

Evolution and Current Status of Influenza A Virus in Chile: A Review

 , and
, and 
Abstract
:1. Introduction
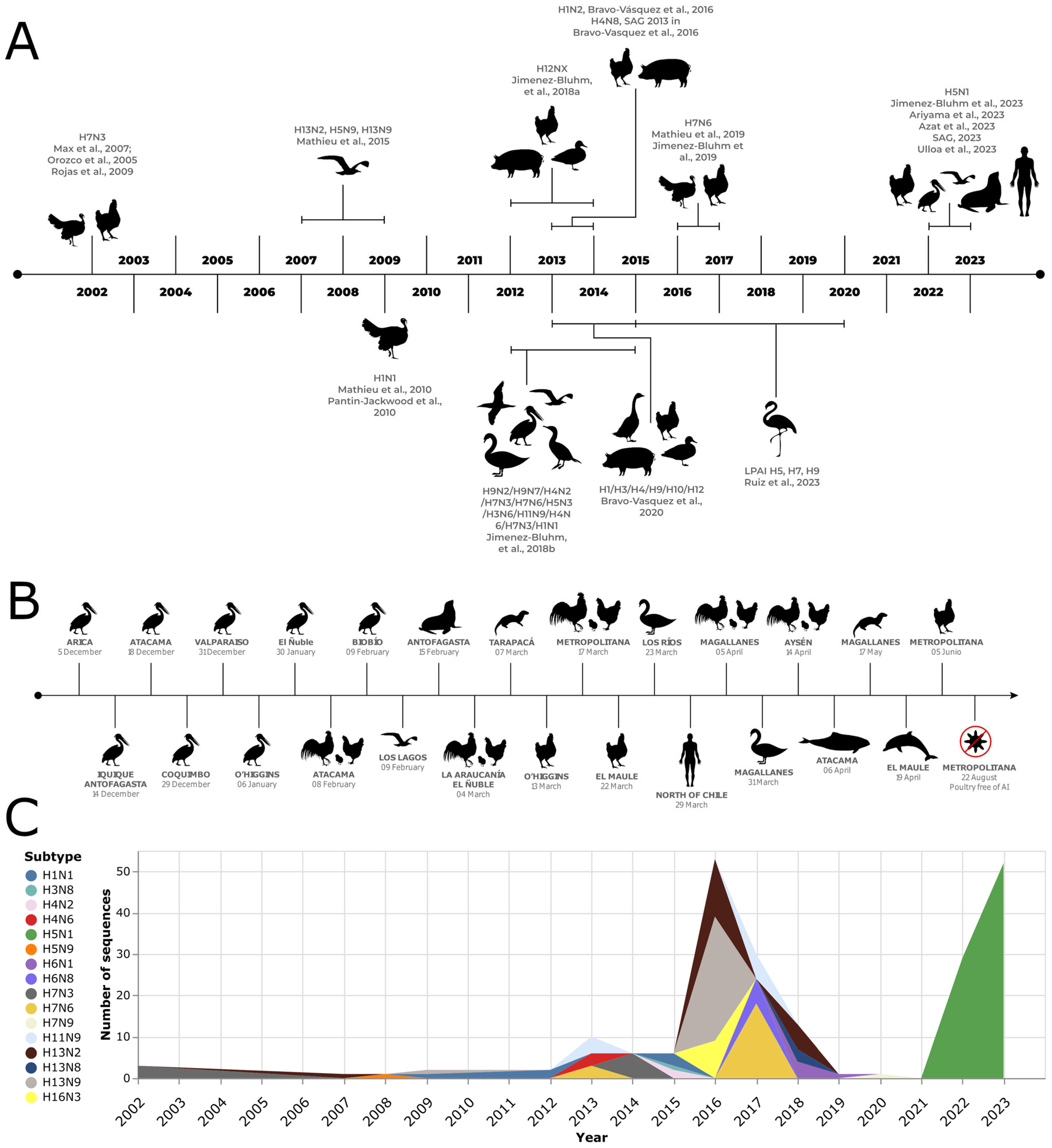
2. Influenza A Virus Subtypes in Commercial Poultry
2.1. AIV H7N3

2.2. Pandemic Influenza A (H1N1) 2009 (H1N1pdm09)
2.3. AIV H7N6
3. IAV Subtypes in Backyard Poultry and Swine
3.1. IAV H12NX
3.2. SIV H1N2
3.3. IAV H1/H3/H4/H9/H10/H12
4. Avian Influenza Virus Subtypes in Wild Birds
4.1. AIVs H13N2, H5N9, and H13N9
4.2. AIVs H9N2, H9N7, H4N2, H7N3, H7N6, H5N3, H3N6, H11N9, H4N6, H7N3, and H1N1
4.3. AIVs H9N2, H9N7, H3N8, H13N9, H13N8, H5N3, H13N2, H7N9
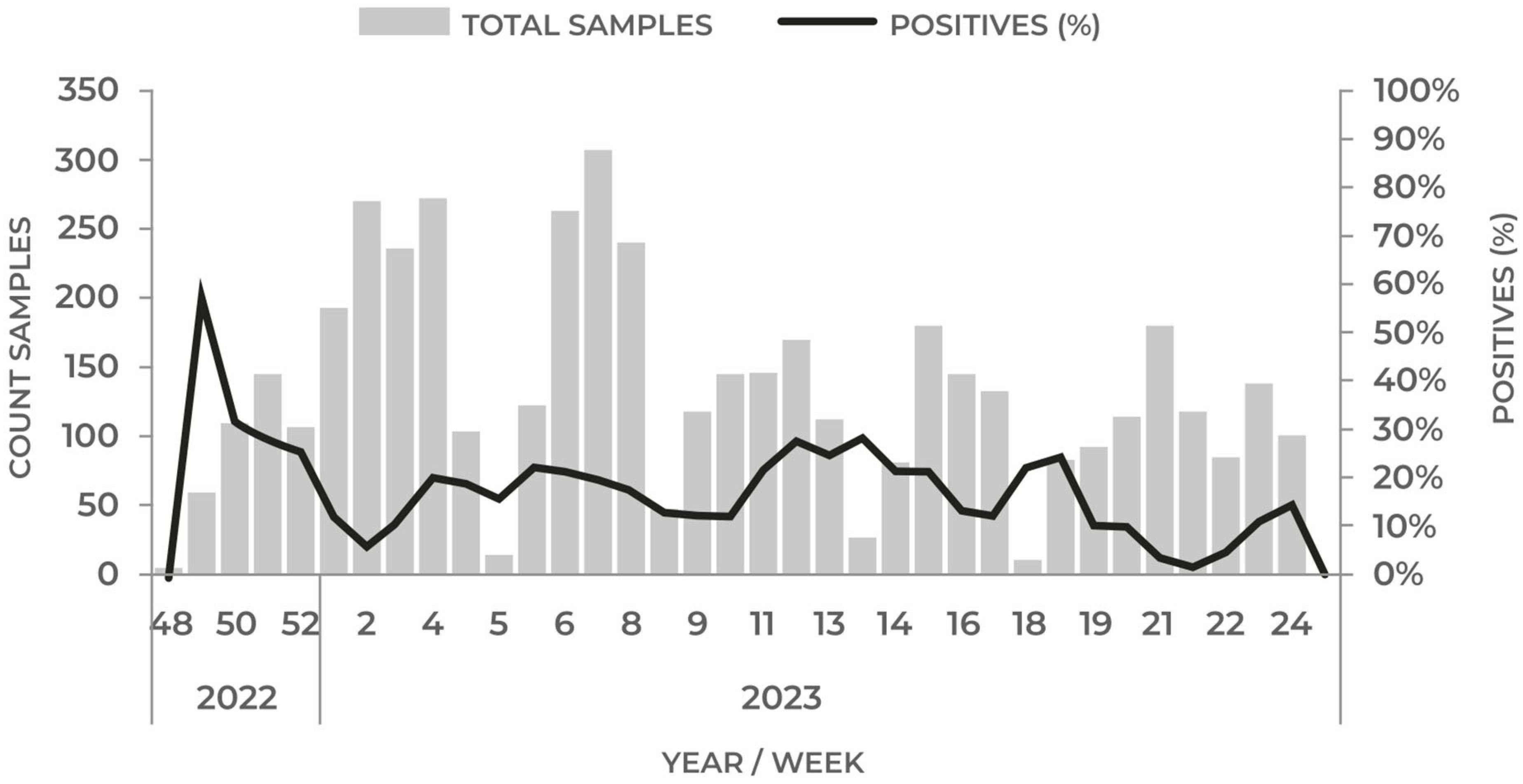
5. Highly Pathogenic Avian Influenza (HPAI) H5N1
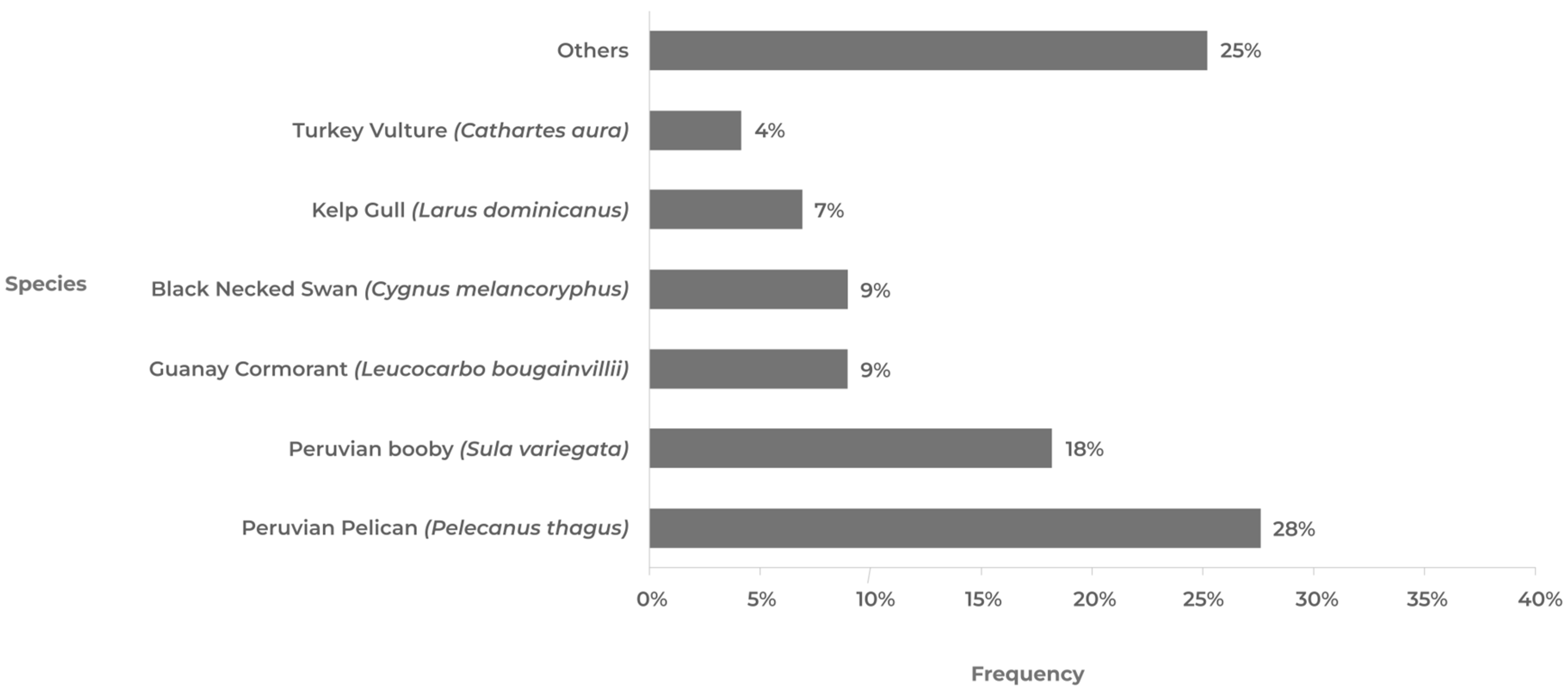
5.1. AIV H5N1 in Wild Birds

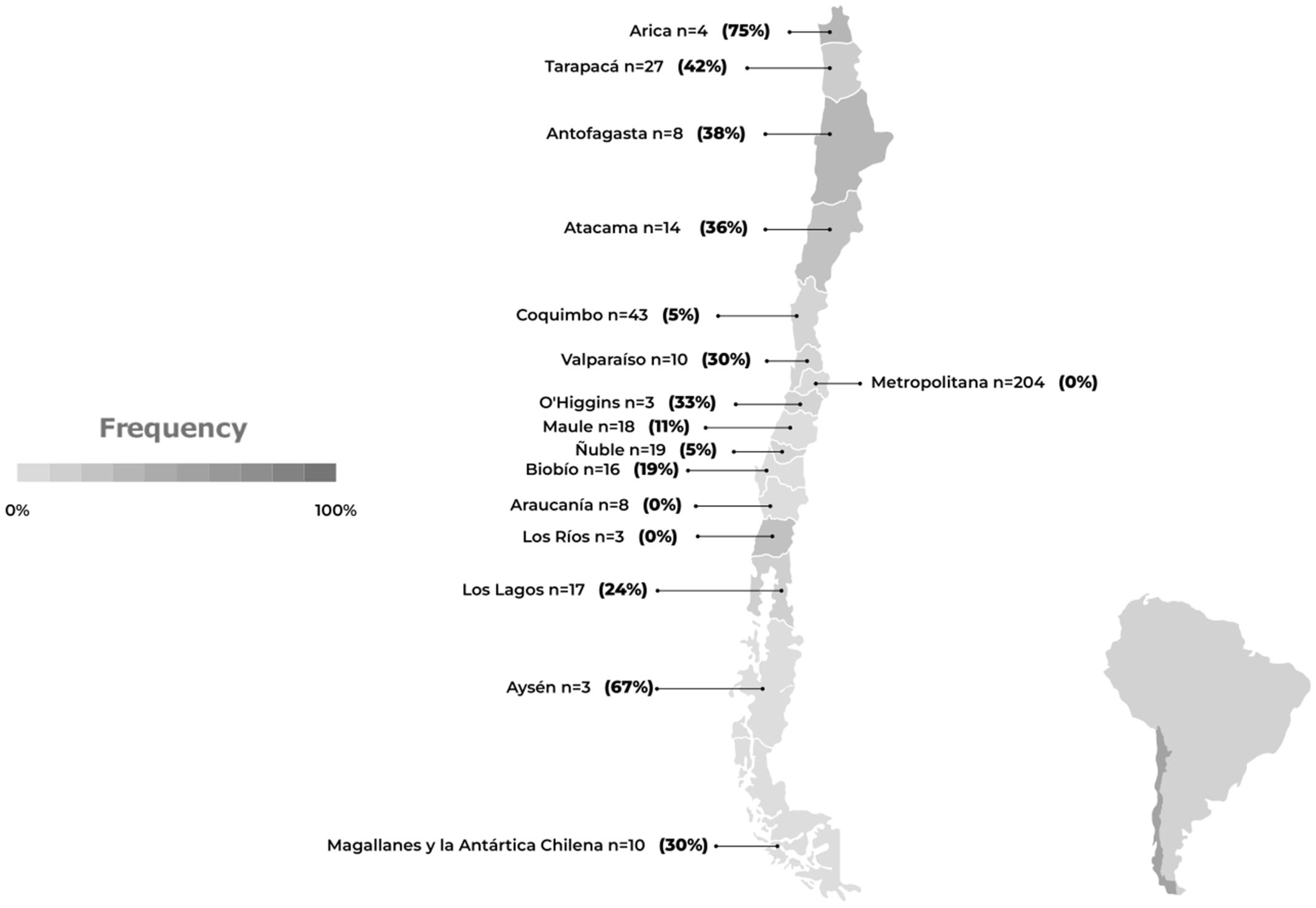
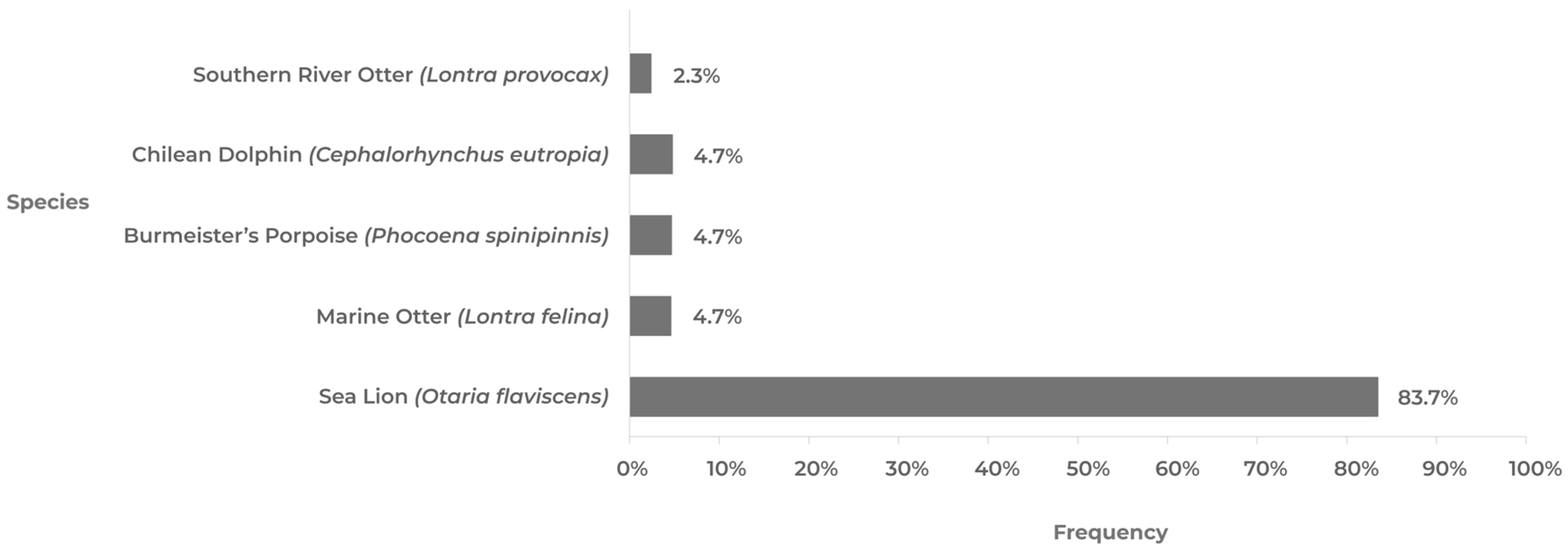
5.2. AIV H5N1 in Marine Mammals
5.3. AIV H5N1 in Humans
6. Conclusions
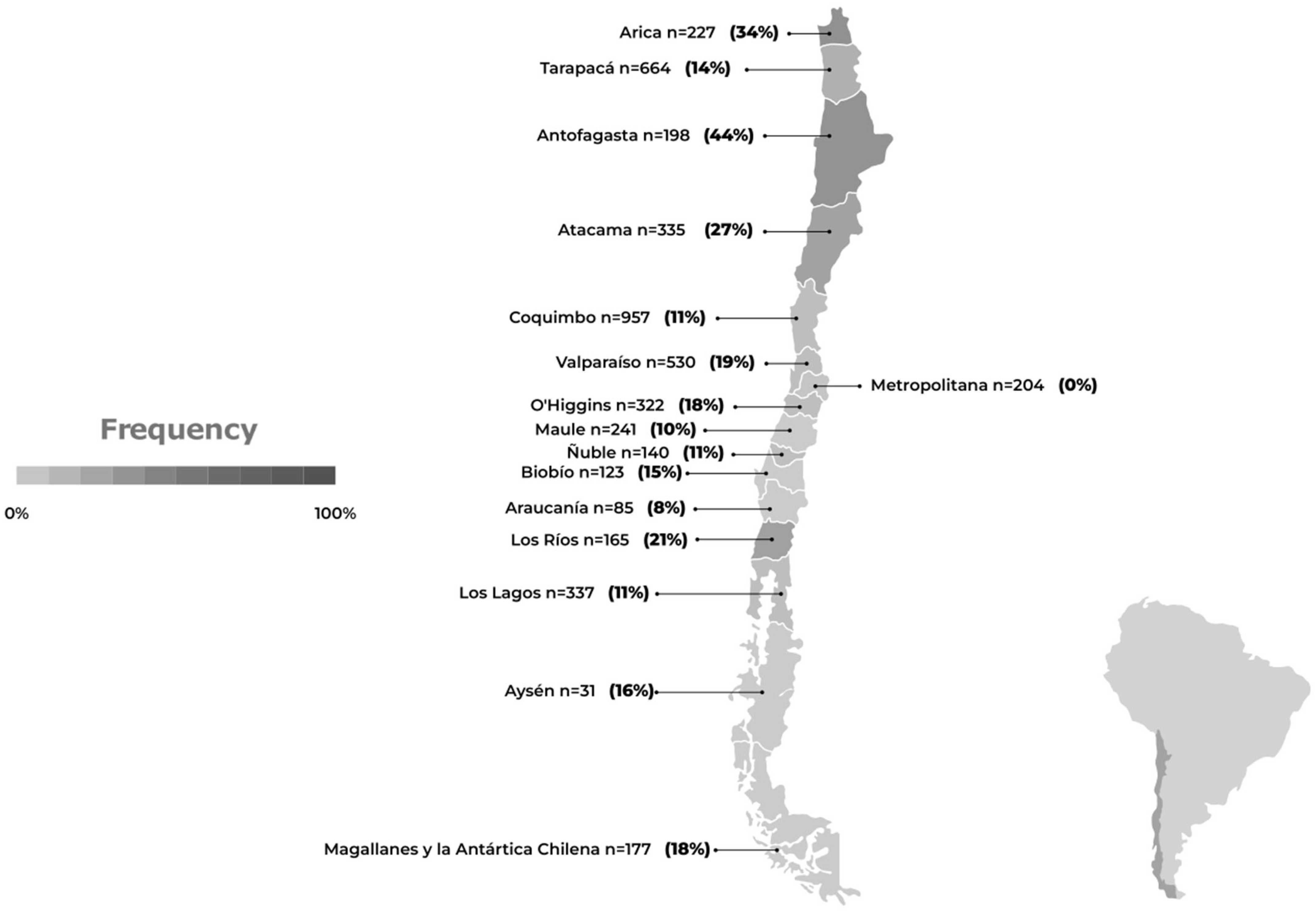
- Proactive surveillance of migratory birds: As we anticipate upcoming migratory seasons, it is essential to implement a proactive strategy encompassing enhanced detection and genomic surveillance, to ensure the effective control and monitoring of wild birds. Continued monitoring of critical locations along migratory routes, such as the Lluta River wetland in Northern Chile, remains crucial to early detection and intervention.
- Comprehensive stranding control: Addressing avian influenza also necessitates a holistic approach including not only the control of stranding events but also the systematic detection and subtyping of AIVs in the affected marine mammals and bird populations. This multifaceted approach can provide valuable insights into the diversity and dynamics of the virus.
- Expanded backyard poultry surveillance: Given the evident role of wild birds and marine mammals in virus dissemination, it is imperative to expand surveillance efforts for avian influenza in backyard poultry across various regions of the country. This broader approach should encompass areas influenced by both migratory birds and marine mammals, recognizing their potential as secondary vectors.
- Coordinated genomic surveillance for one health: Effective risk mitigation necessitates a coordinated One Health approach that integrates the genomic surveillance of avian influenza, addressing both human and animal health concerns. This approach should be underpinned by robust communication channels facilitating collaboration among diverse stakeholders from the public and private sectors involved in managing the disease across different compartments.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Koonin, E.V.; Dolja, V.V.; Krupovic, M.; Varsani, A.; Wolf, Y.I.; Yutin, N.; Zerbini, F.M.; Kuhn, J.H. Create a Megataxonomic Framework, Filling All Principal Taxonomic Ranks, for Realm Riboviria; ICTV: Montolivet, France, 2019; Unpublished. [Google Scholar] [CrossRef]
- Turnbull, O.M.H.; Ortiz-Baez, A.S.; Eden, J.-S.; Shi, M.; Williamson, J.E.; Gaston, T.F.; Zhang, Y.-Z.; Holmes, E.C.; Geoghegan, J.L. Meta-Transcriptomic Identification of Divergent Amnoonviridae in Fish. Viruses 2020, 12, 1254. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.; Mcveigh, R.; O’neill, K.; Robbertse, B.; et al. NCBI Taxonomy: A comprehensive update on curation, resources and tools. Database 2020, 2020, baaa062. [Google Scholar] [CrossRef] [PubMed]
- Parry, R.; Wille, M.; Turnbull, O.M.H.; Geoghegan, J.L.; Holmes, E.C. Divergent Influenza-Like Viruses of Amphibians and Fish Support an Ancient Evolutionary Association. Viruses 2020, 12, 1042. [Google Scholar] [CrossRef]
- García-Sastre, A. Create One New Genus (Mykissvirus) Including One New Species (Mykissvirus tructae) (Articulavirales: Orthomyxoviridae). 2021. Available online: https://ictv.global/ictv/proposals/2021.022M.R.Orthomyxoviridae_1ngen_1nsp_Mykiss.zip (accessed on 5 June 2023).
- Mohr, P.G.; Godwin, S.E.; Morrison, R.N.; Carson, J.; Crane, M.S.t.J.; Moody, N.J.G. Create One New Genus (Sardinovirus) Including One New Species (Sardinovirus pilchardi) (Articulavirales: Orthomyxoviridae). 2021. Available online: https://ictv.global/ictv/proposals/2021.023M.R.Orthomyxoviridae_1ngen_1nsp_Sardino.zip (accessed on 2 June 2023).
- Petrone, M.E.; Parry, R.; Mifsud, J.C.O.; Van Brussel, K.; Vorhees, I.; Richards, Z.T.; Holmes, E.C. Evidence for an Aquatic Origin of Influenza Virus and the order Articulavirales. bioRxiv 2023. preprint. [Google Scholar] [CrossRef]
- Wille, M.; Holmes, E.C. The Ecology and Evolution of Influenza Viruses. Cold Spring Harb. Perspect. Med. 2019, 10, a038489. [Google Scholar] [CrossRef]
- Fouchier, R.A.M.; Munster, V.; Wallensten, A.; Bestebroer, T.M.; Herfst, S.; Smith, D.; Rimmelzwaan, G.F.; Olsen, B.; Osterhaus, A.D.M.E. Characterization of a Novel Influenza A Virus Hemagglutinin Subtype (H16) Obtained from Black-Headed Gulls. J. Virol. 2005, 79, 2814–2822. [Google Scholar] [CrossRef]
- Short, K.R.; Richard, M.; Verhagen, J.H.; van Riel, D.; Schrauwen, E.J.; Brand, J.M.v.D.; Mänz, B.; Bodewes, R.; Herfst, S. One health, multiple challenges: The inter-species transmission of influenza A virus. One Health 2015, 1, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Kahn, R.E.; Richt, J.A. The pig as a mixing vessel for influenza viruses: Human and veterinary implications. J. Mol. Genet. Med. 2008, 3, 158–166. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2702078/ (accessed on 5 June 2023). [CrossRef]
- Yang, W.; Schountz, T.; Ma, W. Bat Influenza Viruses: Current Status and Perspective. Viruses 2021, 13, 547. [Google Scholar] [CrossRef]
- Uribe, M.; Rodríguez-Posada, M.E.; Ramirez-Nieto, G.C. Molecular Evidence of Orthomyxovirus Presence in Colombian Neotropical Bats. Front. Microbiol. 2022, 13, 845546. [Google Scholar] [CrossRef] [PubMed]
- Kessler, S.; Harder, T.C.; Schwemmle, M.; Ciminski, K. Influenza A Viruses and Zoonotic Events—Are We Creating Our Own Reservoirs? Viruses 2021, 13, 2250. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Types of Influenza Viruses. 2023. Available online: https://www.cdc.gov/flu/about/viruses/types.html (accessed on 5 June 2023).
- Olsen, B.; Munster, V.J.; Wallensten, A.; Waldenström, J.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Global Patterns of Influenza A Virus in Wild Birds. Science 2006, 312, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Kandeil, A.; Patton, C.; Jones, J.C.; Jeevan, T.; Harrington, W.N.; Trifkovic, S.; Seiler, J.P.; Fabrizio, T.; Woodard, K.; Turner, J.C.; et al. Rapid evolution of A(H5N1) influenza viruses after intercontinental spread to North America. Nat. Commun. 2023, 14, 3082. [Google Scholar] [CrossRef]
- Gamarra-Toledo, V.; Plaza, P.I.; Gutiérrez, R.; Inga-Diaz, G.; Saravia-Guevara, P.; Pereyra-Meza, O.; Coronado-Flores, E.; Calderón-Cerrón, A.; Quiroz-Jiménez, G.; Martinez, P.; et al. Mass Mortality of Marine Mammals Associated to Highly Pathogenic Influenza Virus (H5N1) in South America. bioRxiv 2023. preprint. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Human Infection with Highly Pathogenic Avian Influenza A(H5N1) Virus in Chile. 2023. Available online: https://www.cdc.gov/flu/avianflu/spotlights/2022-2023/chile-first-case-h5n1-addendum.htm (accessed on 30 May 2023).
- World Animal Health Information System. High Pathogenicity Avian Influenza (HPAI)—Situation Report. 2023. Available online: https://www.woah.org/en/document/high-pathogenicity-avian-influenza-hpai-situation-report-47/ (accessed on 7 September 2023).
- Orozco, C. The SPS Agreement and Crisis Management: The Chile–EU Avian Influenza Experience. Managing the Challenges of WTO Participation; Cambridge University Press: Cambridge, UK, 2005; pp. 150–166. [Google Scholar] [CrossRef]
- Max, V.; Herrera, J.; Moreira, R.; Rojas, H. Avian Influenza in Chile: A Successful Experience. Avian Dis. 2007, 51, 363–365. [Google Scholar] [CrossRef]
- Suarez, D.L.; Senne, D.A.; Banks, J.; Brown, I.H.; Essen, S.C.; Lee, C.-W.; Manvell, R.J.; Mathieu-Benson, C.; Moreno, V.; Pedersen, J.C.; et al. Recombination Resulting in Virulence Shift in Avian Influenza Outbreak, Chile. Emerg. Infect. Dis. 2004, 10, 693–699. [Google Scholar] [CrossRef]
- Rojas Olavarría, H.; Moreira Zúñiga, R.; Mundial, B. Influenza Aviar en Chile 2002: Una Sinopsis; Instituto Interamericano de Cooperación para la Agricultura (IICA)—Banco Mundial (BM): Santiago, Chile, 2009; 32p. [Google Scholar]
- Pantin-Jackwood, M.; Wasilenko, J.L.; Spackman, E.; Suarez, D.L.; Swayne, D.E. Susceptibility of turkeys to pandemic-H1N1 virus by reproductive tract insemination. Virol. J. 2010, 7, 27. [Google Scholar] [CrossRef]
- Mathieu, C.; Moreno, V.; Retamal, P.; Gonzalez, A.; Rivera, A.; Fuller, J.; Jara, C.; Lecocq, C.; Rojas, M.; García, A.; et al. Pandemic (H1N1) 2009 in Breeding Turkeys, Valparaiso, Chile. Emerg. Infect. Dis. 2010, 16, 709–711. [Google Scholar] [CrossRef]
- Mathieu, C.; Gonzalez, A.; Garcia, A.; Johow, M.; Badia, C.; Jara, C.; Nuñez, P.; Neira, V.; Montiel, N.A.; Killian, M.L.; et al. H7N6 low pathogenic avian influenza outbreak in commercial turkey farms in Chile caused by a native South American Lineage. Transbound. Emerg. Dis. 2019, 68, 2–12. [Google Scholar] [CrossRef]
- Jimenez-Bluhm, P.; Bravo-Vasquez, N.; Torchetti, M.K.; Killian, M.L.; Livingston, B.; Herrera, J.; Fuentes, M.; Schultz-Cherry, S.; Hamilton-West, C. Low pathogenic avian influenza (H7N6) virus causing an outbreak in commercial Turkey farms in Chile. Emerg. Microbes Infect. 2019, 8, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Bluhm, P.; Di Pillo, F.; Bahl, J.; Osorio, J.; Schultz-Cherry, S.; Hamilton-West, C. Circulation of influenza in backyard productive systems in central Chile and evidence of spillover from wild birds. Prev. Vet. Med. 2018, 153, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Vasquez, N.; Di Pillo, F.; Lazo, A.; Jiménez-Bluhm, P.; Schultz-Cherry, S.; Hamilton-West, C. Presence of influenza viruses in backyard poultry and swine in El Yali wetland, Chile. Prev. Vet. Med. 2016, 134, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Vasquez, N.; Baumberger, C.; Jimenez-Bluhm, P.; Di Pillo, F.; Lazo, A.; Sanhueza, J.; Schultz-Cherry, S.; Hamilton-West, C. Risk factors and spatial relative risk assessment for influenza A virus in poultry and swine in backyard production systems of central Chile. Vet. Med. Sci. 2020, 6, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, C.; Moreno, V.; Pedersen, J.; Jeria, J.; Agredo, M.; Gutiérrez, C.; García, A.; Vásquez, M.; Avalos, P.; Retamal, P. Avian Influenza in wild birds from Chile, 2007–2009. Virus Res. 2015, 199, 42–45. [Google Scholar] [CrossRef]
- Jiménez-Bluhm, P.; Karlsson, E.A.; Freiden, P.; Sharp, B.; Di Pillo, F.; Osorio, J.E.; Hamilton-West, C.; Schultz-Cherry, S. Wild birds in Chile Harbor diverse avian influenza A viruses. Emerg. Microbes Infect. 2018, 7, 1–4. [Google Scholar] [CrossRef]
- Ruiz, S.; Galdames, P.; Baumberger, C.; Gonzalez, M.A.; Rojas, C.; Oyarzun, C.; Orozco, K.; Mattar, C.; Freiden, P.; Sharp, B.; et al. Remote Sensing and Ecological Variables Related to Influenza A Prevalence and Subtype Diversity in Wild Birds in the Lluta Wetland of Northern Chile. Viruses 2023, 15, 1241. [Google Scholar] [CrossRef]
- Ariyama, N.; Pardo Roa, C.; Munoz, G.; Aguayo, C.; Avila, C.; Mathieu, C.; Brito, B.; Medina, R.; Johow, M.; Neira-Ramirez, V. Emergence and rapid dissemination of highly pathogenic avian influenza virus H5N1 clade 2.3.4.4b in wild birds, Chile. bioRxiv 2023. preprint. [Google Scholar] [CrossRef]
- SAG (Agricultural and Livestock Service). Servicio Agrícola y Ganadero, Ministerio de Agricultura de Chile. Plataforma de Influenza Aviar. 2023. Available online: https://www.sag.gob.cl/ia (accessed on 25 May 2023).
- Jimenez-Bluhm, P.; Siegers, J.Y.; Tan, S.; Sharp, B.; Freiden, P.; Johow, M.; Orozco, K.; Ruiz, S.; Baumberger, C.; Galdames, P.; et al. Detection and phylogenetic analysis of highly pathogenic A/H5N1 avian influenza clade 2.3.4.4b virus in Chile, 2022. Emerg. Microbes Infect. 2023, 12, 2220569. [Google Scholar] [CrossRef]
- Azat, C.; Alvarado-Rybak, M.; Aguilera, J.F.; Benavides, J.A. Spatio-Temporal Dynamics and Drivers of Highly Pathogenic Avian Influenza H5N1 in Chile. bioRxiv 2023. preprint. [Google Scholar] [CrossRef]
- Ulloa, M.; Fernández, A.; Ariyama, N.; Colom-Rivero, A.; Rivera, C.; Nuñez, P.; Sanhueza, P.; Johow, M.; Araya, H.; Torres, J.C.; et al. Mass Mortality Event in South American Sea Lions (Otaria flavescens) Correlated to Highly Pathogenic Avian Influenza (HPAI) H5N1 Outbreak in Chile. In Veterinary Quarterly; Informa UK Limited: London, UK, 2023; pp. 1–13. [Google Scholar] [CrossRef]
- SERNAPESCA. Servicio Nacional de Pesca. Plataforma de Influenza Aviar. 2023. Available online: http://www.sernapesca.cl/influenza-aviar (accessed on 7 September 2023).
- Nucleotide. Bethesda (MD): National Library of Medicine (US), National Center for Biotechnology Information. 2004. Available online: https://www.ncbi.nlm.nih.gov/nuccore (accessed on 7 September 2023).
- Spackman, E.; McCracken, K.G.; Winker, K.; Swayne, D.E. H7N3 Avian Influenza Virus Found in a South American Wild Duck Is Related to the Chilean 2002 Poultry Outbreak, Contains Genes from Equine and North American Wild Bird Lineages, and Is Adapted to Domestic Turkeys. J. Virol. 2006, 80, 7760–7764. [Google Scholar] [CrossRef] [PubMed]
- Kibenge, F.S.B. A One Health approach to mitigate the impact of influenza A virus (IAV) reverse zoonosis is by vaccinating humans and susceptible farmed and pet animals. Am. J. Vet. Res. 2023, 84, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Trifonov, V.; Khiabanian, H.; Rabadan, R. Geographic Dependence, Surveillance, and Origins of the 2009 Influenza A (H1N1) Virus. N. Engl. J. Med. 2009, 361, 115–119. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Manual for the Laboratory Diagnosis and Virological Surveillance of Influenza; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Gilbertson, B.; Subbarao, K. Mammalian infections with highly pathogenic avian influenza viruses renew concerns of pandemic potential. J. Exp. Med. 2023, 220, e20230447. [Google Scholar] [CrossRef] [PubMed]
- Chmielewski, R.; Swayne, D.E. Avian Influenza: Public Health and Food Safety Concerns. Annu. Rev. Food Sci. Technol. 2011, 2, 37–57. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control; European Union Reference Laboratory for Avian Influenza; Adlhoch, C.; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Marangon, S.; Mirinaviciute, G.; Niqueux, É.; et al. Avian influenza overview December 2022–March 2023. EFSA J. 2023, 21, e07917. [Google Scholar] [CrossRef]
- Sagong, M.; Lee, Y.; Song, S.; Cha, R.M.; Lee, E.; Kang, Y.; Cho, H.; Kang, H.; Lee, Y.; Lee, K. Emergence of clade 2.3.4.4b novel reassortant H5N1 high pathogenicity avian influenza virus in South Korea during late 2021. Transbound. Emerg. Dis. 2022, 69, E3255–E3260. [Google Scholar] [CrossRef]
- El-Shesheny, R.; Moatasim, Y.; Mahmoud, S.H.; Song, Y.; El Taweel, A.; Gomaa, M.; Kamel, M.N.; El Sayes, M.; Kandeil, A.; Lam, T.T.Y.; et al. Highly Pathogenic Avian Influenza A(H5N1) Virus Clade 2.3.4.4b in Wild Birds and Live Bird Markets, Egypt. Pathogens 2022, 12, 36. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control; European Union Reference Laboratory for Avian Influenza; Adlhoch, C.; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Marangon, S.; Niqueux, É.; Staubach, C.; et al. Avian influenza overview June–September 2022. EFSA J. 2022, 20, e07597. [Google Scholar] [CrossRef]
- Harvey, J.A.; Mullinax, J.M.; Runge, M.C.; Prosser, D.J. The changing dynamics of highly pathogenic avian influenza H5N1: Next steps for management & science in North America. Biol. Conserv. 2023, 282, 110041. [Google Scholar] [CrossRef]
- Günther, A.; Krone, O.; Svansson, V.; Pohlmann, A.; King, J.; Hallgrimsson, G.T.; Skarphéðinsson, K.H.; Sigurðardóttir, H.; Jónsson, S.R.; Beer, M.; et al. Iceland as Stepping Stone for Spread of Highly Pathogenic Avian Influenza Virus between Europe and North America. Emerg. Infect. Dis. 2022, 28, 2383–2388. [Google Scholar] [CrossRef] [PubMed]
- Leguia, M.; Garcia-Glaessner, A.; Muñoz-Saavedra, B.; Juarez, D.; Barrera, P.; Calvo-Mac, C.; Jara, J.; Silva, W.; Ploog, K.; Amaro, L.; et al. Highly pathogenic avian influenza A (H5N1) in marine mammals and seabirds in Peru. Nat. Commun. 2023, 14, 5489. [Google Scholar] [CrossRef] [PubMed]
- eBird. eBird: An Online Database of Bird Distribution and Abundance; eBird; Cornell Lab of Ornithology: Ithaca, NY, USA, 2021; Available online: http://www.ebird.org (accessed on 30 May 2023).
- Reperant, L.A.; Kuiken, T.; Osterhaus, A.D. Adaptive pathways of zoonotic influenza viruses: From exposure to establishment in humans. Vaccine 2012, 30, 4419–4434. [Google Scholar] [CrossRef] [PubMed]
- Puryear, W.; Sawatzki, K.; Hill, N.; Foss, A.; Stone, J.J.; Doughty, L.; Walk, D.; Gilbert, K.; Murray, M.; Cox, E.; et al. Highly Pathogenic Avian Influenza A(H5N1) Virus Outbreak in New England Seals, United States. Emerg. Infect. Dis. 2023, 29, 786–791. [Google Scholar] [CrossRef]
- Agüero, M.; Monne, I.; Sánchez, A.; Zecchin, B.; Fusaro, A.; Ruano, M.J.; del Valle Arrojo, M.; Fernández-Antonio, R.; Souto, A.M.; Tordable, P.; et al. Highly pathogenic avian influenza A(H5N1) virus infection in farmed minks, Spain, October 2022. Euro Surveill. 2023, 28, 2300001. [Google Scholar] [CrossRef]
- Bowen, W.D.; Harrison, G.D. Comparison of harbour seal diets in two inshore habitats of Atlantic Canada. Can. J. Zool. 1996, 74, 125–135. [Google Scholar] [CrossRef]
- World Health Organization (WHO): Human Infection Caused by Avian Influenza A (H5N1)—Chile. Available online: https://www.who.int/emergencies/disease-outbreak-news/item/2023-DON461 (accessed on 5 June 2023).
- Pardo-Roa, C.; Nelson, M.I.; Ariyama, N.; Aguayo, C.; Almonacid, L.I.; Munoz, G.; Navarro, C.; Avila, C.; Ulloa, M.; Reyes, R.; et al. Cross-Species Transmission and PB2 Mammalian Adaptations of Highly Pathogenic Avian Influenza A/H5N1 Viruses in Chile. bioRxiv 2023. preprint. [Google Scholar] [CrossRef]
- Castillo, A.; Fasce, R.; Parra, B.; Andrade, W.; Covarrubias, P.; Hueche, A.; Campano, C.; Tambley, C.; Rojas, M.; Araya, M.; et al. The first case of human infection with H5N1 avian Influenza A virus in Chile. J. Travel Med. 2023, 30, taad083. [Google Scholar] [CrossRef]
- Vreman, S.; Kik, M.; Germeraad, E.; Heutink, R.; Harders, F.; Spierenburg, M.; Engelsma, M.; Rijks, J.; Brand, J.v.D.; Beerens, N. Zoonotic Mutation of Highly Pathogenic Avian Influenza H5N1 Virus Identified in the Brain of Multiple Wild Carnivore Species. Pathogens 2023, 12, 168. [Google Scholar] [CrossRef]
- Na Ayudhya, S.S.; Kuiken, T. Reverse Zoonosis of COVID-19: Lessons From the 2009 Influenza Pandemic. Vet. Pathol. 2020, 58, 234–242. [Google Scholar] [CrossRef]
- Evans, R.; Bommineni, Y.; Falk, J.; Blackway, A.; Young, B.; Isenhart, C. Mature Turkey Breeder Hens Exposed to Pandemic Influenza H1N1: Resultant Effects on Morbidity, Mortality, and Fecundity. Avian Dis. 2015, 59, 171–174. [Google Scholar] [CrossRef] [PubMed]
- González-Acuña, D.; Llanos-Soto, S. Una revisión sistemática de los patógenos virales y bacterianos de aves silvestres en Chile. In Revista Chilena de Infectología. SciELO Agencia Nacional de Investigacion y Desarrollo (ANID); SciELO: Manaus, Brazil, 2020; Volume 37, pp. 422–442. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Peruvian Pelican (Pelecanus thagus) | Peruvian Booby (Sula variegata) | Guanay Cormorant (Leucocarbo bougainvillii) | Kelp Gull (Larus dominicanus) | Black-Necked Swan (Cygnus melancoryphus) | Turkey Vulture (Cathartes aura) | Others | Total |
|---|---|---|---|---|---|---|---|---|
| Arica | 7 | 18 | 12 | 4 | 0 | 2 | 34 | 77 |
| Tarapacá | 8 | 19 | 29 | 4 | 0 | 8 | 23 | 91 |
| Antofagasta | 31 | 23 | 2 | 0 | 0 | 13 | 16 | 85 |
| Atacama | 41 | 24 | 7 | 6 | 0 | 2 | 10 | 90 |
| Coquimbo | 21 | 33 | 12 | 14 | 0 | 2 | 22 | 104 |
| Valparaíso | 62 | 13 | 3 | 3 | 1 | 3 | 16 | 101 |
| O’Higgins | 21 | 8 | 5 | 6 | 4 | 3 | 11 | 58 |
| Maule | 9 | 3 | 0 | 3 | 0 | 0 | 9 | 24 |
| Ñuble | 10 | 0 | 0 | 6 | 0 | 0 | 0 | 16 |
| Bío Bío | 5 | 1 | 0 | 6 | 0 | 0 | 6 | 18 |
| Araucanía | 0 | 0 | 0 | 0 | 3 | 0 | 4 | 7 |
| Los Ríos | 0 | 0 | 0 | 0 | 30 | 0 | 5 | 35 |
| Los Lagos | 1 | 0 | 0 | 1 | 22 | 0 | 12 | 36 |
| Aysen | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 5 |
| Magallanes | 0 | 0 | 0 | 1 | 10 | 0 | 20 | 31 |
| Total | 216 | 142 | 70 | 54 | 70 | 33 | 193 | 778 |
| Region | Marine Otter (Lontra felina) | Chilean Dolphin (Cephalorhynchus eutropia) | Southern River Otter (Lontra provocax) | Sea Lion (Otaria flavescens) | Burmeister’s Porpoise (Phocoena spinipinnis) | Total |
|---|---|---|---|---|---|---|
| Arica | 1 | 0 | 0 | 2 | 0 | 3 |
| Tarapacá | 1 | 0 | 0 | 10 | 0 | 11 |
| Antofagasta | 0 | 0 | 0 | 2 | 1 | 3 |
| Atacama | 0 | 0 | 0 | 4 | 1 | 5 |
| Coquimbo | 0 | 0 | 0 | 2 | 0 | 2 |
| Valparaíso | 0 | 0 | 0 | 3 | 0 | 3 |
| O’Higgins | 0 | 0 | 0 | 1 | 0 | 1 |
| Maule | 0 | 1 | 0 | 1 | 0 | 2 |
| Ñuble | 0 | 1 | 0 | 0 | 0 | 1 |
| Bío Bío | 0 | 0 | 0 | 3 | 0 | 3 |
| Araucanía | 0 | 0 | 0 | 0 | 0 | 0 |
| Los Ríos | 0 | 0 | 0 | 0 | 0 | 0 |
| Los Lagos | 0 | 0 | 0 | 4 | 0 | 4 |
| Aysén | 0 | 0 | 0 | 2 | 0 | 2 |
| Magallanes | 0 | 0 | 1 | 2 | 0 | 3 |
| Total | 2 | 2 | 1 | 36 | 2 | 43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Godoy, M.; Oca, M.M.d.; Caro, D.; Pontigo, J.P.; Kibenge, M.; Kibenge, F. Evolution and Current Status of Influenza A Virus in Chile: A Review. Pathogens 2023, 12, 1252. https://doi.org/10.3390/pathogens12101252
Godoy M, Oca MMd, Caro D, Pontigo JP, Kibenge M, Kibenge F. Evolution and Current Status of Influenza A Virus in Chile: A Review. Pathogens. 2023; 12(10):1252. https://doi.org/10.3390/pathogens12101252
Chicago/Turabian StyleGodoy, Marcos, Marco Montes de Oca, Diego Caro, Juan Pablo Pontigo, Molly Kibenge, and Frederick Kibenge. 2023. "Evolution and Current Status of Influenza A Virus in Chile: A Review" Pathogens 12, no. 10: 1252. https://doi.org/10.3390/pathogens12101252
APA StyleGodoy, M., Oca, M. M. d., Caro, D., Pontigo, J. P., Kibenge, M., & Kibenge, F. (2023). Evolution and Current Status of Influenza A Virus in Chile: A Review. Pathogens, 12(10), 1252. https://doi.org/10.3390/pathogens12101252