
A Landscape of CRISPR/Cas Technique for Emerging Viral Disease Diagnostics and Therapeutics: Progress and Prospects
 ,
,  and
and
Abstract
:1. Introduction
2. The Revolution by CRISPR/Cas Techniques

{kind=link}
{kind=link}
{kind=link}
| Phases | Year | Revolutionary Events | References |
|---|---|---|---|
| Identification phase | 1987 | The genome of the bacteria E. coli was where short direct repeats were initially discovered. | [10] |
| 1989 | First application of gene transfer in humans. | [38] | |
| 1990 | First gene delivery for therapeutic intent in ADA-SCID patients. | ||
| 1993 | CRISPR locus described. | [39] | |
| 2000 | Other bacteria and archaea were found to contain the repetitive sequence that Ishino first discovered in E. coli. Gene therapy induced leukaemia in ADA-SCID patients. | ||
| 2002 | Identification of the Cas gene and the proposed name for CRISPR. CRISPR transcript was discovered. | ||
| 2003 | The first information of investigational discovery of a protein related to CRISPR repeats. | ||
| 2005 | Identification of the plasmid and viral sources of spacers. The suggestion that bacteria’s CRISPR/Cas defence mechanism might be adaptive. Recognition of a protospacer-related motif (PAM). | ||
| 2007 | S. thermophilus provides the first experimental confirmation for the CRISPR adaptive immune system. | [40] | |
| 2008 | Recognition of mature CRISPR RNAs (crRNAs) in E. coli as anti-phage defence complex guides with Cas proteins. Researching the Staphylococcus epidermidis Type III (Csm) CRISPR/Cas interference activity. | [41] | |
| 2009 | CRISPR/Cas systems are used to research Pyrococcus furiosus’s antiviral capabilities. Identification of the ssRNA-cleaving type III-B Cmr complex. | [42] | |
| 2010 | Determination of the CRISPR/Cas bacterial immunity cleavage at a position 3 nucleotides before the PAM sequences. CRISPR/Cas 9 is recognised as the prokaryotic adaptive immune system. | [23] | |
| Application phase | 2011 | Classification of the three main category types 1, 2, and 3 of CRISPR/Cas systems. Trans activating CRISPR RNA was discovered (tracrRNA). Using the CRISPR/Cas system from S. thermophilus type II in E. coli and broadcasting that it is functional in several far-related organisms. | [43] |
| 2012 | The modification of the type2 “CRISPR” system (developed by S. pyogenes) for use in cells of mammalian origin. The primary in vitro example of targeting DNA cleavage by CRISPR. A simplification-focused sgRNA construct. | [44] | |
| 2013 | Successfully alteration of the genome in eukaryotic cells using Cas9. | [45] | |
| 2014 | Cas, apo/Cas9, guide RNA, and target RNA were isolated as crystal structures. | [46,47] | |
| 2015 | Chimeric Cmr complex crystal structure; use of CRISPR/Cas9 in humanoid embryo. Scientists used a technique to fix the HBB locus, which when it is mutated, causes thalassemia blood diseases. Due to its off-target behaviours and inability to forecast the results of gene editing using pre-implantation genetic diagnostic (PGD), the experiment was ineffective. | [48] | |
| 2016 | Target recognition and CRISPR/Cas9 nuclear dynamics were discovered in live cells. The NIH gave its clearance to the first CRISPR gene editing human trial. Oncogenic mutations were rendered inactive using CRISPR/Cas9. | [49] | |
| 2017 | The discovery of a particular CRISPR protein (CRISPR/Cas 13) that prefers to target RNA to DNA. Creating CRISPR–gold technology, a highly effective CRISPR/Cas9 variant that uses gold nanoparticles to transport the gene-editing tool to cells. The first time CRISPR/Cas9 was used to correct globin gene mutations in human embryos. The first human embryo with CRISPR germline editing was implanted. Initial HIV-1 therapy CRISPR clinical trial. | [50] | |
| 2018 | Discovered previously present immunoglobulins that target the Cas9 protein. The likelihood that immune responses could render CRISPR/Cas9 gene therapy ineffective. The first CRISPR-based cancer immunotherapy clinical trial. | ||
| 2019 | The first in vivo CRISPR clinical trials for the management of blindness in the “United States” Human cells with Cas12a orthologs showing editing abilities. Several brand-new subtypes of the Type5 CRISPR system are recognized. Cas12k was found to be an RNA-mediated, site-specific incorporation system in E. coli. Several Class 1 CRISPR effectors are used; the Fok I domain is combined. | [51] | |
| 2020 | Nobel Prize for CRISPR/Cas9 genome editing. | [52] |
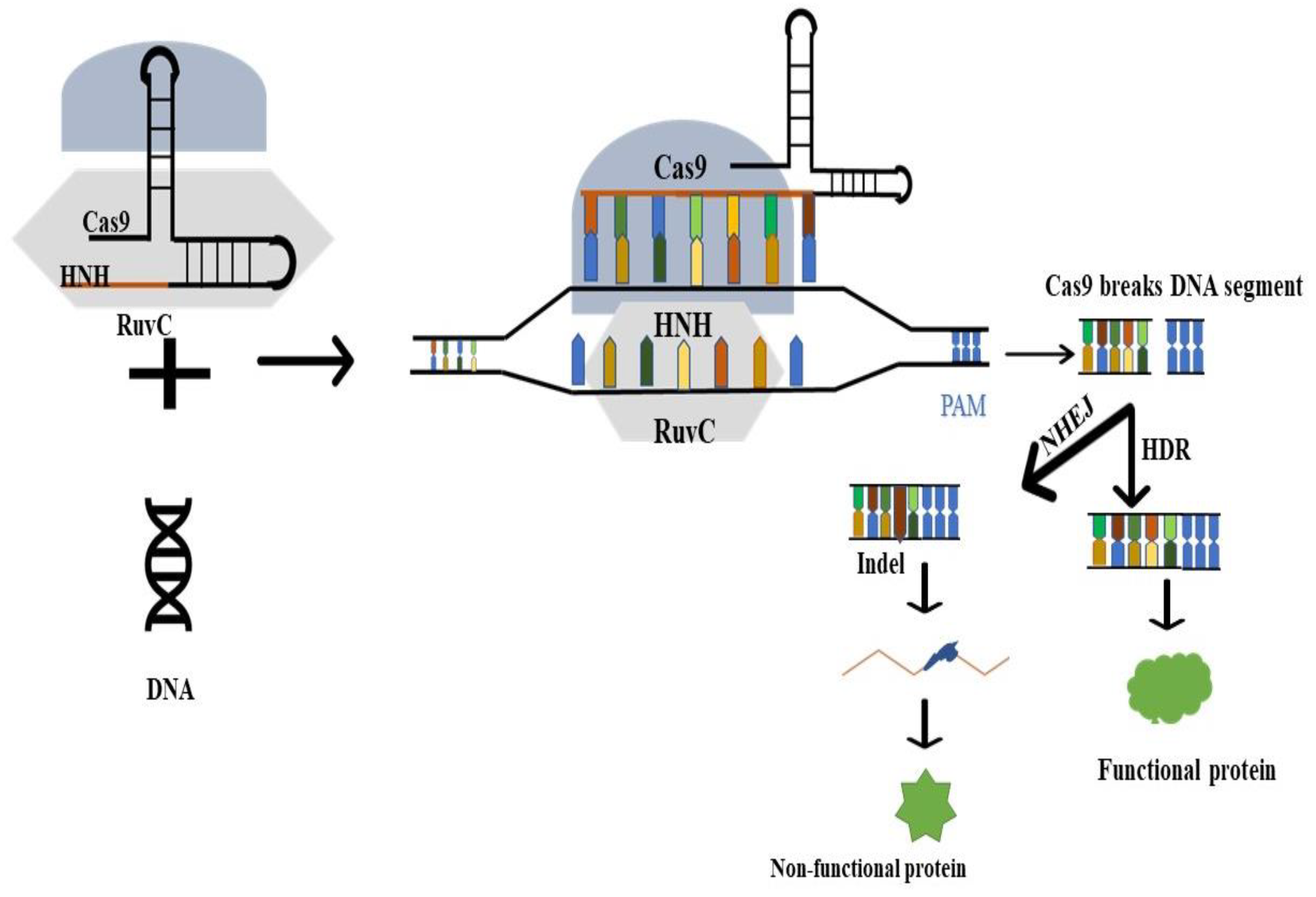
3. CRISPR/Cas9 Gene-Editing Technology
3.1. Structure of CRISPR
3.2. Working Principle
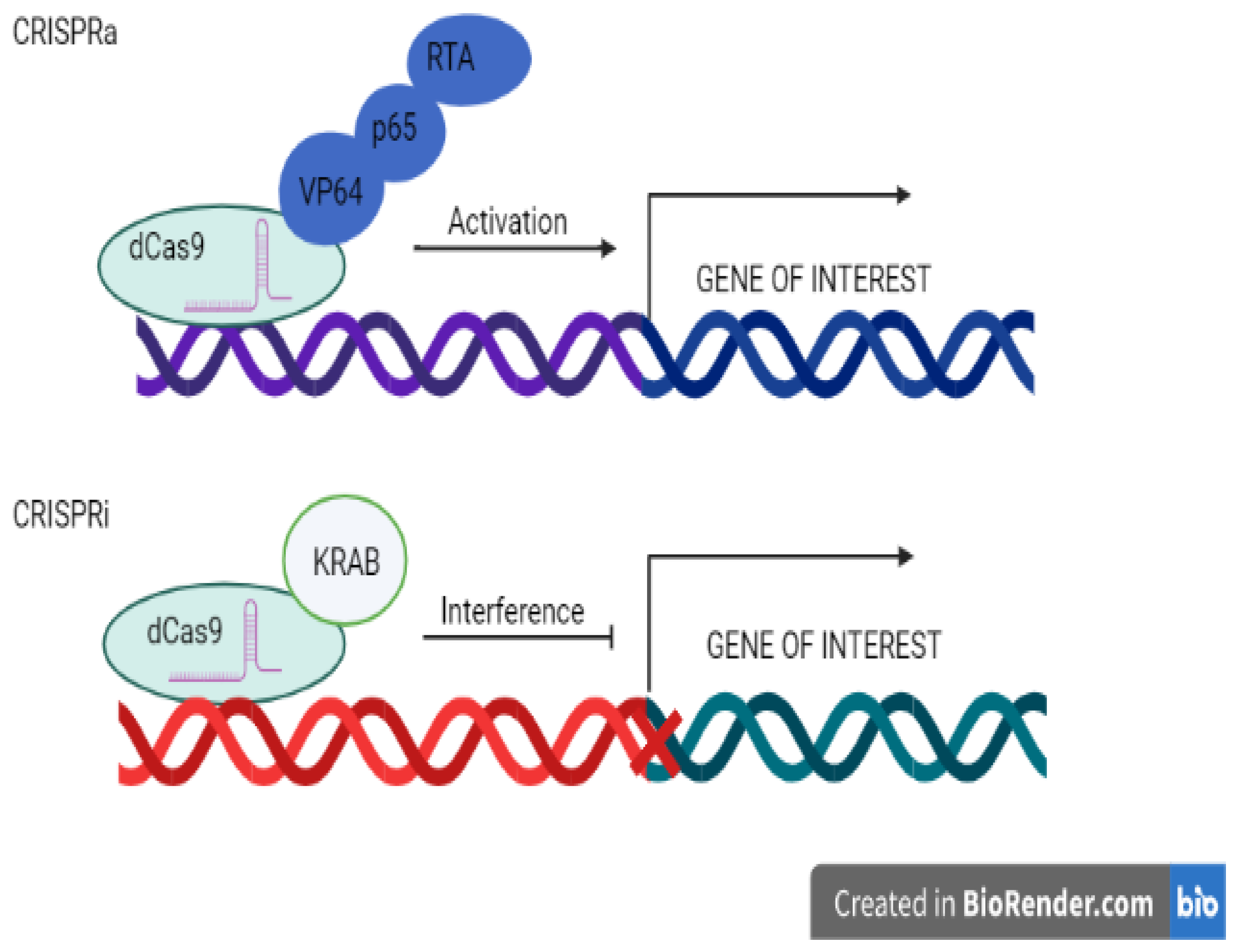
3.3. Transcriptional Regulations
4. CRISPR as a Diagnostic Tool Studying Emerging Viral Infections
4.1. Virus Detection Using Cas9
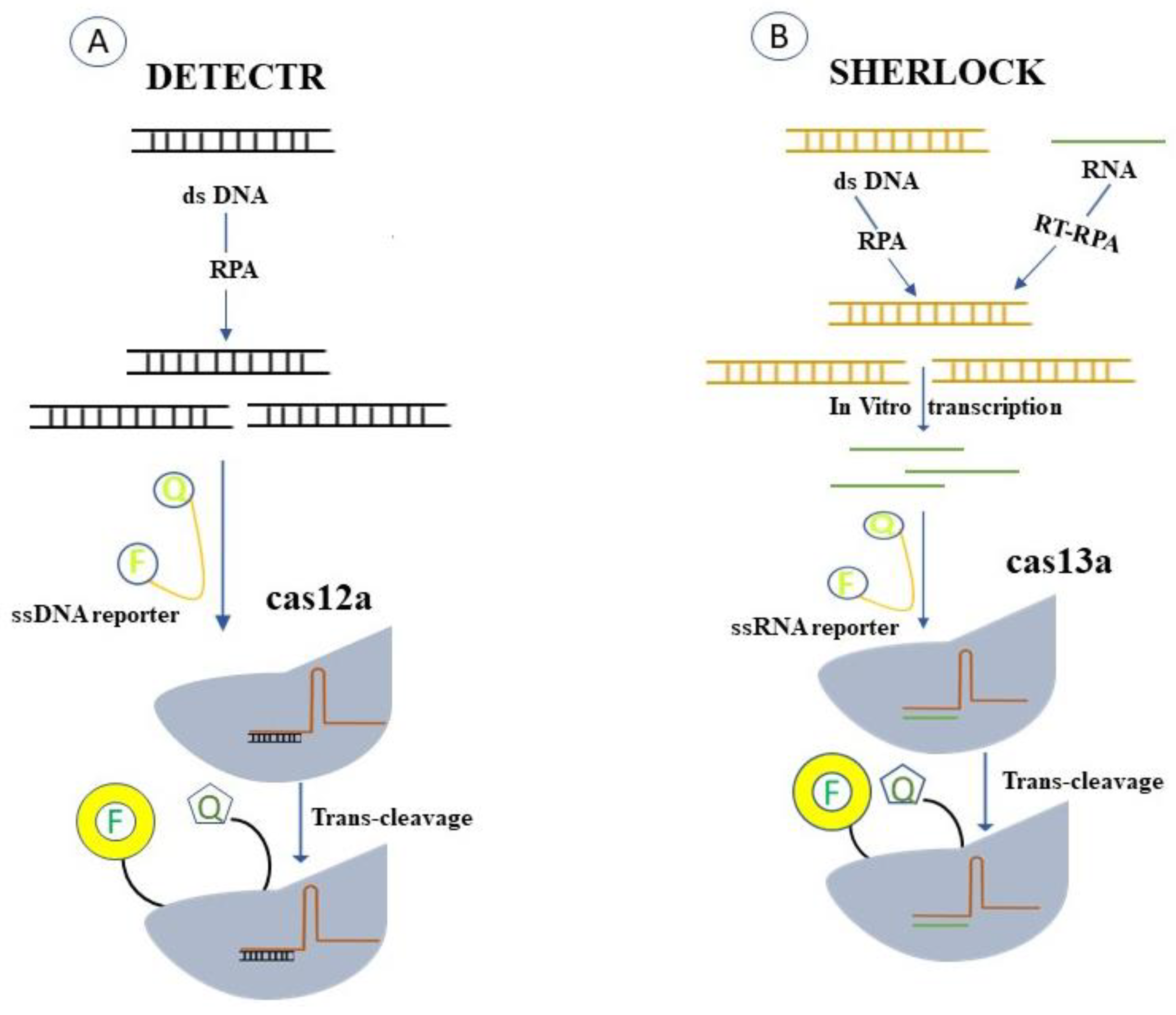
4.2. Cas13- and Cas12-Based Detection Technologies
4.3. Cas13- and Cas12-Based Detection of a Single Virus
5. Utilizing CRISPR/Cas Systems to Fight against Viral Infections
5.1. Human Papillomavirus (HPV)
5.2. Hepatitis B Virus (HBV)
5.3. Human Immunodeficiency Virus (HIV)
5.4. Herpes Simplex Virus (HSV)
5.5. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2)
6. Utilizing CRISPR/Cas9 Technology to Fight Viral Infections in Plants
7. Pros and Cons of CRISPR-Based Diagnosis Systems
8. Bioethical Issues
9. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nakata, A.; Amemura, M.; Makino, K. Unusual Nucleotide Arrangement with Repeated Sequences in the Escherichia Coli K-12 Chromosome. J. Bacteriol. 1989, 171, 3553–3556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mojica, F.J.; Ferrer, C.; Juez, G.; Rodríguez-Valera, F. Long Stretches of Short Tandem Repeats Are Present in the Largest Replicons of the Archaea Haloferax Mediterranei and Haloferax Volcanii and Could Be Involved in Replicon Partitioning. Mol. Microbiol. 1995, 17, 85–93. [Google Scholar] [CrossRef]
- Groenen, P.M.; Bunschoten, A.E.; van Soolingen, D.; van Embden, J.D. Nature of DNA Polymorphism in the Direct Repeat Cluster of Mycobacterium Tuberculosis; Application for Strain Differentiation by a Novel Typing Method. Mol. Microbiol. 1993, 10, 1057–1065. [Google Scholar] [CrossRef]
- Mojica, F.J.; Díez-Villaseñor, C.; Soria, E.; Juez, G. Biological Significance of a Family of Regularly Spaced Repeats in the Genomes of Archaea, Bacteria and Mitochondria. Mol. Microbiol. 2000, 36, 244–246. [Google Scholar] [CrossRef] [PubMed]
- Grissa, I.; Vergnaud, G.; Pourcel, C. The CRISPRdb Database and Tools to Display CRISPRs and to Generate Dictionaries of Spacers and Repeats. BMC Bioinform. 2007, 8, 172. [Google Scholar] [CrossRef] [Green Version]
- Bolotin, A.; Quinquis, B.; Sorokin, A.; Dusko Ehrlich, S. Clustered Regularly Interspaced Short Palindrome Repeats (CRISPRs) Have Spacers of Extrachromosomal Origin. Microbiology 2005, 151, 2551–2561. [Google Scholar] [CrossRef] [Green Version]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR Provides Acquired Resistance Against Viruses in Prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Wolf, Y.I.; Iranzo, J.; Shmakov, S.A.; Alkhnbashi, O.S.; Brouns, S.J.J.; Charpentier, E.; Cheng, D.; Haft, D.H.; Horvath, P.; et al. Evolutionary Classification of CRISPR–Cas Systems: A Burst of Class 2 and Derived Variants. Nat. Rev. Microbiol. 2020, 18, 67–83. [Google Scholar] [CrossRef] [PubMed]
- Ishino, Y.; Krupovic, M.; Forterre, P. History of CRISPR-Cas from Encounter with a Mysterious. J. Bacteriol. 2018, 200, e00580-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakatura, A. Nucleotide Sequence of the Iap Gene, Responsible for Alkaline Phosphatase Isoenzyme Conversion in Escherichia Coli, and Identification of the Gene Product. J. Bacteriol. 1987, 169, 5429–5433. [Google Scholar] [CrossRef]
- Sorek, R.; Kunin, V.; Hugenholtz, P. CRISPR—A Widespread System That Provides Acquired Resistance against Phages in Bacteria and Archaea. Nat. Rev. Microbiol. 2008, 6, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.W.; Kim, S.; Kim, J.M.; Kim, J.S. Targeted Genome Engineering in Human Cells with the Cas9 RNA-Guided Endonuclease. Nat. Biotechnol. 2013, 31, 230–232. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; East, A.; Cheng, A.; Lin, S.; Ma, E.; Doudna, J. RNA-Programmed Genome Editing in Human Cells. eLife 2013, 2, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.; Li, G.; Peng, X.; Deng, A.; Ye, L.; Shi, L.; Wang, T.; He, J. The Use of CRISPR/Cas9 as a Tool to Study Human Infectious Viruses. Front. Cell. Infect. Microbiol. 2021, 11, 590989. [Google Scholar] [CrossRef]
- Torres-ruiz, R.; Rodriguez-perales, S. CRISPR-Cas9 Technology: Applications and Human Disease Modelling. Brief. Funct. Genom. 2016, 16, 4–12. [Google Scholar] [CrossRef] [Green Version]
- Maeder, M.; Therapy, C.G.-M. Genome-Editing Technologies for Gene and Cell Therapy; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Hryhorowicz, M.; Lipiński, D.; Zeyland, J.; Słomski, R. CRISPR/Cas9 Immune System as a Tool for Genome Engineering. Arch. Immunol. Ther. Exp. 2017, 65, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Citorik, R.; Mimee, M.; biotechnology, T.L.-N. Sequence-Specific Antimicrobials Using Efficiently Delivered RNA-Guided Nucleases. Nat. Biotechnol. 2014, 32, 1141–1145. [Google Scholar] [CrossRef] [Green Version]
- Hirakawa, M.P.; Krishnakumar, R.; Timlin, J.A.; Carney, J.P.; Butler, K.S. Gene Editing and CRISPR in the Clinic: Current and Future Perspectives. Biosci. Rep. 2020, 40, BSR20200127. [Google Scholar] [CrossRef] [Green Version]
- Richter, H.; Zoephel, J.; Schermuly, J.; Maticzka, D.; Backofen, R.; Randau, L. Characterization of CRISPR RNA Processing in Clostridium Thermocellum and Methanococcus Maripaludis. Nucleic Acids Res. 2012, 40, 9887–9896. [Google Scholar] [CrossRef] [Green Version]
- Haurwitz, R.E.; Jinek, M.; Wiedenheft, B.; Zhou, K.; Doudna, J.A. Sequence- and Structure-Specific RNA Processing by a CRISPR Endonuclease. Science 2010, 329, 1355–1358. [Google Scholar] [CrossRef]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; Van Der Oost, J.; Regev, A.; et al. Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rath, D.; Amlinger, L.; Rath, A.; Lundgren, M. The CRISPR-Cas Immune System: Biology, Mechanisms and Applications. Biochimie 2015, 117, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Babu, M.; Beloglazova, N.; Flick, R.; Graham, C.; Skarina, T.; Nocek, B.; Gagarinova, A.; Pogoutse, O.; Brown, G.; Binkowski, A.; et al. A Dual Function of the CRISPR–Cas System in Bacterial Antivirus Immunity and DNA Repair. Mol. Microbiol. 2011, 79, 484–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, A.; Goren, M.; Yosef, I.; Auster, O.; Manor, M.; Amitai, G.; Edgar, R.; Qimron, U.; Sorek, R. CRISPR Adaptation Biases Explain Preference for Acquisition of Foreign DNA. Nature 2015, 520, 505–510. [Google Scholar] [CrossRef] [Green Version]
- Perez-Rodriguez, R.; Haitjema, C.; Huang, Q.; Nam, K.H.; Bernardis, S.; Ke, A.; DeLisa, M.P. Envelope Stress Is a Trigger of CRISPR RNA-Mediated DNA Silencing in Escherichia Coli. Mol. Microbiol. 2011, 79, 584–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morens, D.M.; Fauci, A.S. Emerging Infectious Diseases: Threats to Human Health and Global Stability. PLoS Pathog. 2013, 9, e1003467. [Google Scholar] [CrossRef] [Green Version]
- Doerflinger, M.; Forsyth, W.; Ebert, G.; Pellegrini, M.; Herold, M.; Hall, E. CRISPR/Cas9—The Ultimate Weapon to Battle Infectious Diseases? Wiley Online Libr. 2016, 19, e12693. [Google Scholar] [CrossRef] [Green Version]
- White, M.; Hu, W.; Khalili, K. The CRISPR/Cas9 Genome Editing Methodology as a Weapon against Human Viruses. Discov. Med. 2015, 19, 255–262. [Google Scholar]
- Parrish, C.R.; Holmes, E.C.; Morens, D.M.; Park, E.-C.; Burke, D.S.; Calisher, C.H.; Laughlin, C.A.; Saif, L.J.; Daszak, P. Cross-Species Virus Transmission and the Emergence of New Epidemic Diseases. Microbiol. Mol. Biol. Rev. 2008, 72, 457–470. [Google Scholar] [CrossRef] [Green Version]
- Villa, T.; Feijoo-Siota, L.; Pharmacology, J.R.-B. Antivirals against Animal Viruses; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Castilla, V.; Ramirez, J.; Coto, C.E. Plant and Animal Steroids a New Hope to Search for Antiviral Agents. Curr. Med. Chem. 2010, 17, 1858–1873. [Google Scholar] [CrossRef]
- Costa-Neto, E.M. Animal-Based Medicines: Biological Prospection and the Sustainable Use of Zootherapeutic Resources. An. Acad. Bras. Cienc. 2005, 77, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Kuprash, D.V.; Garib, F.Y.; Nedospasov, S.A. Antibody-Based Drugs and Other Recombinant Proteins for Diagnostics and Therapy of Viral Infections, Autoimmune Diseases and Cancer. Mol. Biol. 2017, 51, 769–771. [Google Scholar] [CrossRef]
- Zündorf, I.; Dingermann, T. New Genetically Engineered Drugs. Pharm. Unserer Zeit 2000, 29, 167–173. [Google Scholar] [CrossRef]
- Guo, N.; Liu, J.B.; Li, W.; Ma, Y.S.; Fu, D. The Power and the Promise of CRISPR/Cas9 Genome Editing for Clinical Application with Gene Therapy. J. Adv. Res. 2021, 40, 135–152. [Google Scholar] [CrossRef]
- Nidhi, S.; Anand, U.; Oleksak, P.; Tripathi, P.; Lal, J.A.; Thomas, G.; Kuca, K.; Tripathi, V. Novel Crispr–Cas Systems: An Updated Review of the Current Achievements, Applications, and Future Research Perspectives. Int. J. Mol. Sci. 2021, 22, 3327. [Google Scholar] [CrossRef]
- Scheller, E.L.; Krebsbach, P.H. Gene Therapy: Design and Prospects for Craniofacial Regeneration. J. Dent. Res. 2009, 88, 585–596. [Google Scholar] [CrossRef]
- Mojica, F.J.M.; Díez-Villaseñor, C.; García-Martínez, J.; Soria, E. Intervening Sequences of Regularly Spaced Prokaryotic Repeats Derive from Foreign Genetic Elements. J. Mol. Evol. 2005, 60, 174–182. [Google Scholar] [CrossRef]
- Makarova, K.S.; Haft, D.H.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Horvath, P.; Moineau, S.; Mojica, F.J.M.; Wolf, Y.I.; Yakunin, A.F.; et al. Evolution and Classification of the CRISPR-Cas Systems. Nat. Rev. Microbiol. 2011, 9, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Marraffini, L.A.; Sontheimer, E.J. CRISPR Interference Limits Horizontal Gene Transfer in Staphylococci by Targeting DNA. Science 2008, 322, 1843–1845. [Google Scholar] [CrossRef] [Green Version]
- Hale, C.R.; Zhao, P.; Olson, S.; Duff, M.O.; Graveley, B.R.; Wells, L.; Terns, R.M.; Terns, M.P. RNA-Guided RNA Cleavage by a CRISPR RNA-Cas Protein Complex. Cell 2009, 139, 945–956. [Google Scholar] [CrossRef] [Green Version]
- Carte, J.; Christopher, R.T.; Smith, J.T.; Olson, S.; Barrangou, R.; Moineau, S.; Iii, C.V.C.G.; Graveley, B.R.; Terns, R.M.; Terns, M.P. The three major types of CRISPR-Cas systems function independently in CRISPR RNA biogenesis in Streptococcus thermophilus. Mol. Microbiol. 2014, 93, 98–112. [Google Scholar] [CrossRef] [PubMed]
- Marraffini, L.A. The CRISPR-Cas System of Streptococcus Pyogenes: Function and Applications. In Streptococcus Pyogenes: Basic Biology to Clinical Manifestations; The University of Oklahoma Health Sciences Center: Oklahoma City, OK, USA, 2016; pp. 1–14. [Google Scholar]
- Simonson, R.E. Variations on the Theme Mental Health in Nursing. Public Health Rep. 1956, 71, 700–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osawa, T.; Inanaga, H.; Sato, C.; Numata, T. Crystal Structure of the Crispr-Cas RNA Silencing Cmr Complex Bound to a Target Analog. Mol. Cell 2015, 58, 418–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uddin, F.; Rudin, C.M.; Sen, T. CRISPR Gene Therapy: Applications, Limitations, and Implications for the Future. Front. Oncol. 2020, 10, 1387. [Google Scholar] [CrossRef] [PubMed]
- Pavani, G.; Fabiano, A.; Laurent, M.; Amor, F.; Cantelli, E.; Chalumeau, A.; Maule, G.; Tachtsidi, A.; Concordet, J.P.; Cereseto, A.; et al. Correction of B-Thalassemia by CRISPR/Cas9 Editing of the a-Globin Locus in Human Hematopoietic Stem Cells. Blood Adv. 2021, 5, 1137–1153. [Google Scholar] [CrossRef]
- Reardon, S. First CRISPR Clinical Trial Gets Green Light from US Panel. Nature 2020, 9, 1–2. [Google Scholar] [CrossRef]
- Rose, B.I.; Brown, S. Genetically Modified Babies and a First Application of Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR-Cas9). Obstet. Gynecol. 2019, 134, 157–162. [Google Scholar] [CrossRef]
- Ledford, H. CRISPR Treatment Inserted Directly into the Body for First Time. Nature 2020, 579, 185. [Google Scholar] [CrossRef] [Green Version]
- Westermann, L.; Neubauer, B.; Köttgen, M. Nobel Prize 2020 in Chemistry Honors CRISPR: A Tool for Rewriting the Code of Life. Pflugers Arch. Eur. J. Physiol. 2021, 473, 1–2. [Google Scholar] [CrossRef]
- Karre, A. Gene Editing Technology. 2020. Available online: https://www.researchgate.net/publication/347442835_GENE_EDITING_TECHNOLOGY (accessed on 10 November 2022).
- Crudele, J.M.; Chamberlain, J.S. Cas9 Immunity Creates Challenges for CRISPR Gene Editing Therapies. Nat. Commun. 2018, 9, 3497. [Google Scholar] [CrossRef] [Green Version]
- Arora, L.; Narula, A. Gene Editing and Crop Improvement Using CRISPR-Cas9 System. Front. Plant Sci. 2017, 8, 1932. [Google Scholar] [CrossRef] [PubMed]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome Engineering Using the CRISPR-Cas9 System. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bialk, P.; Rivera-Torres, N.; Strouse, B.; Kmiec, E.B. Regulation of Gene Editing Activity Directed by Single-Stranded Oligonucleotides and CRISPR/Cas9 Systems. PLoS ONE 2015, 10, e0129308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, P.; Barrangou, R. CRISPR/Cas, the Immune System of Bacteria and Archaea. Science 2010, 327, 167–170. [Google Scholar] [CrossRef] [Green Version]
- Sander, J.D.; Joung, J.K. CRISPR-Cas Systems for Editing, Regulating and Targeting Genomes. Nat. Biotechnol. 2014, 32, 347–355. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, C.; Zhao, W. Recent Advances of the Biological and Biomedical Applications of CRISPR/Cas Systems. Mol. Biol. Rep. 2022, 49, 7087–7100. [Google Scholar] [CrossRef]
- Kweon, J.; Yoon, J.K.; Jang, A.H.; Shin, H.R.; See, J.E.; Jang, G.; Kim, J.I.; Kim, Y. Engineered Prime Editors with PAM Flexibility. Mol. Ther. 2021, 29, 2001–2007. [Google Scholar] [CrossRef]
- Fu, Y.W.; Dai, X.Y.; Wang, W.T.; Yang, Z.X.; Zhao, J.J.; Zhang, J.P.; Wen, W.; Zhang, F.; Oberg, K.C.; Zhang, L.; et al. Dynamics and Competition of CRISPR-Cas9 Ribonucleoproteins and AAV Donor-Mediated NHEJ, MMEJ and HDR Editing. Nucleic Acids Res. 2021, 49, 969–985. [Google Scholar] [CrossRef]
- Barman, A.; Deb, B.; Chakraborty, S. A Glance at Genome Editing with CRISPR–Cas9 Technology. Curr. Genet. 2019, 66, 447–462. [Google Scholar] [CrossRef]
- Kirby, E.N.; Shue, B.; Thomas, P.Q.; Beard, M.R. CRISPR Tackles Emerging Viral Pathogens. Viruses 2021, 13, 2157. [Google Scholar] [CrossRef]
- Uppada, V.; Gokara, M.; Rasineni, G.K. Diagnosis and Therapy with CRISPR Advanced CRISPR Based Tools for Point of Care Diagnostics and Early Therapies. Geme 2018, 656, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Agrotis, A.; Ketteler, R. A New Age in Functional Genomics Using CRISPR/Cas9 in Arrayed Library Screening. Front. Genet. 2015, 6, 300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ménová, P.; Raindlová, V.; Hocek, M. Scope and Limitations of the Nicking Enzyme Amplification Reaction for the Synthesis of Base-Modified Oligonucleotides and Primers for PCR. Bioconjug. Chem. 2013, 24, 1081–1093. [Google Scholar] [CrossRef] [PubMed]
- Piepenburg, O.; Williams, C.H.; Stemple, D.L.; Armes, N.A. DNA Detection Using Recombination Proteins. PLoS Biol. 2006, 4, e204. [Google Scholar] [CrossRef]
- Compton, J. Nucleic Acid Sequence-Based Amplification. Nature 1991, 350, 91–92. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-Mediated Isothermal Amplification of DNA. Nucleic Acids Res. 2000, 28, E63. [Google Scholar] [CrossRef] [Green Version]
- Kurosaki, Y.; Martins, D.B.G.; Kimura, M.; Catena, A.d.S.; Borba, M.A.C.S.M.; Mattos, S.d.S.; Abe, H.; Yoshikawa, R.; de Lima Filho, J.L.; Yasuda, J. Development and Evaluation of a Rapid Molecular Diagnostic Test for Zika Virus Infection by Reverse Transcription Loop-Mediated Isothermal Amplification. Sci. Rep. 2017, 7, 13503. [Google Scholar] [CrossRef] [Green Version]
- Patel, P.; Abd El Wahed, A.; Faye, O.; Prüger, P.; Kaiser, M.; Thaloengsok, S.; Ubol, S.; Sakuntabhai, A.; Leparc-Goffart, I.; Hufert, F.T.; et al. A Field-Deployable Reverse Transcription Recombinase Polymerase Amplification Assay for Rapid Detection of the Chikungunya Virus. PLoS Negl. Trop. Dis. 2016, 10, e0004953. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.S.; Ma, E.; Harrington, L.B.; Da Costa, M.; Tian, X.; Palefsky, J.M.; Doudna, J.A. CRISPR-Cas12a Target Binding Unleashes Indiscriminate Single-Stranded DNase Activity. Science 2018, 360, 436. [Google Scholar] [CrossRef] [Green Version]
- East-Seletsky, A.; O’Connell, M.R.; Knight, S.C.; Burstein, D.; Cate, J.H.D.; Tjian, R.; Doudna, J.A. Two Distinct RNase Activities of CRISPR-C2c2 Enable Guide-RNA Processing and RNA Detection. Nature 2016, 538, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Gootenberg, C.; Abudayyeh, O.O.; Kellner, M.J.; Joung, J.; Collins, J.J.; Zhang, F. Nucleic Acid Detection Platform with Cas13, Cas12a, and Csm6. Science 2018, 360, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, B.; Xu, X.; Long, F.; Wang, J. CRISPR-Typing PCR (CtPCR), a New Cas9-Based DNA Detection Method. Sci. Rep. 2018, 8, 14126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.-F.; Chen, G.-J.; Luo, Y.-L.; Zhang, Y.; Zhao, G.; Lu, Z.-D.; Czarna, A.; Gu, Z.; Wang, J. Rational Designs of in Vivo CRISPR-Cas Delivery Systems. Adv. Drug Deliv. Rev. 2021, 168, 3–29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Xia, Q.; Wang, Q.; Xia, X.; Wang, J. Detecting and Typing Target DNA with a Novel CRISPR-Typing PCR (CtPCR) Technique. Anal. Biochem. 2018, 561–562, 37–46. [Google Scholar] [CrossRef]
- Pardee, K.; Green, A.A.; Takahashi, M.K.; Braff, D.; Lambert, G.; Lee, J.W.; Ferrante, T.; Ma, D.; Donghia, N.; Fan, M.; et al. Rapid, Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell 2016, 165, 1255–1266. [Google Scholar] [CrossRef] [Green Version]
- Kaminski, M.M.; Abudayyeh, O.O.; Gootenberg, J.S.; Zhang, F.; Collins, J.J. CRISPR-Based Diagnostics. Nat. Biomed. Eng. 2021, 5, 643–656. [Google Scholar] [CrossRef]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A.; et al. Nucleic Acid Detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellner, M.J.; Koob, J.G.; Gootenberg, J.S.; Abudayyeh, O.O.; Zhang, F. SHERLOCK: Nucleic Acid Detection with CRISPR Nucleases. Nat. Protoc. 2019, 14, 2986–3012. [Google Scholar] [CrossRef]
- Li, S.; Ding, L.; Gao, H.; Chen, C.; Liu, Z.; Deng, Z. Adaptive Neural Network Tracking Control-Based Reinforcement Learning for Wheeled Mobile Robots with Skidding and Slipping. Neurocomputing 2018, 283, 20–30. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, R.; Li, J. CRISPR/Cas Systems Redefine Nucleic Acid Detection: Principles and Methods. Biosens. Bioelectron. 2020, 165, 112430. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, S.-X.; Wang, F.; Zeng, M.-S. Room Temperature Detection of Plasma Epstein–Barr Virus DNA with CRISPR–Cas13. Clin. Chem. 2019, 65, 591–592. [Google Scholar] [CrossRef] [PubMed]
- Normandin, E.; Solomon, I.H.; Zamirpour, S.; Lemieux, J.; Freije, C.A.; Mukerji, S.S.; Tomkins-Tinch, C.; Park, D.; Sabeti, P.C.; Piantadosi, A. Powassan Virus Neuropathology and Genomic Diversity in Patients with Fatal Encephalitis. Open Forum Infect. Dis. 2020, 7, ofaa392. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, H.; Liu, C.; Peng, L.; Khan, H.; Cui, L.; Huang, R.; Wu, C.; Shen, S.; Wang, S.; et al. CRISPR-Cas13a Nanomachine Based Simple Technology for Avian Influenza A (H7N9) Virus on-Site Detection. J. Biomed. Nanotechnol. 2019, 15, 790–798. [Google Scholar] [CrossRef] [PubMed]
- Curti, L.A.; Pereyra-Bonnet, F.; Repizo, G.D.; Fay, J.V.; Salvatierra, K.; Blariza, M.J.; Ibañez-Alegre, D.; Rinflerch, A.R.; Miretti, M.; Gimenez, C.A. CRISPR-Based Platform for Carbapenemases and Emerging Viruses Detection Using Cas12a (Cpf1) Effector Nuclease. Emerg. Microbes Infect. 2020, 9, 1140–1148. [Google Scholar] [CrossRef] [PubMed]
- Barnes, K.G.; Lachenauer, A.E.; Nitido, A.; Siddiqui, S.; Gross, R.; Beitzel, B.; Siddle, K.J.; Freije, C.A.; Dighero-Kemp, B.; Mehta, S.B.; et al. Deployable CRISPR-Cas13a Diagnostic Tools to Detect and Report Ebola and Lassa Virus Cases in Real-Time. Nat. Commun. 2020, 11, 4131. [Google Scholar] [CrossRef] [PubMed]
- Broughton, J.P.; Deng, X.; Yu, G.; Fasching, C.L.; Singh, J.; Streithorst, J.; Granados, A.; Sotomayor-Gonzalez, A.; Zorn, K.; Gopez, A.; et al. Rapid Detection of 2019 Novel Coronavirus SARS-CoV-2 Using a CRISPR-Based DETECTR Lateral Flow Assay. medRxiv, 2020; preprint. [Google Scholar] [CrossRef]
- Curti, L.; Pereyra-Bonnet, F.; Gimenez, C.A. An Ultrasensitive, Rapid, and Portable Coronavirus SARS-CoV-2 Sequence Detection Method Based on CRISPR-Cas12. BioRxiv, 2020; preprint. [Google Scholar] [CrossRef] [Green Version]
- Metsky, H.C.; Freije, C.A.; Kosoko-Thoroddsen, T.S.F.; Sabeti, P.C.; Myhrvold, C. CRISPR-Based Surveillance for COVID-19 Using Genomically-Comprehensive Machine Learning Design. BioRxiv, 2020; preprint. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Abudayyeh, O.O.; Gootenberg, J.S.; Sciences, C.; Mathers, L. A Protocol for Detection of COVID-19 Using CRISPR Diagnostics. Bioarchive 2020, 8, 1–8. [Google Scholar]
- Patchsung, M.; Jantarug, K.; Pattama, A.; Aphicho, K.; Suraritdechachai, S.; Meesawat, P.; Sappakhaw, K.; Leelahakorn, N.; Ruenkam, T.; Wongsatit, T.; et al. Clinical Validation of a Cas13-Based Assay for the Detection of SARS-CoV-2 RNA. Nat. Biomed. Eng. 2020, 4, 1140–1149. [Google Scholar] [CrossRef]
- Ackerman, C.M.; Myhrvold, C.; Thakku, S.G.; Freije, C.A.; Metsky, H.C.; Yang, D.K.; Ye, S.H.; Boehm, C.K.; Kosoko-Thoroddsen, T.-S.F.; Kehe, J.; et al. Massively Multiplexed Nucleic Acid Detection with Cas13. Nature 2020, 582, 277–282. [Google Scholar] [CrossRef]
- Ali, Z.; Aman, R.; Mahas, A.; Rao, G.S.; Tehseen, M.; Marsic, T.; Salunke, R.; Subudhi, A.K.; Hala, S.M.; Hamdan, S.M.; et al. ISCAN: An RT-LAMP-Coupled CRISPR-Cas12 Module for Rapid, Sensitive Detection of SARS-CoV-2. Virus Res. 2020, 288, 198129. [Google Scholar] [CrossRef] [PubMed]
- Arizti-Sanz, J.; Freije, C.A.; Stanton, A.C.; Petros, B.A.; Boehm, C.K.; Siddiqui, S.; Shaw, B.M.; Adams, G.; Kosoko-Thoroddsen, T.-S.F.; Kemball, M.E.; et al. Streamlined Inactivation, Amplification, and Cas13-Based Detection of SARS-CoV-2. Nat. Commun. 2020, 11, 5921. [Google Scholar] [CrossRef]
- SARS-CoV-2 RNA Detectr Assay Accelerated Emergency Use Authorization (eua) Summary SARS-CoV-2 RNA Detectr Assay (UCSF Health Clinical Laboratories, UCSF Clinical Labs at China Basin). 1988. Available online: file:///C:/Users/MDPI/Downloads/EUA-UCSF-DETECTR-EUAsum.pdf (accessed on 10 November 2022).
- Bhattacharyya, R.P.; Thakku, S.G.; Hung, D.T. Harnessing CRISPR Effectors for Infectious Disease Diagnostics. ACS Infect. Dis. 2018, 4, 1278–1282. [Google Scholar] [CrossRef] [PubMed]
- Kocak, D.D.; Gersbach, C.A. From CRISPR Scissors to Virus Sensors. Nature 2018, 557, 168–169. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.; Chen, Z.; Shi, L.; Lin, H. Developmental Progress of CRISPR/Cas9 and Its Therapeutic Applications for HIV-1 Infection. Rev. Med. Virol. 2018, 28, e1998. [Google Scholar] [CrossRef]
- Khambhati, K.; Bhattacharjee, G.; Singh, V. Current Progress in CRISPR-based Diagnostic Platforms. J. Cell. Biochem. 2019, 120, 2721. [Google Scholar] [CrossRef] [PubMed]
- Myhrvold, C.; Freije, C.A.; Gootenberg, J.S.; Abudayyeh, O.O.; Metsky, H.C.; Durbin, A.F.; Kellner, M.J.; Tan, A.L.; Paul, L.M.; Parham, L.A.; et al. Field-Deployable Viral Diagnostics Using CRISPR-Cas13. Science 2018, 360, 444–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johns Hopkins Coronavirus Resource Center. COVID-19 Map; Johns Hopkins Coronavirus Resource Center: Baltimore, MD, USA, 2022. [Google Scholar]
- Broughton, J.P.; Deng, X.; Yu, G.; Fasching, C.L.; Servellita, V.; Singh, J.; Miao, X.; Streithorst, J.A.; Granados, A.; Sotomayor-Gonzalez, A.; et al. CRISPR-Cas12-Based Detection of SARS-CoV-2. Nat. Biotechnol. 2020, 38, 870–874. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Prado, E.; Simbaña-Rivera, K.; Gómez- Barreno, L.; Rubio-Neira, M.; Guaman, L.P.; Kyriakidis, N.C.; Muslin, C.; Jaramillo, A.M.G.; Barba-Ostria, C.; Cevallos-Robalino, D.; et al. Clinical, Molecular, and Epidemiological Characterization of the SARS-CoV-2 Virus and the Coronavirus Disease 2019 (COVID-19), a Comprehensive Literature Review. Diagn. Microbiol. Infect. Dis. 2020, 98, 115094. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Upadhyay, D.J.; Srivastava, A. Next-Generation Molecular Diagnostics Development by CRISPR/Cas Tool: Rapid Detection and Surveillance of Viral Disease Outbreaks. Front. Mol. Biosci. 2020, 7, 582499. [Google Scholar] [CrossRef]
- Ebrahimi, S.; Teimoori, A.; Khanbabaei, H.; Tabasi, M. Harnessing CRISPR/Cas 9 System for Manipulation of DNA Virus Genome. Rev. Med. Virol. 2019, 29, e2009. [Google Scholar] [CrossRef]
- Gupta, N.; Parashar, P.; Mittal, M.; Mehra, V.; Khatri, M.; Rajguru, S. Antibacterial Potential of Elletaria Cardamomum, Syzygium Aromaticum and Piper Nigrum, Their Synergistic Effects and Phytochemical Determination. J. Pharm. Res. 2014, 8, 1091–1097. [Google Scholar]
- Ryndock, E.J.; Meyers, C. A Risk for Non-Sexual Transmission of Human Papillomavirus? Expert Rev. Anti. Infect. Ther. 2014, 12, 1165–1170. [Google Scholar] [CrossRef] [PubMed]
- Moens, U. Human Polyomaviruses and Papillomaviruses. Int. J. Mol. Sci. 2018, 19, 2360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C. CRISPR/Cas9-Based Antiviral Strategy: Current Status and the Potential Challenge. Molecules 2019, 24, 1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moody, C.A.; Laimins, L.A. Human Papillomavirus Oncoproteins: Pathways to Transformation. Nat. Rev. Cancer 2010, 10, 550–560. [Google Scholar] [CrossRef]
- Kennedy, E.M.; Cullen, B.R. Gene Editing: A New Tool for Viral Disease. Annu. Rev. Med. 2017, 68, 401–411. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Human Papillomavirus Vaccines: WHO Position Paper, October 2014. Relev. Epidemiol. Hebd. 2014, 89, 465–491. [Google Scholar]
- World Health Organization. Weekly Epidemiological Record = Relevé Épidémiologique Hebdomadaire; WHO: Geneva, Switzerland, 2016; pp. 561–584. [Google Scholar]
- Brotherton, J.M.L.; Zuber, P.L.F.; Bloem, P.J.N. Primary Prevention of HPV through Vaccination: Update on the Current Global Status. Curr. Obstet. Gynecol. Rep. 2016, 5, 210–224. [Google Scholar] [CrossRef]
- Dorji, T.; Tshomo, U.; Phuntsho, S.; Tamang, T.D.; Tshokey, T.; Baussano, I.; Franceschi, S.; Clifford, G. Introduction of a National HPV Vaccination Program into Bhutan. Vaccine 2015, 33, 3726–3730. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin-Drubin, M.E.; Münger, K. Oncogenic Activities of Human Papillomaviruses. Virus Res. 2009, 143, 195–208. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.; Yu, L.; Zhu, D.; Ding, W.; Wang, X.; Zhang, C.; Wang, L.; Jiang, X.; Shen, H.; He, D.; et al. Disruption of HPV16-E7 by CRISPR/Cas System Induces Apoptosis and Growth Inhibition in HPV16 Positive Human Cervical Cancer Cells. Biomed. Res. Int. 2014, 2014, 612823. [Google Scholar] [CrossRef] [Green Version]
- Seo, Y.; Yano, Y. Short- and Long-Term Outcome of Interferon Therapy for Chronic Hepatitis B Infection. World J. Gastroenterol. 2014, 20, 13284–13292. [Google Scholar] [CrossRef] [PubMed]
- Andleeb, S.; Naseer, A.; Ali, S.; Mustafa, R.G.; Zafar, A.; Shafique, I.; Ihsan-ul-Haq; Ismail, M.; Saleem, M.; Mansoor, Q. Biological Activities and Secondary Metabolite Screening of Rumex Hastatus Extract through Fourier Transform Infrared and Raman Spectroscopy. Infect. Disord.-Drug Targets 2018, 18, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Emery, J.S.; Feld, J.J. Treatment of Hepatitis B Virus with Combination Therapy Now and in the Future. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Maepa, M.B.; Roelofse, I.; Ely, A.; Arbuthnot, P. Progress and Prospects of Anti-HBV Gene Therapy Development. Int. J. Mol. Sci. 2015, 16, 17589–17610. [Google Scholar] [CrossRef] [Green Version]
- Weber, N.D.; Stone, D.; Sedlak, R.H.; De Silva Feelixge, H.S.; Roychoudhury, P.; Schiffer, J.T.; Aubert, M.; Jerome, K.R. AAV-Mediated Delivery of Zinc Finger Nucleases Targeting Hepatitis B Virus Inhibits Active Replication. PLoS ONE 2014, 9, e97579. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.R.; Yang, H.C.; Kuo, Y.T.; Liu, C.J.; Yang, T.Y.; Sung, K.C.; Lin, Y.Y.; Wang, H.Y.; Wang, C.C.; Shen, Y.C.; et al. The CRISPR/Cas9 System Facilitates Clearance of the Intrahepatic HBV Templates In Vivo. Mol. Ther. Nucleic Acids 2014, 3, e186. [Google Scholar] [CrossRef]
- Seeger, C.; Sohn, J.A. Targeting Hepatitis B Virus With CRISPR/Cas9. Mol. Ther. Nucleic Acids 2014, 3, e216. [Google Scholar] [CrossRef]
- Karimova, M.; Beschorner, N.; Dammermann, W.; Chemnitz, J.; Indenbirken, D.; Bockmann, J.H.; Grundhoff, A.; Luth, S.; Buchholz, F.; zur Wiesch, J.S.; et al. CRISPR/Cas9 Nickase-Mediated Disruption of Hepatitis B Virus Open Reading Frame S and X. Sci. Rep. 2015, 5, 13734. [Google Scholar] [CrossRef] [Green Version]
- Ramanan, V.; Shlomai, A.; Cox, D.B.T.; Schwartz, R.E.; Michailidis, E.; Bhatta, A.; Scott, D.A.; Zhang, F.; Rice, C.M.; Bhatia, S.N. CRISPR/Cas9 Cleavage of Viral DNA Efficiently Suppresses Hepatitis B Virus. Sci. Rep. 2015, 5, 10833. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Xu, Z.W.; Liu, S.; Zhang, R.Y.; Ding, S.L.; Xie, X.M.; Long, L.; Chen, X.M.; Zhuang, H.; Lu, F.M. Dual GRNAs Guided CRISPR/Cas9 System Inhibits Hepatitis B Virus Replication. World J. Gastroenterol. 2015, 21, 9554. [Google Scholar] [CrossRef]
- Zhen, S.; Lu, J.; Liu, Y.H.; Chen, W.; Li, X. Synergistic Antitumor Effect on Cervical Cancer by Rational Combination of PD1 Blockade and CRISPR-Cas9-Mediated HPV Knockout. Cancer Gene Ther. 2020, 27, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Kostyusheva, A.P.; Brezgin, S.A.; Zarifyan, D.N.; Chistyakov, D.S.; Gegechkory, V.I.; Bayurova, E.O.; Volchkova, E.A.; Kostyushev, D.S.; Chulanov, V.P. Hepatitis B Virus and Site-Specific Nucleases: Effects of Genetic Modifications in CRISPR/Cas9 on Antiviral Activity. Russ. J. Infect. Immun. 2019, 9, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Schinazi, R.F.; Ehteshami, M.; Bassit, L.; Asselah, T. Towards HBV Curative Therapies. Liver Int. 2018, 38, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pattanayak, V.; Lin, S.; Guilinger, J.P.; Ma, E.; Doudna, J.A.; Liu, D.R. High-Throughput Profiling of off-Target DNA Cleavage Reveals RNA-Programmed Cas9 Nuclease Specificity. Nat. Biotechnol. 2013, 31, 839–843. [Google Scholar] [CrossRef] [PubMed]
- Krasnopolsky, S.; Kuzmina, A.; Taube, R. Genome-Wide Crispr Knockout Screen Identifies Znf304 as a Silencer of HIV Transcription That Promotes Viral Latency. PLoS Pathog. 2020, 16, e1008834. [Google Scholar] [CrossRef] [PubMed]
- Dash, P.K.; Kaminski, R.; Bella, R.; Su, H.; Mathews, S.; Ahooyi, T.M.; Chen, C.; Mancuso, P.; Sariyer, R.; Ferrante, P.; et al. Sequential LASER ART and CRISPR Treatments Eliminate HIV-1 in a Subset of Infected Humanized Mice. Nat. Commun. 2019, 10, 2753. [Google Scholar] [CrossRef] [Green Version]
- Campbell-Yesufu, O.T.; Gandhi, R.T. Update on Human Immunodeficiency Virus (HIV)-2 Infection. Clin. Infect. Dis. 2011, 52, 780–787. [Google Scholar] [CrossRef] [Green Version]
- Alimonti, J.B.; Ball, T.B.; Fowke, K.R. Mechanisms of CD4+ T Lymphocyte Cell Death in Human Immunodeficiency Virus Infection and AIDS. J. Gen. Virol. 2003, 84, 1649–1661. [Google Scholar] [CrossRef]
- Arribas, J.R.; Eron, J. Advances in Antiretroviral Therapy. Curr. Opin. HIV AIDS 2013, 8, 341–349. [Google Scholar] [CrossRef]
- Wigdahl, B.; Dampier, W.; Nonnemacher, M.R.; Sullivan Sullivan, N.T.; Jacobson Jacobson, J.M. HIV Excision Utilizing CRISPR/Cas9 Technology: Attacking the Proviral Quasispecies in Reservoirs to Achieve a Cure. MOJ Immunol. 2014, 1, 00022. [Google Scholar] [CrossRef]
- Feinberg, M.B. Changing the Natural History of HIV Disease. Lancet 1996, 348, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Kitahata, M.M.; Koepsell, T.D.; Deyo, R.A.; Maxwell, C.L.; Dodge, W.T.; Wagner, E.H. Physicians’ Experience with the Acquired Immunodeficiency Syndrome as a Factor in Patients’ Survival. N. Engl. J. Med. 1996, 334, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, W.E.; Tisnado, D.M.; Lui, H.H.; Nakazono, T.T.; Carlisle, D.M. The Effect of Hospital Experience on Mortality among Patients Hospitalized with Acquired Immunodeficiency Syndrome in California. Am. J. Med. 1999, 107, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Rackal, J.M.; Tynan, A.-M.; Handford, C.D.; Rzeznikiewiz, D.; Agha, A.; Glazier, R. Provider Training and Experience for People Living with HIV/AIDS. Cochrane Database Syst. Rev. 2011, 6, CD003938. [Google Scholar] [CrossRef] [Green Version]
- Nucleoside Analogues. LiverTox: Clinical and Research Information on Drug-Induced Liver Injury; National Institute of Diabetes and Digestive and Kidney Diseases: Bethesda, MD, USA, 2012. [Google Scholar]
- Lucas, G.M.; Ross, M.J.; Stock, P.G.; Shlipak, M.G.; Wyatt, C.M.; Gupta, S.K.; Atta, M.G.; Wools-Kaloustian, K.K.; Pham, P.A.; Bruggeman, L.A.; et al. Clinical Practice Guideline for the Management of Chronic Kidney Disease in Patients Infected with HIV: 2014 Update by the HIV Medicine Association of the Infectious Diseases Society of America. Clin. Infect. Dis. 2014, 59, e96–e138. [Google Scholar] [CrossRef] [Green Version]
- Lyseng-Williamson, K.A.; Reynolds, N.A.; Plosker, G.L. Tenofovir Disoproxil Fumarate: A Review of Its Use in the Management of HIV Infection. Drugs 2005, 65, 413–432. [Google Scholar] [CrossRef]
- Martin, A.M.; Nolan, D.; Gaudieri, S.; Almeida, C.A.; Nolan, R.; James, I.; Carvalho, F.; Phillips, E.; Christiansen, F.T.; Purcell, A.W.; et al. Predisposition to Abacavir Hypersensitivity Conferred by HLA-B*5701 and a Haplotypic Hsp70-Hom Variant. Proc. Natl. Acad. Sci. USA 2004, 101, 4180–4185. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-F.; Stampley, J.E.; Irving, B.A.; Dugas, T.R. Chronic Nucleoside Reverse Transcriptase Inhibitors Disrupt Mitochondrial Homeostasis and Promote Premature Endothelial Senescence. Toxicol. Sci. 2019, 172, 445–456. [Google Scholar] [CrossRef]
- Ebina, H.; Misawa, N.; Kanemura, Y.; Koyanagi, Y. Harnessing the CRISPR/Cas9 System to Disrupt Latent HIV-1 Provirus. Sci. Rep. 2013, 3, 2510. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R.K.; Peppa, D.; Hill, A.L.; Gálvez, C.; Salgado, M.; Pace, M.; McCoy, L.E.; Griffith, S.A.; Thornhill, J.; Alrubayyi, A.; et al. Evidence for HIV-1 Cure after CCR5Δ32/Δ32 Allogeneic Haemopoietic Stem-Cell Transplantation 30 Months Post Analytical Treatment Interruption: A Case Report. Lancet HIV 2020, 7, e340–e347. [Google Scholar] [CrossRef] [Green Version]
- Allen, A.G.; Chung, C.H.; Atkins, A.; Dampier, W.; Khalili, K.; Nonnemacher, M.R.; Wigdahl, B. Gene Editing of HIV-1 Co-Receptors to Prevent and/or Cure Virus Infection. Front. Microbiol. 2018, 9, 2940. [Google Scholar] [CrossRef]
- Xiao, Q.; Guo, D.; Chen, S. Application of CRISPR/Cas9-Based Gene Editing in HIV-1/AIDS Therapy. Front. Cell. Infect. Microbiol. 2019, 9, 69. [Google Scholar] [CrossRef]
- Roehm, P.C.; Shekarabi, M.; Wollebo, H.S.; Bellizzi, A.; He, L.; Salkind, J.; Khalili, K. Inhibition of HSV-1 Replication by Gene Editing Strategy. Sci. Rep. 2016, 6, 23146. [Google Scholar] [CrossRef] [Green Version]
- Zhang, I.; Hsiao, Z.; Liu, F. Development of Genome Editing Approaches against Herpes Simplex Virus Infections. Viruses 2021, 13, 338. [Google Scholar] [CrossRef]
- Roizman, B.; Zhou, G. The 3 Facets of Regulation of Herpes Simplex Virus Gene Expression: A Critical Inquiry. Virology 2015, 479, 562–567. [Google Scholar] [CrossRef] [Green Version]
- van Diemen, F.R.; Kruse, E.M.; Hooykaas, M.J.G.; Bruggeling, C.E.; Schürch, A.C.; van Ham, P.M.; Imhof, S.M.; Nijhuis, M.; Wiertz, E.J.H.J.; Lebbink, R.J. CRISPR/Cas9-Mediated Genome Editing of Herpesviruses Limits Productive and Latent Infections. PLoS Pathog. 2016, 12, e1005701. [Google Scholar] [CrossRef] [Green Version]
- Karpov, D.S.; Karpov, V.L.; Klimova, R.R.; Demidova, N.A.; Kushch, A.A. A Plasmid-Expressed CRISPR/Cas9 System Suppresses Replication of HSV Type I in a Vero Cell Culture. Mol. Biol. 2019, 53, 70–78. [Google Scholar] [CrossRef]
- Park, S.E. Epidemiology, Virology, and Clinical Features of Severe Acute Respiratory Syndrome -Coronavirus-2 (SARS-CoV-2; Coronavirus Disease-19). Clin. Exp. Pediatr. 2020, 63, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Rothan, H.A.; Byrareddy, S.N. The Epidemiology and Pathogenesis of Coronavirus Disease (COVID-19) Outbreak. J. Autoimmun. 2020, 109, 102433. [Google Scholar] [CrossRef]
- Daikopoulou, V.; Apostolou, P.; Mourati, S.; Vlachou, I.; Gougousi, M.; Papasotiriou, I. Targeting Sars-Cov-2 Polymerase with New Nucleoside Analogues. Molecules 2021, 26, 3461. [Google Scholar] [CrossRef] [PubMed]
- Javalkote, V.S.; Kancharla, N.; Bhadra, B.; Shukla, M.; Soni, B.; Sapre, A.; Goodin, M.; Bandyopadhyay, A.; Dasgupta, S. CRISPR-Based Assays for Rapid Detection of SARS-CoV-2. Methods 2022, 203, 594–603. [Google Scholar] [CrossRef]
- Ullah, M.; Ibrar, M.; Uddin Khan, S.; Ullah, H.; Ali Khan, N. COVID-19 Detection: Comparison and Accuracy of Several Diagnostic Tests. Nov. Res. Microbiol. J. 2020, 4, 868–883. [Google Scholar]
- Yoshimi, K.; Takeshita, K.; Yamayoshi, S.; Shibumura, S.; Yamauchi, Y.; Yamamoto, M.; Yotsuyanagi, H.; Kawaoka, Y.; Mashimo, T. Rapid and Accurate Detection of Novel Coronavirus SARS-CoV-2 Using CRISPR-Cas3. medRxiv, 2020; preprint. [Google Scholar] [CrossRef]
- Ebrahimi, S.; Makvandi, M.; Abbasi, S.; Azadmanesh, K.; Teimoori, A. Developing Oncolytic Herpes Simplex Virus Type 1 through UL39 Knockout by CRISPR-Cas9. Iran. J. Basic Med. Sci. 2020, 23, 937–944. [Google Scholar] [CrossRef]
- Velásquez, A.C.; Castroverde, C.D.M.; He, S.Y. Plant-Pathogen Warfare under Changing Climate Conditions. Curr. Biol. 2018, 28, R619–R634. [Google Scholar] [CrossRef] [Green Version]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The Global Burden of Pathogens and Pests on Major Food Crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and Applications of CRISPR-Cas9 for Genome Engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef] [Green Version]
- Larson, M.H.; Gilbert, L.A.; Wang, X.; Lim, W.A.; Weissman, J.S.; Qi, L.S. CRISPR Interference (CRISPRi) for Sequence-Specific Control of Gene Expression. Nat. Protoc. 2013, 8, 2180–2196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidi, S.S.-A.; Mahas, A.; Vanderschuren, H.; Mahfouz, M.M. Engineering Crops of the Future: CRISPR Approaches to Develop Climate-Resilient and Disease-Resistant Plants. Genome Biol. 2020, 21, 289. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, M.; Mukhtar, S.; Sakina, A.; Dar, A.A.; Bhat, R.; Deshmukh, R.; Molla, K.; Kundoo, A.A.; Dar, M.S. Tweaking Genome-Editing Approaches for Virus Interference in Crop Plants. Plant Physiol. Biochem. 2020, 147, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Wei, X.; Sheng, Z.; Hu, P.; Tang, S. CRISPR/Cas9 for Development of Disease Resistance in Plants: Recent Progress, Limitations and Future Prospects. Brief. Funct. Genom. 2020, 19, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Baltes, N.J.; Hummel, A.W.; Konecna, E.; Cegan, R.; Bruns, A.N.; Bisaro, D.M.; Voytas, D.F. Conferring Resistance to Geminiviruses with the CRISPR–Cas Prokaryotic Immune System. Nat. Plants 2015, 1, 15145. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, J.N.; Ntui, V.O.; Ron, M.; Muiruri, S.K.; Britt, A.; Tripathi, L. CRISPR/Cas9 Editing of Endogenous Banana Streak Virus in the B Genome of Musa spp. Overcomes a Major Challenge in Banana Breeding. Commun. Biol. 2019, 2, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, A.A.; Sampson, T.R.; Ratner, H.K.; Grakoui, A.; Weiss, D.S. Cas9-Mediated Targeting of Viral RNA in Eukaryotic Cells. Proc. Natl. Acad. Sci. USA 2015, 112, 6164–6169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aman, R.; Ali, Z.; Butt, H.; Mahas, A.; Aljedaani, F.; Khan, M.Z.; Ding, S.; Mahfouz, M. RNA Virus Interference via CRISPR/Cas13a System in Plants. Genome Biol. 2018, 19, 1. [Google Scholar] [CrossRef]
- Zhang, T. Establishing RNA Virus Resistance in Plants by Harnessing CRISPR Immune System. Plant Biotechnol. J. 2018, 16, 1415–1423. [Google Scholar] [CrossRef] [Green Version]
- Ali, Z.; Eid, A.; Ali, S.; Mahfouz, M.M. Pea Early-Browning Virus-Mediated Genome Editing via the CRISPR/Cas9 System in Nicotiana Benthamiana and Arabidopsis. Virus Res. 2018, 244, 333–337. [Google Scholar] [CrossRef]
- Zhan, X.; Zhang, F.; Zhong, Z.; Chen, R.; Wang, Y.; Chang, L.; Bock, R.; Nie, B.; Zhang, J. Generation of Virus-Resistant Potato Plants by RNA Genome Targeting. Plant Biotechnol. J. 2019, 17, 1814–1822. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Meng, Z.; Abid, M.A.; Zhao, X. Novel Pollen Magnetofection System for Transformation of Cotton Plant with Magnetic Nanoparticles as Gene Carriers BT. In Transgenic Cotton: Methods and Protocols; Zhang, B., Ed.; Springer: New York, NY, USA, 2019; pp. 47–54. ISBN 978-1-4939-8952-2. [Google Scholar]
- Muhammad, S.; Zaidi, S.; Saud, A. Future of US-China Relations: Conflict, Competition or Cooperation? Asian Soc. Sci. 2020, 16, 1–14. [Google Scholar] [CrossRef]
- Hou, T.; Zeng, W.; Yang, M.; Chen, W.; Ren, L.; Ai, J.; Wu, J.; Liao, Y.; Gou, X.; Li, Y.; et al. Development and Evaluation of a Rapid CRISPR-Based Diagnostic for COVID-19. PLoS Pathog. 2020, 16, e1008705. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Hirano, J.; Murakami, K.; Hayashi, T. CRISPR-Cas9-Based Technology for Studying Enteric Virus Infection. Front. Genome Ed. 2022, 4, 888878. [Google Scholar] [CrossRef]
- Krishan, K.; Kanchan, T.; Singh, B.; Baryah, N.; Puri, S. Germline Editing: Editors Cautionary. Clin. Ter. 2018, 169, e58–e59. [Google Scholar] [CrossRef]
- Shinwari, Z.K.; Tanveer, F.; Khalil, A.T. Ethical Issues Regarding Crispr-Mediated Genome Editing. Curr. Issues Mol. Biol. 2018, 26, 103–110. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tripathi, S.; Khatri, P.; Fatima, Z.; Pandey, R.P.; Hameed, S. A Landscape of CRISPR/Cas Technique for Emerging Viral Disease Diagnostics and Therapeutics: Progress and Prospects. Pathogens 2023, 12, 56. https://doi.org/10.3390/pathogens12010056
Tripathi S, Khatri P, Fatima Z, Pandey RP, Hameed S. A Landscape of CRISPR/Cas Technique for Emerging Viral Disease Diagnostics and Therapeutics: Progress and Prospects. Pathogens. 2023; 12(1):56. https://doi.org/10.3390/pathogens12010056
Chicago/Turabian StyleTripathi, Shyam, Purnima Khatri, Zeeshan Fatima, Ramendra Pati Pandey, and Saif Hameed. 2023. "A Landscape of CRISPR/Cas Technique for Emerging Viral Disease Diagnostics and Therapeutics: Progress and Prospects" Pathogens 12, no. 1: 56. https://doi.org/10.3390/pathogens12010056
APA StyleTripathi, S., Khatri, P., Fatima, Z., Pandey, R. P., & Hameed, S. (2023). A Landscape of CRISPR/Cas Technique for Emerging Viral Disease Diagnostics and Therapeutics: Progress and Prospects. Pathogens, 12(1), 56. https://doi.org/10.3390/pathogens12010056