
Respiratory Commensal Bacteria Increase Protection against Hypermucoviscous Carbapenem-Resistant Klebsiella pneumoniae ST25 Infection

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Murine Infection Model
2.3. Respiratory Infection with K. pneumoniae LABACER 01 and LABACER 27
2.4. K. pneumoniae Counts in Lungs
2.5. Lung Damage
2.6. Leukocytes Counts
2.7. Serum Cytokines and Bronchoalveolar Lavages
2.8. Statistical Analysis
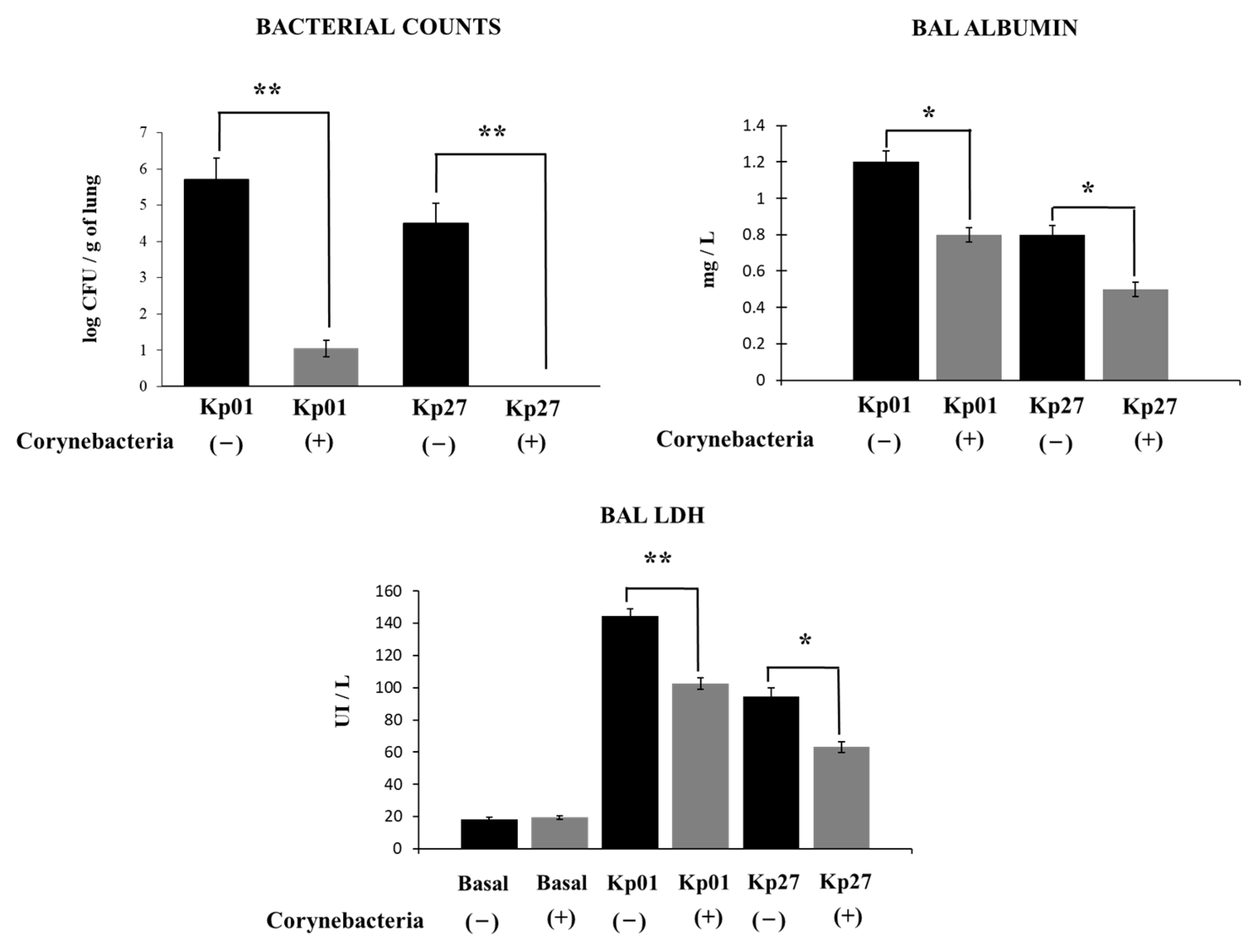
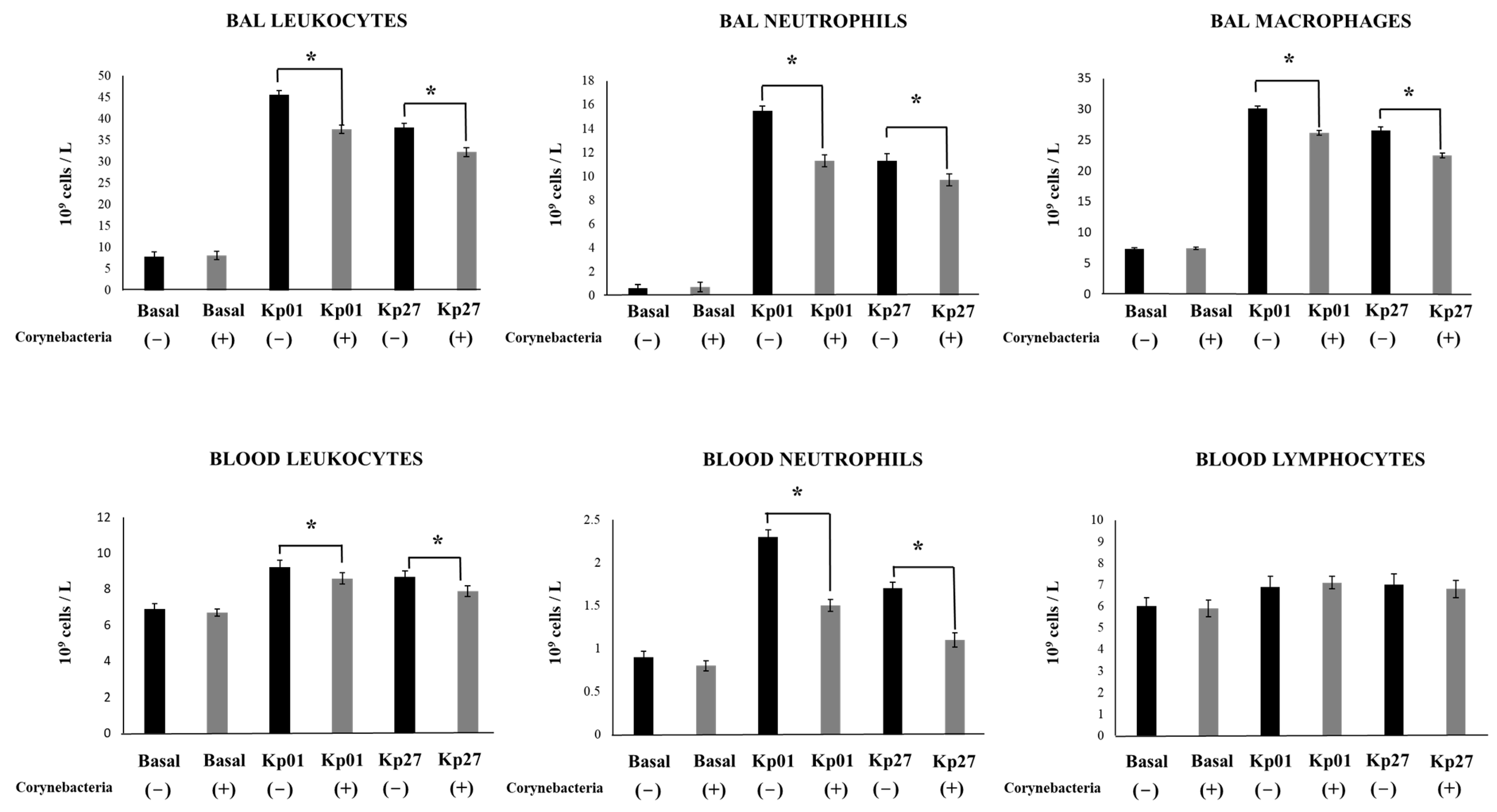
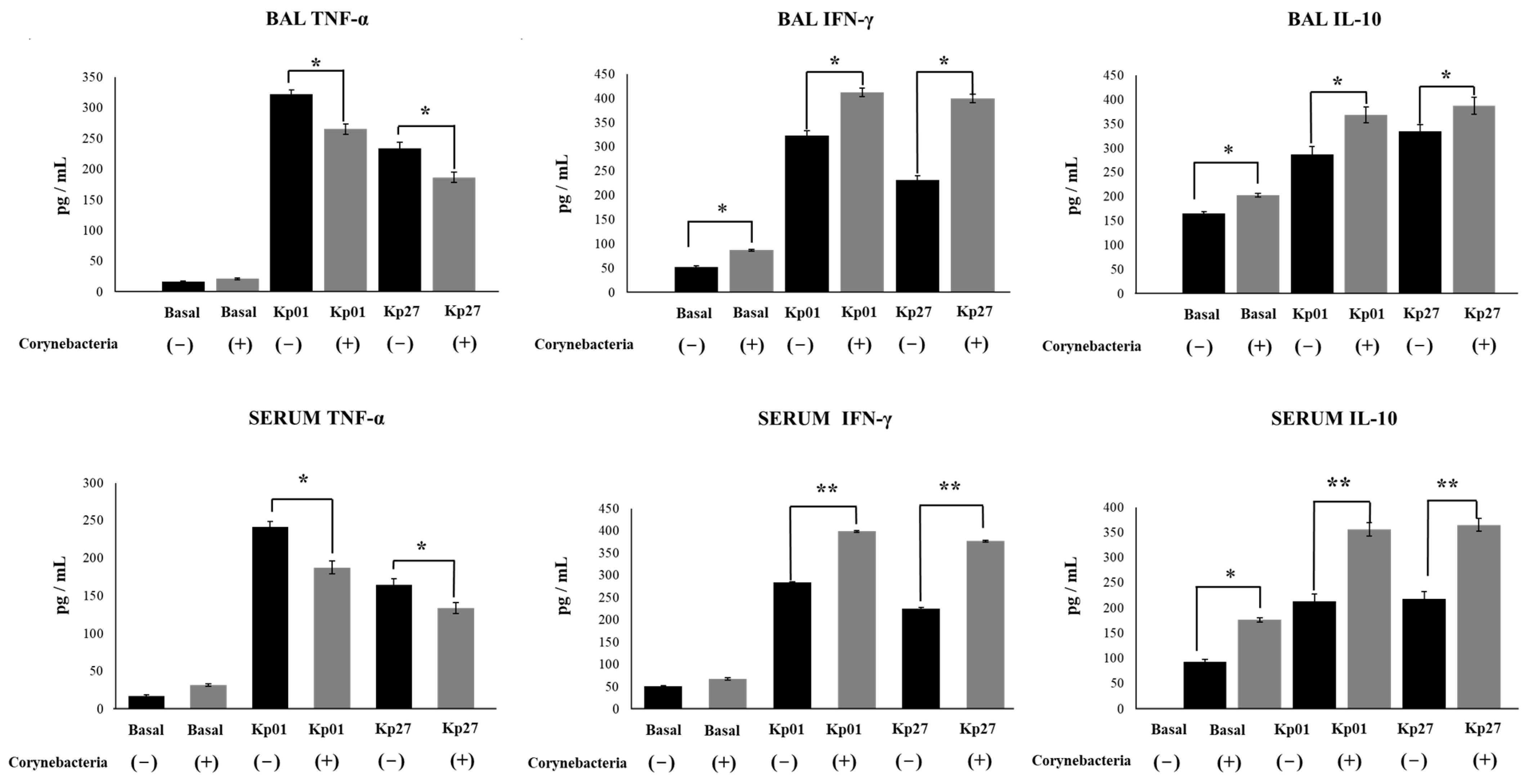
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gomez, S.A.; Pasteran, F.G.; Faccone, D.; Tijet, N.; Rapoport, M.; Lucero, C.; Lastovetska, O.; Albornoz, E.; Galas, M.; KPC Group; et al. Clonal dissemination of Klebsiella pneumoniae ST258 harbouring KPC-2 in Argentina. Clin. Microbiol. Infect. 2011, 17, 1520–1524. [Google Scholar] [CrossRef] [PubMed]
- Cejas, D.; Fernandez Canigia, L.; Nastro, M.; Rodríguez, C.; Tanco, A.; Rodríguez, H.; Vay, C.; Maldonado, I.; Famiglietti, A.; Giovanakis, M.; et al. Hyperendemic Clone of KPC producing Klebsiella pneumoniae ST 258 in Buenos Aires hospitals. Infect. Genet. Evol. 2012, 12, 499–501. [Google Scholar] [CrossRef] [PubMed]
- Cejas, D.; Elena, A.; Guevara Nuñez, D.; Sevillano Platero, P.; De Paulis, A.; Magariños, F.; Alfonso, C.; Berger, M.A.; Fernández-Canigia, L.; Gutkind, G.; et al. Changing epidemiology of KPC-producing Klebsiella pneumoniae in Argentina: Emergence of hypermucoviscous ST25 and high-risk clone ST307. J. Glob. Antimicrob. Resist. 2019, 18, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Vargas, J.M.; Moreno Mochi, M.P.; Nuñez, J.M.; Cáceres, M.; Mochi, S.; Del Campo Moreno, R.; Jure, M.A. Virulence factors and clinical patterns of multiple-clone hypermucoviscous KPC-2 producing K. pneumoniae. Heliyon 2019, 5, e01829. [Google Scholar] [CrossRef]
- Jure, M.A.; Castillo, M.E.; Musa, H.E.; López, C.G.; Cáceres, M.A.; Mochi, S.D.; Bousquet, A.A.; Genel, N.A.; Arlet, G.A.; Decré, D.C. Novel patterns in the molecular epidemiology of KPC-producing Klebsiella pneumoniae in Tucumán, Argentina. J. Glob. Antimicrob. Resist. 2019, 19, 183–187. [Google Scholar] [CrossRef]
- Jure, M.A.; Albarracin, L.; Vargas, J.M.; Maidana, S.D.; Zamar, J.C.; Kitazawa, H.; Villena, J. Draft genome sequences of two hypermucoviscous carbapenem-resistant ST25 Klebsiella Pneumoniae strains causing respiratory and systemic infections. J. Glob. Antimicrob. Resist. 2021, 26, 174–176. [Google Scholar] [CrossRef]
- Villena, J.; Oliveira, M.L.S.; Ferreira, P.C.D.; Salva, S.; Alvarez, S. Lactic acid bacteria in the prevention of pneumococcal respiratory infection: Future opportunities and challenges. Int. Immunopharmacol. 2011, 11, 1633–1645. [Google Scholar] [CrossRef]
- Salva, S.; Kolling, Y.; Ivir, M.; Gutiérrez, F.; Alvarez, S. The role of immunobiotics and postbiotics in the recovery of immune cell populations from respiratory mucosa of malnourished hosts: Effect on the resistance against respiratory infections. Front. Nutr. 2021, 8, 704868. [Google Scholar] [CrossRef]
- Zelaya, H.; Alvarez, S.; Kitazawa, H.; Villena, J. Respiratory antiviral immunity and immunobiotics: Beneficial effects on inflammation-coagulation interaction during Influenza Virus infection. Front. Immunol. 2016, 7, 633. [Google Scholar] [CrossRef]
- Kitazawa, H.; Villena, J. Modulation of respiratory TLR3-anti-viral response by probiotic microorganisms: Lessons learned from Lactobacillus rhamnosus CRL1505. Front. Immunol. 2014, 5, 201. [Google Scholar] [CrossRef] [Green Version]
- Villena, J.; Kitazawa, H. The modulation of mucosal antiviral immunity by immunobiotics: Could they offer any benefit in the SARS-CoV-2 pandemic? Front. Physiol. 2020, 11, 699. [Google Scholar] [CrossRef] [PubMed]
- Lagrafeuille, R.; Miquel, S.; Balestrino, D.; Vareille-Delarbre, M.; Chain, F.; Langella, P.; Forestier, C. Opposing effect of lactobacillus on in vitro Klebsiella pneumoniae in biofilm and in an in vivo intestinal colonization model. Benef. Microbes 2018, 9, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Raras, T.Y.M.; Firdausy, A.F.; Kinant, I.R.; Noorhamdani, N. Anti-biofilm activity of lactic acid bacteria isolated from kefir against multidrug-resistant Klebsiella pneumoniae. J. Pure Appl. Microbiol. 2019, 13, 983–992. [Google Scholar] [CrossRef]
- El-Mokhtar, M.A.; Hassanein, K.M.; Ahmed, A.S.; Gad, G.F.; Amin, M.M.; Hassanein, O.F. Antagonistic activities of cell-free supernatants of Lactobacilli against extended-spectrum β-lactamase producing Klebsiella pneumoniae and Pseudomonas aeruginosa. Infect. Drug Resist. 2020, 13, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Lu, Y.; Wu, X.; Yu, P.; Lan, P.; Wu, X.; Jiang, Y.; Li, Q.; Pi, X.; Liu, W.; et al. Anticolonization of carbapenem-resistant Klebsiella pneumoniae by Lactobacillus plantarum LP1812 through accumulated acetic acid in mice intestinal. Front. Cell Infect. Microbiol. 2021, 11, 804253. [Google Scholar] [CrossRef]
- Ermolenko, E.; Rybalchenko, O.; Borshev, Y.; Tarasova, E.; Kramskaya, T.; Leontieva, G.; Kotyleva, M.; Orlova, O.; Abdurasulova, I.; Suvorov, A. Influence of monostrain and multistrain probiotics on immunity, intestinal ultrastructure and microbiota in experimental dysbiosis. Benef. Microbes 2018, 9, 937–949. [Google Scholar] [CrossRef] [PubMed]
- Savinova, O.S.; Glazunova, O.A.; Moiseenko, K.V.; Begunova, A.V.; Rozhkova, I.V.; Fedorova, T.V. Exoproteome analysis of antagonistic interactions between the probiotic bacteria Limosilactobacillus reuteri LR1 and Lacticaseibacillus rhamnosus F and multidrug resistant strain of Klebsiella pneumoniae. Int. J. Mol. Sci. 2021, 22, 10999. [Google Scholar] [CrossRef]
- Vareille-Delarbre, M.; Miquel, S.; Garcin, S.; Bertran, T.; Balestrino, D.; Evrard, B.; Forestier, C. Immunomodulatory effects of Lactobacillus plantarum on inflammatory response induced by Klebsiella pneumoniae. Infect. Immun. 2019, 87, e00570-19. [Google Scholar] [CrossRef]
- Kanmani, P.; Clua, P.; Vizoso-Pinto, M.G.; Rodriguez, C.; Alvarez, S.; Melnikov, V.; Takahashi, H.; Kitazawa, H.; Villena, J. Respiratory commensal bacteria Corynebacterium pseudodiphtheriticum improves resistance of infant mice to respiratory Syncytial Virus and Streptococcus pneumoniae superinfection. Front. Microbiol. 2017, 8, 1613. [Google Scholar] [CrossRef]
- Shon, A.S.; Bajwa, R.P.; Russo, T.A. Hypervirulent (hypermucoviscous) Klebsiella pneumoniae: A new and dangerous breed. Virulence 2013, 4, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Albarracin, L.; Ortiz Moyano, R.; Vargas, J.M.; Andrade, B.G.N.; Cortez Zamar, J.; Dentice Maidana, S.; Fukuyama, K.; Kurata, S.; Jure, M.Á.; Kitazawa, H.; et al. Genomic and immunological characterization of hypermucoviscous carbapenem-resistant Klebsiella pneumoniae ST25 isolates from Northwest Argentina. Int. J. Mol. Sci. 2022, 23, 7361. [Google Scholar] [CrossRef] [PubMed]
- Ortiz Moyano, R.; Raya Tonetti, F.; Tomokiyo, M.; Kanmani, P.; Vizoso-Pinto, M.G.; Kim, H.; Quilodrán-Vega, S.; Melnikov, V.; Alvarez, S.; Takahashi, H.; et al. The ability of respiratory commensal bacteria to beneficially modulate the lung innate immune response is a strain dependent characteristic. Microorganisms 2020, 8, 727. [Google Scholar] [CrossRef] [PubMed]
- Raya Tonetti, F.; Tomokiyo, M.; Ortiz Moyano, R.; Quilodrán-Vega, S.; Yamamuro, H.; Kanmani, P.; Melnikov, V.; Kurata, S.; Kitazawa, H.; Villena, J. The respiratory commensal bacterium Dolosigranulum pigrum 040417 improves the innate immune response to Streptococcus pneumoniae. Microorganisms 2021, 9, 1324. [Google Scholar] [CrossRef]
- Lasko, M.J.; Abdelraouf, K.; Nicolau, D.P. Comparative in vivo activity of human-simulated plasma and epithelial lining fluid exposures of WCK 5222 (cefepime/zidebactam) against KPC- and OXA-48-like-producing Klebsiella pneumoniae in the neutropenic murine pneumonia model. J. Antimicrob. Chemother. 2021, 76, 2310–2316. [Google Scholar] [CrossRef]
- Tan, S.; Gao, J.; Li, Q.; Guo, T.; Dong, X.; Bai, X.; Yang, J.; Hao, S.; He, F. Synergistic effect of chlorogenic acid and levofloxacin against Klebsiella pneumonia infection in vitro and in vivo. Sci. Rep. 2020, 10, 20013. [Google Scholar] [CrossRef]
- Xiong, H.; Carter, R.A.; Leiner, I.M.; Tang, Y.-W.; Chen, L.; Kreiswirth, B.N.; Pamer, E.G. Distinct contributions of neutrophils and CCR2+ monocytes to pulmonary clearance of different Klebsiella pneumoniae strains. Infect. Immun. 2015, 83, 3418–3427. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Jiang, C.; Xie, N.; Xu, Y.; Liu, L.; Liu, N. Treatment with andrographolide sulfonate provides additional benefits to imipenem in a mouse model of Klebsiella pneumoniae pneumonia. Biomed. Pharmacother. 2019, 117, 109065. [Google Scholar] [CrossRef]
- Meijer, M.T.; de Vos, A.F.; Scicluna, B.P.; Roelofs, J.J.; Abou Fayçal, C.; Orend, G.; Uhel, F.; van der Poll, T. Tenascin-C deficiency is associated with reduced bacterial outgrowth during Klebsiella pneumoniae-evoked pneumosepsis in mice. Front. Immunol. 2021, 12, 600979. [Google Scholar] [CrossRef]
- McDaniel, D.K.; Allen, I.C. Using Klebsiella pneumoniae to model acute lung inflammation in mice. Methods Mol. Biol. 2019, 1960, 169–180. [Google Scholar] [CrossRef]
- Ramirez-Moral, I.; Blok, D.C.; Bernink, J.H.; Garcia-Laorden, M.I.; Florquin, S.; Boon, L.; van’t Veer, C.; Mack, M.; Saluzzo, S.; Knapp, S.; et al. Interleukin-33 improves local immunity during gram-negative pneumonia by a combined effect on neutrophils and inflammatory monocytes. J. Pathol. 2021, 253, 374–383. [Google Scholar] [CrossRef]
- Hsieh, P.-F.; Lin, T.-L.; Lee, C.-Z.; Tsai, S.-F.; Wang, J.-T. Serum-induced iron-acquisition systems and TonB contribute to virulence in Klebsiella pneumoniae causing primary pyogenic liver abscess. J. Infect. Dis. 2008, 197, 1717–1727. [Google Scholar] [CrossRef] [PubMed]
- El Fertas-Aissani, R.; Messai, Y.; Alouache, S.; Bakour, R. Virulence profiles and antibiotic susceptibility patterns of Klebsiella pneumoniae strains isolated from different clinical specimens. Pathol. Biol. 2013, 61, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Lavender, H.; Jagnow, J.J.; Clegg, S. Klebsiella pneumoniae type 3 fimbria-mediated immunity to infection in the murine model of respiratory disease. Int. J. Med. Microbiol. 2005, 295, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Holt, K.E.; Wertheim, H.; Zadoks, R.N.; Baker, S.; Whitehouse, C.A.; Dance, D.; Jenney, A.; Connor, T.R.; Hsu, L.Y.; Severin, J.; et al. Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proc. Natl. Acad. Sci. USA 2015, 112, E3574–E3581. [Google Scholar] [CrossRef] [PubMed]
- Clarke, T.B. Early innate immunity to bacterial infection in the lung is regulated systemically by the commensal microbiota via NOD-like receptor ligands. Infect. Immunol. 2014, 82, 4596–4606. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Chen, Z.; Yuan, Y.; Jing, H.; Zou, J.; Zhang, X.; Zeng, X.; Zhang, W.; Zou, Q.; Zhang, J. Innate immune effectors play essential roles in acute respiratory infection caused by Klebsiella pneumoniae. J. Immunol. Res. 2020, 2020, 5291714. [Google Scholar] [CrossRef]
- Olonisakin, T.F.; Li, H.; Xiong, Z.; Kochman, E.J.K.; Yu, M.; Qu, Y.; Hulver, M.; Kolls, J.K.; St Croix, C.; Doi, Y.; et al. CD36 provides host protection against Klebsiella pneumoniae intrapulmonary infection by enhancing lipopolysaccharide responsiveness and macrophage phagocytosis. J. Infect. Dis. 2016, 214, 1865–1875. [Google Scholar] [CrossRef]
- Andrade, B.G.N.; Cuadrat, R.R.C.; Tonetti, F.R.; Kitazawa, H.; Villena, J. The role of respiratory microbiota in the protection against viral diseases: Respiratory commensal bacteria as next-generation probiotics for COVID-19. Biosci. Microbiota Food Health 2022, 41, 94–102. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dentice Maidana, S.; Ortiz Moyano, R.; Vargas, J.M.; Fukuyama, K.; Kurata, S.; Melnikov, V.; Jure, M.Á.; Kitazawa, H.; Villena, J. Respiratory Commensal Bacteria Increase Protection against Hypermucoviscous Carbapenem-Resistant Klebsiella pneumoniae ST25 Infection. Pathogens 2022, 11, 1063. https://doi.org/10.3390/pathogens11091063
Dentice Maidana S, Ortiz Moyano R, Vargas JM, Fukuyama K, Kurata S, Melnikov V, Jure MÁ, Kitazawa H, Villena J. Respiratory Commensal Bacteria Increase Protection against Hypermucoviscous Carbapenem-Resistant Klebsiella pneumoniae ST25 Infection. Pathogens. 2022; 11(9):1063. https://doi.org/10.3390/pathogens11091063
Chicago/Turabian StyleDentice Maidana, Stefania, Ramiro Ortiz Moyano, Juan Martin Vargas, Kohtaro Fukuyama, Shoichiro Kurata, Vyacheslav Melnikov, María Ángela Jure, Haruki Kitazawa, and Julio Villena. 2022. "Respiratory Commensal Bacteria Increase Protection against Hypermucoviscous Carbapenem-Resistant Klebsiella pneumoniae ST25 Infection" Pathogens 11, no. 9: 1063. https://doi.org/10.3390/pathogens11091063