


Acquired blaVIM and blaGES Carbapenemase-Encoding Genes in Pseudomonas aeruginosa: A Seven-Year Survey Highlighting an Increasing Epidemiological Threat

 , and
, and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Clinical Strains, Identification, and Antimicrobial Susceptibility Profiles
2.2. Genotypic Detection of Carbapenemases
2.3. Sequencing
2.4. Phenotypic Detection of Carbapenemases
2.5. Statistical Analysis
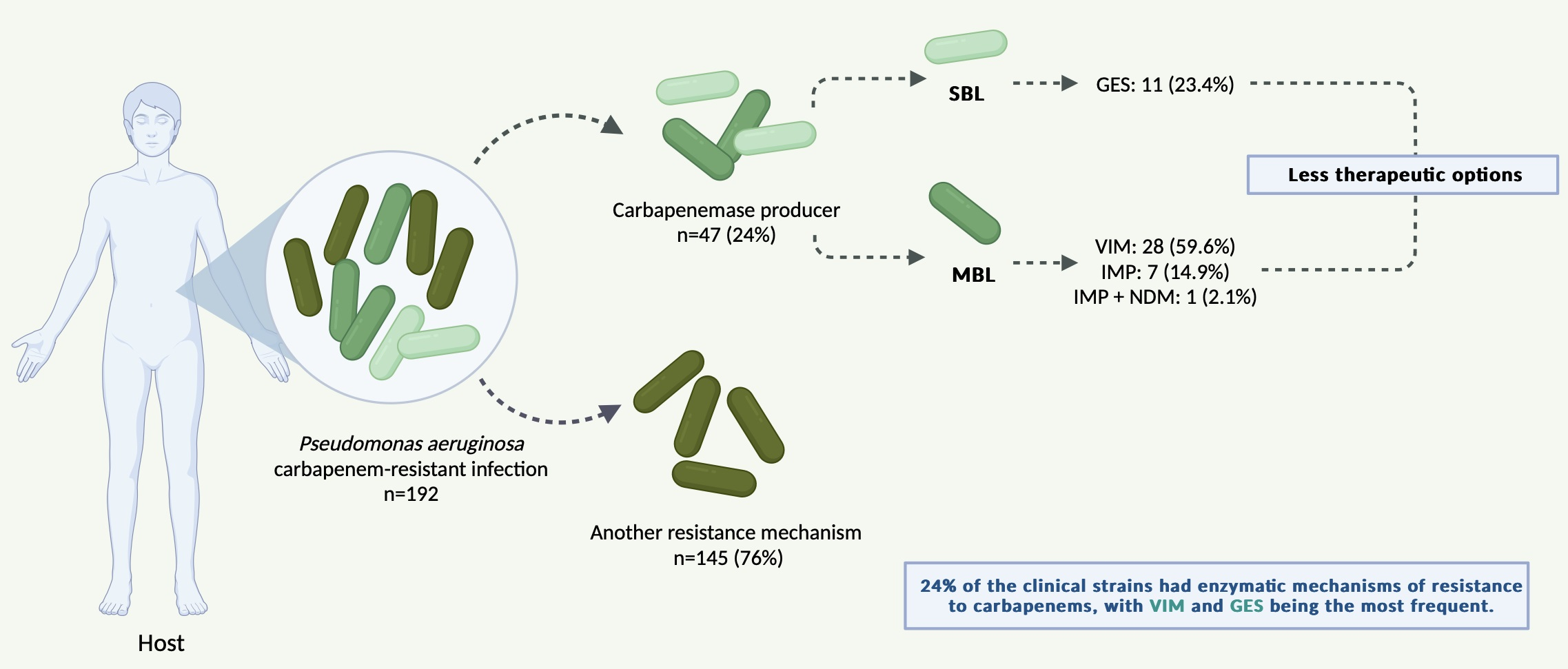
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Schoch, C.L.; Ciufo, S.; Domrachev, M.; Hotton, C.L.; Kannan, S.; Khovanskaya, R.; Leipe, D.; Mcveigh, R.; O’Neill, K.; Robbertse, B.; et al. NCBI Taxonomy: A Comprehensive Update on Curation, Resources and Tools. Database 2020, 2020, baaa062. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Jin, Y.; Bai, F.; Jin, S. Pseudomonas aeruginosa. In Molecular Medical Microbiology; Academic Press: Cambridge, MA, USA, 2015; Chapter 41; pp. 753–767. [Google Scholar]
- Klockgether, J.; Munder, A.; Neugebauer, J.; Davenport, C.F.; Stanke, F.; Larbig, K.D.; Heeb, S.; Schöck, U.; Pohl, T.M.; Wiehlmann, L.; et al. Genome Diversity of Pseudomonas aeruginosa PAO1 Laboratory Strains. J. Bacteriol. 2010, 192, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Amanati, A.; Sajedianfard, S.; Khajeh, S.; Ghasempour, S.; Mehrangiz, S.; Nematolahi, S.; Shahhosein, Z. Bloodstream Infections in Adult Patients with Malignancy, Epidemiology, Microbiology, and Risk Factors Associated with Mortality and Multi-Drug Resistance. BMC Infect. Dis. 2021, 21, 636. [Google Scholar] [CrossRef] [PubMed]
- Litwin, A.; Rojek, S.; Gozdzik, W.; Duszynska, W. Pseudomonas aeruginosa Device Associated—Healthcare Associated Infections and Its Multidrug Resistance at Intensive Care Unit of University Hospital: Polish, 8.5-Year, Prospective, Single-Centre Study. BMC Infect. Dis. 2021, 21, 180. [Google Scholar] [CrossRef]
- CDC. Pseudomonas aeruginosa in Healthcare Settings. Available online: https://www.cdc.gov/hai/organisms/pseudomonas.html (accessed on 20 August 2021).
- Rouzé, A.; Martin-Loeches, I.; Povoa, P.; Makris, D.; Artigas, A.; Bouchereau, M.; Lambiotte, F.; Metzelard, M.; Cuchet, P.; Boulle Geronimi, C.; et al. Relationship between SARS-CoV-2 Infection and the Incidence of Ventilator-Associated Lower Respiratory Tract Infections: A European Multicenter Cohort Study. Intensive Care Med. 2021, 47, 188–198. [Google Scholar] [CrossRef]
- World Health Organization WHO. Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 20 August 2021).
- Bassetti, M.; Poulakou, G.; Ruppe, E.; Bouza, E.; Van Hal, S.J.; Brink, A. Antimicrobial Resistance in the next 30 Years, Humankind, Bugs and Drugs: A Visionary Approach. Intensive Care Med. 2017, 43, 1464–1475. [Google Scholar] [CrossRef]
- Aslam, B.; Rasool, M.; Muzammil, S.; Siddique, A.B.; Nawaz, Z.; Shafique, M.; Zahoor, M.A.; Binyamin, R.; Waseem, M.; Khurshid, M.; et al. Carbapenem Resistance: Mechanisms and Drivers of Global Menace; IntechOpen: London, UK, 2020; ISBN 978-1-78985-988-1. [Google Scholar]
- Garza-González, E.; Franco-Cendejas, R.; Morfín-Otero, R.; Echaniz-Aviles, G.; Rojas-Larios, F.; Bocanegra-Ibarias, P.; Flores-Treviño, S.; Ponce-de-León, A.; Rodríguez-Noriega, E.; Alavez-Ramírez, N.; et al. The Evolution of Antimicrobial Resistance in Mexico During the Last Decade: Results from the INVIFAR Group. Microb. Drug Resist. 2020, 26, 1372–1382. [Google Scholar] [CrossRef]
- Lee, Y.-L.; Ko, W.-C.; Hsueh, P.-R. Geographic Patterns of Carbapenem-Resistant Pseudomonas aeruginosa in the Asia-Pacific Region: Results from the Antimicrobial Testing Leadership and Surveillance (ATLAS) Program, 2015–2019. Antimicrob. Agents Chemother. 2021, 66, e02000-21. [Google Scholar] [CrossRef]
- El Amin, N.; Giske, C.G.; Jalal, S.; Keijser, B.; Kronvall, G.; Wretlind, B. Carbapenem Resistance Mechanisms in Pseudomonas aeruginosa: Alterations of Porin OprD and Efflux Proteins Do Not Fully Explain Resistance Patterns Observed in Clinical Isolates. APMIS 2005, 113, 187–196. [Google Scholar] [CrossRef]
- Livermore, D.M. Of Pseudomonas, Porins, Pumps and Carbapenems. J. Antimicrob. Chemother. 2001, 47, 247–250. [Google Scholar] [CrossRef]
- Xavier, D.E.; Picão, R.C.; Girardello, R.; Fehlberg, L.C.; Gales, A.C. Efflux Pumps Expression and Its Association with Porin Down-Regulation and β-Lactamase Production among Pseudomonas aeruginosa Causing Bloodstream Infections in Brazil. BMC Microbiol. 2010, 10, 217. [Google Scholar] [CrossRef]
- Hammoudi Halat, D.; Ayoub Moubareck, C. The Current Burden of Carbapenemases: Review of Significant Properties and Dissemination among Gram-Negative Bacteria. Antibiotics 2020, 9, 186. [Google Scholar] [CrossRef]
- Queenan, A.M.; Bush, K. Carbapenemases: The Versatile Beta-Lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef] [PubMed]
- Yoon, E.-J.; Jeong, S.H. Mobile Carbapenemase Genes in Pseudomonas aeruginosa. Front. Microbiol. 2021, 12, 30. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Figueiredo, S.; Cattoir, V.; Carattoli, A.; Nordmann, P. Acinetobacter radioresistens as a Silent Source of Carbapenem Resistance for Acinetobacter spp. Antimicrob. Agents Chemother. 2008, 52, 1252–1256. [Google Scholar] [CrossRef] [PubMed]
- Garza-González, E.; Bocanegra-Ibarias, P.; Bobadilla-Del-Valle, M.; Ponce-de-León-Garduño, L.A.; Esteban-Kenel, V.; Silva-Sánchez, J.; Garza-Ramos, U.; Barrios-Camacho, H.; López-Jácome, L.E.; Colin-Castro, C.A.; et al. Drug Resistance Phenotypes and Genotypes in Mexico in Representative Gram-Negative Species: Results from the Infivar Network. PLoS ONE 2021, 16, e0248614. [Google Scholar] [CrossRef]
- Leber, A.L. Clinical Microbiology Procedures Handbook, 4th ed.; ASM Press: Washington, DC, USA, 2016; Volume 1–3, ISBN 978-1-55581-881-4. [Google Scholar]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; CLSI M07; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 31st ed.; CLSI M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2021. [Google Scholar]
- Bocanegra-Ibarias, P.; Garza-González, E.; Morfín-Otero, R.; Barrios, H.; Villarreal-Treviño, L.; Rodríguez-Noriega, E.; Garza-Ramos, U.; Petersen-Morfin, S.; Silva-Sanchez, J. Molecular and Microbiological Report of a Hospital Outbreak of NDM-1-Carrying Enterobacteriaceae in Mexico. PLoS ONE 2017, 12, e0179651. [Google Scholar] [CrossRef]
- Garza-Ramos, U.; Tinoco, P.; Silva-Sanchez, J.; Morfin-Otero, R.; Rodriguez-Noriega, E.; Leon-Garnica, G.; Sader, H.S.; Jones, R.N. Metallo-Beta-Lactamase IMP-18 Is Located in a Class 1 Integron (In96) in a Clinical Isolate of Pseudomonas aeruginosa from Mexico. Int. J. Antimicrob. Agents 2008, 31, 78–80. [Google Scholar] [CrossRef]
- Garza-Ramos, U.; Tinoco, P.; Rojas, T.; Carrillo, B.; Barajas, J.M.; Suarez, S.; Silva-Sanchez, J. Molecular Characterization of Integron Class 1 (In196) Encoding the VIM-2 Metallo-Beta-Lactamase of Pseudomonas aeruginosa Isolated from a Hospital Environment. J. Chemother. 2009, 21, 590–591. [Google Scholar] [CrossRef]
- Tamayo-Legorreta, E.M.; Garza-Ramos, U.; Barrios-Camacho, H.; Sanchez-Perez, A.; Galicia-Paredes, A.; Meza-Chavez, A.; Silva-Sanchez, J. Identification of OXA-23 Carbapenemases: Novel Variant OXA-239 in Acinetobacter Baumannii ST758 Clinical Isolates in Mexico. New Microbes New Infect. 2014, 2, 173–174. [Google Scholar] [CrossRef]
- Gerard, E. Dallal Randomization. Available online: http://www.jerrydallal.com/random/randomize.htm (accessed on 20 August 2021).
- Okamoto, K.; Gotoh, N.; Nishino, T. Pseudomonas aeruginosa Reveals High Intrinsic Resistance to Penem Antibiotics: Penem Resistance Mechanisms and Their Interplay. Antimicrob. Agents Chemother. 2001, 45, 1964–1971. [Google Scholar] [CrossRef]
- RHOVE. Informes Red Hospitalaria de Vigilancia Epidemiológica (RHOVE). 2015. Available online: http://www.gob.mx/salud/documentos/informes-rhove-2015 (accessed on 20 August 2021).
- Peña, C.; Suarez, C.; Gozalo, M.; Murillas, J.; Almirante, B.; Pomar, V.; Aguilar, M.; Granados, A.; Calbo, E.; Rodríguez-Baño, J.; et al. Prospective Multicenter Study of the Impact of Carbapenem Resistance on Mortality in Pseudomonas aeruginosa Bloodstream Infections. Antimicrob. Agents Chemother. 2012, 56, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Peña, C.; Suarez, C.; Ocampo-Sosa, A.; Murillas, J.; Almirante, B.; Pomar, V.; Aguilar, M.; Granados, A.; Calbo, E.; Rodríguez-Baño, J.; et al. Effect of Adequate Single-Drug vs Combination Antimicrobial Therapy on Mortality in Pseudomonas aeruginosa Bloodstream Infections: A Post Hoc Analysis of a Prospective Cohort. Clin. Infect. Dis. 2013, 57, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Baumgart, A.M.K.; Molinari, M.A.; Silveira, A.C. de O. Prevalence of Carbapenem Resistant Pseudomonas aeruginosa and Acinetobacter Baumannii in High Complexity Hospital. Braz. J. Infect. Dis. 2010, 14, 433–436. [Google Scholar] [CrossRef]
- De Matos, E.C.O.; de Matos, H.J.; Conceição, M.L.; Rodrigues, Y.C.; do Rosário Souza Carneiro, I.C.; Lima, K.V.B. Clinical and Microbiological Features of Infections Caused by Pseudomonas aeruginosa in Patients Hospitalized in Intensive Care Units. Rev. Soc. Bras. Med. Trop. 2016, 49, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, E.; Malecki, M.; Tellez-Castillo, C.J.; Pfennigwerth, N.; Marlinghaus, L.; Higgins, P.G.; Mattner, F.; Wendel, A.F. Molecular Surveillance of Carbapenemase-Producing Pseudomonas aeruginosa at Three Medical Centres in Cologne, Germany. Antimicrob. Resist. Infect. Control 2019, 8, 208. [Google Scholar] [CrossRef]
- Çekin, Z.K.; Dabos, L.; Malkoçoğlu, G.; Fortineau, N.; Bayraktar, B.; Iorga, B.I.; Naas, T.; Aktaş, E. Carbapenemase -Producing Pseudomonas aeruginosa Isolates from Turkey: First Report of P. Aeruginosa High-Risk Clones with VIM-5- and IMP-7-Type Carbapenemases in a Tertiary Hospital. Diagn. Microbiol. Infect. Dis. 2021, 99, 115174. [Google Scholar] [CrossRef]
- Liakopoulos, A.; Mavroidi, A.; Katsifas, E.A.; Theodosiou, A.; Karagouni, A.D.; Miriagou, V.; Petinaki, E. Carbapenemase-Producing Pseudomonas aeruginosa from Central Greece: Molecular Epidemiology and Genetic Analysis of Class I Integrons. BMC Infect. Dis. 2013, 13, 505. [Google Scholar] [CrossRef]
- Garza-Ramos, U.; Barrios, H.; Reyna-Flores, F.; Tamayo-Legorreta, E.; Catalan-Najera, J.C.; Morfin-Otero, R.; Rodríguez-Noriega, E.; Volkow, P.; Cornejo-Juarez, P.; González, A.; et al. Widespread of ESBL- and Carbapenemase GES-Type Genes on Carbapenem-Resistant Pseudomonas aeruginosa Clinical Isolates: A Multicenter Study in Mexican Hospitals. Diagn. Microbiol. Infect. Dis. 2015, 81, 135–137. [Google Scholar] [CrossRef] [PubMed]
- van Duin, D.; Bonomo, R.A. Ceftazidime/Avibactam and Ceftolozane/Tazobactam: Second-Generation β-Lactam/β-Lactamase Inhibitor Combinations. Clin. Infect. Dis. 2016, 63, 234–241. [Google Scholar] [CrossRef]
- Del Barrio-Tofiño, E.; López-Causapé, C.; Oliver, A. Pseudomonas aeruginosa Epidemic High-Risk Clones and Their Association with Horizontally-Acquired β-Lactamases: 2020 Update. Int. J. Antimicrob. Agents 2020, 56, 106196. [Google Scholar] [CrossRef] [PubMed]
- Zavascki, A.P.; Gaspareto, P.B.; Martins, A.F.; Gonçalves, A.L.; Barth, A.L. Outbreak of Carbapenem-Resistant Pseudomonas aeruginosa Producing SPM-1 Metallo-β-Lactamase in a Teaching Hospital in Southern Brazil. J. Antimicrob. Chemother. 2005, 56, 1148–1151. [Google Scholar] [CrossRef] [PubMed]
- Hishinuma, T.; Tada, T.; Kuwahara-Arai, K.; Yamamoto, N.; Shimojima, M.; Kirikae, T. Spread of GES-5 Carbapenemase-Producing Pseudomonas aeruginosa Clinical Isolates in Japan Due to Clonal Expansion of ST235. PLoS ONE 2018, 13, e0207134. [Google Scholar] [CrossRef] [PubMed]
- Bakthavatchalam, Y.D.; Anandan, S.; Veeraraghavan, B. Laboratory Detection and Clinical Implication of Oxacillinase-48 like Carbapenemase: The Hidden Threat. J. Glob. Infect. Dis. 2016, 8, 41–50. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences | Tm (°C) | Amplicon Size (bp) |
|---|---|---|---|
| blaNDM | F: 5′-ATGGAATTGCCGAATATT-3′ | 56 | ~600 |
| R: 5′-TCAGYGCAGCTTGTCGGC-3′ | |||
| blaIMP | F: 5′-GTTTATGTTCATACTTCGTTTG-3′ | 52 | ~400 |
| R: 5′-CAACCAGTTTTGCHTTAC-3′ | |||
| blaVIM | F: 5′-AGATTGVCGATGGTGTTTGGT-3′ | 56 | ~400 |
| R: 5′-GAGCAAGTCTAGACCGCCC-3′ | |||
| blaKPC | F: 5′-ATGTCACTGTATCGCCGTCT-3′ | 56 | 798 |
| R: 5′-TTACTGCCCGTTGACGC-3′ | |||
| blaGES | F: 5′-TCATTCACGCHCTATTVCTGGCA-3′ | 58 | 857 |
| R: 5′-CTATTTGTCCGTGCTCAGG-3′ | |||
| blaOXA-23 | F: 5′-TCTGGTTGTACGGTTCA-3′ | 56 | ~300 |
| R: 5′-TCATTACGTATAGATGCC-3′ | |||
| blaOXA-40 | F: 5′-TGAAGCTCAAACACAGGG-3′ | 56 | ~400 |
| R: 5′-AACACCCATTCCCCATCC-3′ | |||
| blaOXA-48 | F: 5′-GAATGCCTGCGGTAGCAA-3′ | 56 | 438 |
| R: 5′-AAACCATCCGATGTGGGCAT-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Zavaleta, M.G.; Fernández-Rodríguez, D.; Hernández-Durán, M.; Colín-Castro, C.A.; de Lourdes García-Hernández, M.; Becerra-Lobato, N.; Franco-Cendejas, R.; López-Jácome, L.E. Acquired blaVIM and blaGES Carbapenemase-Encoding Genes in Pseudomonas aeruginosa: A Seven-Year Survey Highlighting an Increasing Epidemiological Threat. Pathogens 2023, 12, 1256. https://doi.org/10.3390/pathogens12101256
Martínez-Zavaleta MG, Fernández-Rodríguez D, Hernández-Durán M, Colín-Castro CA, de Lourdes García-Hernández M, Becerra-Lobato N, Franco-Cendejas R, López-Jácome LE. Acquired blaVIM and blaGES Carbapenemase-Encoding Genes in Pseudomonas aeruginosa: A Seven-Year Survey Highlighting an Increasing Epidemiological Threat. Pathogens. 2023; 12(10):1256. https://doi.org/10.3390/pathogens12101256
Chicago/Turabian StyleMartínez-Zavaleta, María Guadalupe, Diana Fernández-Rodríguez, Melissa Hernández-Durán, Claudia A. Colín-Castro, María de Lourdes García-Hernández, Noé Becerra-Lobato, Rafael Franco-Cendejas, and Luis Esaú López-Jácome. 2023. "Acquired blaVIM and blaGES Carbapenemase-Encoding Genes in Pseudomonas aeruginosa: A Seven-Year Survey Highlighting an Increasing Epidemiological Threat" Pathogens 12, no. 10: 1256. https://doi.org/10.3390/pathogens12101256
APA StyleMartínez-Zavaleta, M. G., Fernández-Rodríguez, D., Hernández-Durán, M., Colín-Castro, C. A., de Lourdes García-Hernández, M., Becerra-Lobato, N., Franco-Cendejas, R., & López-Jácome, L. E. (2023). Acquired blaVIM and blaGES Carbapenemase-Encoding Genes in Pseudomonas aeruginosa: A Seven-Year Survey Highlighting an Increasing Epidemiological Threat. Pathogens, 12(10), 1256. https://doi.org/10.3390/pathogens12101256