirus. PLoS ONE 2012, 7, e38634. [Google Scholar] [CrossRef] [PubMed]
- Braidwood, L.; Quito-Avila, D.F.; Cabanas, D.; Bressan, A.; Wangai, A.; Baulcombe, D.C. Maize chlorotic mottle virus exhibits low divergence between differentiated regional sub-populations. Sci. Rep. 2018, 8, 1173. [Google Scholar] [CrossRef]
- Komorowska, B.; Hasiow-Jaroszewska, B.; Elena, S.F. Evolving by deleting: Patterns of molecular evolution of Apple stem pitting virus isolates from Poland. J. Gen. Virol. 2019, 100, 1442–1456. [Google Scholar] [CrossRef]
- Hak, H.; Spiegelman, Z. The Tomato Brown Rugose Fruit Virus Movement Protein Overcomes Tm-2(2) Resistance in Tomato While Attenuating Viral Transport. Mol. Plant Microbe Interact. 2021, 34, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.Y.; Ma, H.Y.; Wang, L.; Tettey, C.; Zhao, M.S.; Geng, C.; Tian, Y.P.; Li, X.D. Identification of genetic determinants of tomato brown rugose fruit virus that enable infection of plants harbouring the Tm-2(2) resistance gene. Mol. Plant Pathol. 2021, 22, 1347–1357. [Google Scholar] [CrossRef]
- Gorbalenya, A.E.; Siddell, S.G. Recognizing species as a new focus of virus research. PLoS Pathog. 2021, 17, e1009318. [Google Scholar] [CrossRef]
- Dutilh, B.E.; Varsani, A.; Tong, Y.; Simmonds, P.; Sabanadzovic, S.; Rubino, L.; Roux, S.; Munoz, A.R.; Lood, C.; Lefkowitz, E.J.; et al. Perspective on taxonomic classification of uncultivated viruses. Curr. Opin. Virol. 2021, 51, 207–215. [Google Scholar] [CrossRef]
- Rumbou, A.; Vainio, E.J.; Buttner, C. Towards the Forest Virome: High-Throughput Sequencing Drastically Expands Our Understanding on Virosphere in Temperate Forest Ecosystems. Microorganisms 2021, 9, 1730. [Google Scholar] [CrossRef]
- Maclot, F.; Candresse, T.; Filloux, D.; Malmstrom, C.M.; Roumagnac, P.; van der Vlugt, R.; Massart, S. Illuminating an Ecological Blackbox: Using High Throughput Sequencing to Characterize the Plant Virome Across Scales. Front. Microbiol. 2020, 11, 578064. [Google Scholar] [CrossRef]
- Desbiez, C.; Joannon, B.; Wipf-Scheibel, C.; Chandeysson, C.; Lecoq, H. Recombination in natural populations of watermelon mosaic virus: New agronomic threat or damp squib? J. Gen. Virol. 2011, 92, 1939–1948. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Family | Genus | Species | Reassortment Effect | Reference |
|---|---|---|---|---|
| Benyviridae | Benyvirus | Beet necrotic yellow vein virus | Impact phylogenetic history | [117] |
| Bromoviridae | Bromovirus, Cucumovirus, Ilarvirus, and Oleavirus | 18 species | Impact phylogenetic history | [118] |
| Closteroviridae | Crinivirus | Tomato chlorosis virus | Impact phylogenetic history | [119] |
| Potyviridae | Bymovirus | Wheat yellow mosaic virus | Pathotype differentiation | [120] |
| Secoviridae | Comovirus | Bean pod mottle virus | Severe symptoms | [121] |
| Virgaviridae | Pomovirus | Potato mop-top virus | Impact phylogenetic history | [122] |
| Aspiviridae | Ophiovirus | Blueberry mosaic associated ophiovirus* | Impact phylogenetic history | [123] |
| Fimoviridae | Emaravirus | Pigeonpea sterility mosaic emaravirus 1* and Pigeonpea sterility mosaic emaravirus 2* | Impact phylogenetic history | [124] |
| Phenuiviridae | Tenuivirus | Maize stripe virus | Impact phylogenetic history | [125] |
| Rhabdoviridae | Dichorhavirus | Orchid fleck dichorhavirus* | New strain | [126] |
| Tospoviridae | Orthotospovirus | Tomato spotted wilt orthotospovirus* | Resistance-breaking | [127] |
| Partitiviridae | Alphapartitivirus, Betapartitivirus, Cryspovirus Deltapartitivirus, and Gammapartitivirus | 12 species | Impact phylogenetic history | [128] |
| Reoviridae | Fijivirus | Rice black-streaked dwarf virus | Impact phylogenetic history | [129] |
| Nanoviridae | Babuvirus | Cardamom bushy dwarf virus | Impact phylogenetic history | [130] |
| Geminiviridae | Begomovirus | Tomato yellow leaf curl Mali virus and Cotton leaf curl Gezira betasatellite | Severe symptoms | [131] |
| Family | Species | Reassortment Frequency | Recombination Frequency | Reference |
|---|---|---|---|---|
| Bromoviridae | Alfalfa mosaic virus | 2/60 = 3.33% | 1/60 = 1.67% | [154] |
| Cucumber mosaic virus | 4% | 7% | [142] | |
| Tospoviridae | Tomato spotted wilt orthotospovirus * | 3/13 = 23.08% | 2/13 = 15.38% | [155] |
| Impatiens necrotic spot orthotospovirus * | 6/18 = 33.33% | 0/18 = 0% | [145] | |
| Geminiviridae | Pepper golden mosaic virus | 20/47 = 42.55% | 27/47 = 57.45% | [156] |
| Pepper huasteco yellow vein virus | 11/42 = 26.19% | 13/42= 31.00% | ||
| Nanoviridae | Faba bean necrotic yellows virus | 6/18 = 33.33% | 13/18 = 72.22% | [157] |
| Secoviridae | Broad bean wilt virus 1 | 6/37 = 16.22% | 2/37 = 5.41% | [158] |
| Broad bean wilt virus 2 | 16/29 = 55.17% | 2/29 = 6.70% | [159] |
| Family | Genus | Species | Recombination Effect | Reference |
|---|---|---|---|---|
| Alphaflexiviridae | Potexvirus | Pepino mosaic virus | Impact phylogenetic history | [167] |
| Benyviridae | Benyvirus | Rice stripe necrosis virus | Impact phylogenetic history | [168] |
| Betaflexiviridae | Citrivirus | Citrus leaf blotch virus-Rec | New species | [169] |
| Bromoviridae | Cucumovirus | Cucumber mosaic virus | Impact phylogenetic history | [170] |
| Closteroviridae | Closterovirus | Citrus tristeza virus-RB | Resistance-breaking | [171] |
| Potyviridae | Potyvirus | Sudan watermelon mosaic virus | New species | [172] |
| Secoviridae | Nepovirus | Grapevine fanleaf virus | Impact phylogenetic history | [173] |
| Solemoviridae | Polerovirus | Sugarcane yellow leaf virus | New species | [174] |
| Tombusviridae | Tombusvirus | Pelargonium necrotic spot virus | New species | [166] |
| Tymoviridae | Tymovirus | Dulcamara mottle virus | New species | [175] |
| Virgaviridae | Tobamovirus | Ribgrass mosaic virus strain FSHS | New strain | [176] |
| Aspiviridae | NA | NA | NA | NA |
| Fimoviridae | Emaravirus | Pigeonpea sterility mosaic emaravirus 1 and Pigeonpea sterility mosaic emaravirus 2 | Impact phylogenetic history | [124] |
| Phenuiviridae | Tenuivirus | Rice stripe tenuivirus * | Impact phylogenetic history | [177] |
| Rhabdoviridae | Alphanucleorhabdovirus | Eggplant mottled dwarf alphanucleorhabdovirus | Impact phylogenetic history | [178] |
| Tospoviridae | Orthotospovirus | Iris yellow spot orthotospovirus * | Genotype separation | [179] |
| Endornaviridae | NA | NA | NA | NA |
| Partitiviridae | NA | NA | NA | NA |
| Pseudoviridae | NA | NA | NA | NA |
| Reoviridae | Fijivirus | Southern rice black-streaked dwarf virus | Impact phylogenetic history | [180] |
| Nanoviridae | Babuvirus | Banana bunchy top virus | Impact phylogenetic history | [181] |
| Geminiviridae | Begomovirus | Tomato leaf curl Mahé virus | New species | [182] |
| Caulimoviridae | Caulimovirus | Cauliflower mosaic virus | Impact phylogenetic history | [183] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
LaTourrette, K.; Garcia-Ruiz, H. Determinants of Virus Variation, Evolution, and Host Adaptation. Pathogens 2022, 11, 1039. https://doi.org/10.3390/pathogens11091039
LaTourrette K, Garcia-Ruiz H. Determinants of Virus Variation, Evolution, and Host Adaptation. Pathogens. 2022; 11(9):1039. https://doi.org/10.3390/pathogens11091039
Chicago/Turabian StyleLaTourrette, Katherine, and Hernan Garcia-Ruiz. 2022. "Determinants of Virus Variation, Evolution, and Host Adaptation" Pathogens 11, no. 9: 1039. https://doi.org/10.3390/pathogens11091039
APA StyleLaTourrette, K., & Garcia-Ruiz, H. (2022). Determinants of Virus Variation, Evolution, and Host Adaptation. Pathogens, 11(9), 1039. https://doi.org/10.3390/pathogens11091039