
Dynamic and Seasonal Distribution of Enteric Viruses in Surface and Well Water in Riyadh (Saudi Arabia)

 , ,
, , 
Abstract
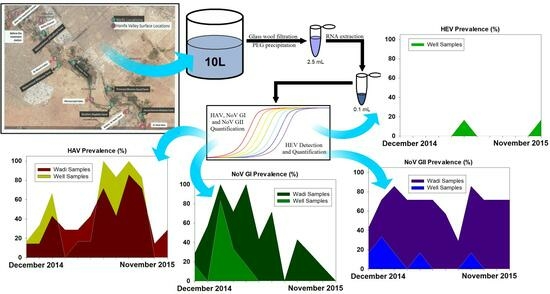
:
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Viral Concentration
2.3. Total Nucleic Acid Extraction
2.4. Virus Detection and Quantification (RT-qPCR)
2.4.1. HAV, NoV GI, and NoV GII
2.4.2. HEV
2.5. Statistical Analysis
3. Results
3.1. Seasonal Distribution of Enteric Viruses in Surface Water
3.1.1. Prevalence of Hepatitis Viruses
3.1.2. Prevalence of NoVGI and GII
3.2. Seasonal Distribution of Enteric Viruses in Wells
3.2.1. Prevalence of Hepatitis Viruses
3.2.2. Prevalence of NoV GI and GII
3.3. Differences between Sampling Areas
3.4. Comparison of Viral Load in Wadi Hanifa Lake and the Neighboring Wells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ganesh, A.; Lin, J. Waterborne human pathogenic viruses of public health concern. Int. J. Environ. Health Res. 2013, 23, 544–564. [Google Scholar] [CrossRef]
- Atmar, R.L.; Opekun, A.R.; Gilger, M.A.; Estes, M.K.; Crawford, S.E.; Neill, F.H.; Ramani, S.; Hill, H.; Ferreira, J.; Graham, D.Y. Determination of the 50% Human Infectious Dose for Norwalk Virus. J. Infect. Dis. 2014, 209, 1016–1022. [Google Scholar] [CrossRef]
- Bosch, A.; Pintó, R.M.; Guix, S. Foodborne viruses. Curr. Opinion Food Sci. 2016, 8, 110–119. [Google Scholar] [CrossRef]
- Lanrewaju, A.A.; Enitan-Folami, A.M.; Sabiu, S.; Edokpayi, J.N.; Swalaha, F.M. Global public health implications of human exposure to viral contaminated water. Front. Microbiol. 2022, 13, 981896. [Google Scholar] [CrossRef]
- Ahmed, S.M.; Hall, A.J.; Robinson, A.E.; Verhoef, L.; Premkumar, P.; Parashar, U.D.; Koopmans, M.; Lopman, B.A. Global Prevalence of Norovirus in Cases of Gastroenteritis: A Systematic Review and Meta-Analysis. Lancet Infect. Dis. 2014, 14, 725–730. [Google Scholar] [CrossRef]
- Lartey, B.L.; Quaye, O.; Damanka, S.A.; Agbemabiese, C.A.; Armachie, J.; Dennis, F.E.; Enweronu-Laryea, C.; Armah, G.E. Understanding pediatric norovirus epidemiology: A decade of study among Ghanaian Children. Viruses 2020, 12, 1321. [Google Scholar] [CrossRef]
- Mans, J. Norovirus Infections and Disease in Lower-Middle and Low-Income Countries, 1997–2018. Viruses 2019, 11, 341. [Google Scholar] [CrossRef]
- Vinjé, J. Advances in laboratory methods for detection and typing of norovirus. J. Clin. Microbiol. 2015, 53, 373–381. [Google Scholar] [CrossRef]
- White, P.A. Evolution of norovirus. Clin. Microbiol. Infect. 2014, 20, 741–745. [Google Scholar] [CrossRef]
- Laconi, A.; Cavicchio, L.; Tassoni, L.; Cunial, G.; Milani, A.; Ustulin, M.; Di Martino, G.; Forzan, M.; Campalto, M.; Monne, I.; et al. Identification of two divergent swine Noroviruses detected at the slaughterhouse in North East Italy. Porcine Health Manag. 2020, 6, 9. [Google Scholar] [CrossRef]
- Mathew, S.; Alansari, K.; Smatti, M.K.; Zaraket, H.; Al Thani, A.A.; Yassine, H.M. Epidemiological, molecular, and clinical features of norovirus infections among pediatric patients in Qatar. Viruses 2019, 11, 400. [Google Scholar] [CrossRef]
- Ge, L.; Chen, X.; Liu, J.; Zheng, L.; Chen, C.; Luo, S.; Guo, P.; Kong, J.; Song, Y.; Huo, Y. Genomic and biological characterization of a pandemic norovirus variant GII. 4 Sydney 2012. Virus Genes 2020, 56, 174–181. [Google Scholar] [CrossRef]
- La Rosa, G.; Pourshaban, M.; Iaconelli, M.; Muscillo, M. Quantitative real-time PCR of enteric viruses in influent and effluent samples from wastewater treatment plants in Italy. Ann. Sanita 2010, 46, 266–273. [Google Scholar]
- Miura, T.; Gima, A.; Akiba, M. Detection of Norovirus and Rotavirus present in suspended and dissolved forms in drinking water sources. Food Environ. Virol. 2019, 11, 9–19. [Google Scholar] [CrossRef]
- Mounts, A.W.; Ando, T.; Koopmans, M.; Bresee, J.S.; Noel, J.; Glass, R.I. Cold weather seasonality of gastroenteritis associated with Norwalk-like viruses. J. Infect. Dis. 2000, 181, S284–S287. [Google Scholar] [CrossRef]
- Van Damme, P.; Pintó, R.M.; Feng, Z.; Cui, F.; Gentile, A.; Shouval, A. Hepatitis A virus infection. Nat. Rev. Dis. Primers. 2023; in press. [Google Scholar]
- Pintó, R.M.; Pérez-Rodríguez, F.J.; Costafreda, M.I.; Chavarría-Miró, G.; Guix, S.; Ribes, E.; Bosch, A. Pathogenicity and virulenceof hepatitis A virus. Virulence 2021, 12, 1174–1185. [Google Scholar] [CrossRef]
- Rodríguez-Lázaro, D.; Cook, N.; Ruggeri, F.M.; Sellwood, J.; Nasser, A.; Nascimento, M.S.J.; Van der Poel, W.H. Virus hazards from food, water and other contaminated environments. FEMS Microbiol. Rev. 2012, 36, 786–814. [Google Scholar] [CrossRef]
- Cook, N.; Bertrand, I.; Gantzer, C.; Pintó, R.M.; Bosch, A. Persistence of Hepatitis A Virus in Fresh Produce and Production Environments, and the Effect of Disinfection Procedures: A Review. Food Environ. Virol. 2018, 10, 253–262. [Google Scholar] [CrossRef]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Met. 2006, 131, 65–71. [Google Scholar] [CrossRef]
- Schwarz, N.G.; Revillion, M.; Roque-Afonso, A.M.; Dussaix, E.; Giraud, M.; Liberpre, C.; Astagneau, E.D. A food-borne outbreak of hepatitis A virus (HAV) infection in a secondary school in Upper Normandy, France, in November 2006. Eurosurveillance 2008, 13, 18885. [Google Scholar] [CrossRef]
- Purpari, G.; Macaluso, G.; Di Bella, S.; Gucciardi, F.; Mira, F.; Di Marco, P.; Guercio, A. Molecular characterization of human enteric viruses in food, water samples, and surface swabs in Sicily. Inter. J. Infect. Dis. 2019, 80, 66–72. [Google Scholar] [CrossRef]
- Tanaka, S.; Kishi, T.; Ishihara, A.; Watanabe, D.; Uehira, T.; Ishida, H.; Mita, E. Outbreak of hepatitis A linked to European outbreaks among men who have sex with men in Osaka, Japan, from March to July 2018. Hepatol. Res. 2019, 49, 705–710. [Google Scholar] [CrossRef]
- Elbashir, S.; Parveen, S.; Schwarz, J.; Rippen, T.; Jahncke, M.; DePaola, A. Seafood Pathogens and Information on Antimicrobial Resistance: A review. Food Microbiol. 2018, 70, 85–93. [Google Scholar] [CrossRef]
- Ahmed, A.; Ali, I.A.; Ghazal, H.; Fazili, J.; Nusrat, S. Mystery of Hepatitis E Virus: Recent Advances in Its Diagnosis and Management. Int. J. Hepatol. 2015, 2015, 872431. [Google Scholar] [CrossRef]
- Navaneethan, U.; Al Mohajer, M.; Shata, M.T. Hepatitis E and pregnancy: Understanding the pathogenesis. Liver Int. 2008, 28, 1190–1199. [Google Scholar] [CrossRef]
- Raji, Y.E.; Toung, O.P.; Taib, N.M.; Sekawi, Z.B. Hepatitis E Virus: An emerging enigmatic and underestimated pathogen. Saudi J. Biol. Sci. 2022, 29, 499–512. [Google Scholar] [CrossRef]
- Purdy, M.A.; Harrison, T.J.; Jameel, S.; Meng, X.J.; Okamoto, H.; Van der Poel, W.H.M.; Smith, D.B.; Consortium, I.R. ICTV virus taxonomy profile: Hepeviridae. J. Gen. Virol. 2017, 98, 2645. [Google Scholar] [CrossRef]
- Upfold, N.S.; Luke, G.A.; Knox, C. Occurrence of human enteric viruses in water sources and shellfish: A focus on Africa. Food Environ. Virol. 2021, 13, 1–31. [Google Scholar] [CrossRef]
- Lee, G.H.; Tan, B.H.; Teo, E.C.Y.; Lim, S.G.; Dan, Y.Y.; Wee, A.; Aw, P.P.K.; Zhu, Y.; Hibberd, M.L.; Tan, C.K.; et al. Chronic infection with camelid hepatitis E virus in a liver transplant recipient who regularly consumes camel meat and milk. Gastroenterology 2016, 150, 355–357. [Google Scholar] [CrossRef]
- Smith, D.B.; Simmonds, P.; International Committee on the Taxonomy of Viruses Hepeviridae Study Group; Jameel, S.; Emerson, S.U.; Harrison, T.J.; Meng, X.J.; Okamoto, H.; Van der Poel, W.H.M.; Purdy, M.A. Consensus proposals for classification of the family Hepeviridae. J. Gen. Virol. 2014, 95, 2223–2232. [Google Scholar] [CrossRef]
- Sander, A.L.; Corman, V.M.; Lukashev, A.N.; Drexler, J.F. Evolutionary origins of enteric hepatitis viruses. Cold Spring Harbor Perspect. Med. 2018, 8, a031690. [Google Scholar] [CrossRef]
- Badur, S.; Öztürk, S.; Ozakay, A.; Khalaf, M.; Saha, D.; Van Damme, P.A. Review of the Experience of Childhood Hepatitis A Vaccination in Saudi Arabia and Turkey: Implications for hepatitis A Control and Prevention in the Middle East and North African Region. Hum. Vaccin. Immunother. 2021, 17, 3710–3728. [Google Scholar] [CrossRef]
- Nour, I.; Hanif, A.; Alanazi, I.O.; Al-Ashkar, I.; Alhetheel, A.; Eifan, S. Novel insights of waterborne human rotavirus A in Riyadh (Saudi Arabia) involving G2 predominance and Emergence of a Thermotolerant sequence. Sci. Rep. 2021, 11, 12132. [Google Scholar] [CrossRef]
- Arif, M.; Qattan, I.; Al-Faleh, F.; Ramia, S. Epidemiology of Hepatitis E virus (HEV) Infection in Saudi Arabia. Ann. Trop. Med. Parasitol. 1994, 88, 163–168. [Google Scholar] [CrossRef]
- Kheyami, A.M. Rotavirus gastroenteritis and strain diversity in Saudi Arabia. Current status and future prospects. Saudi Med. J. 2010, 31, 276–279. [Google Scholar]
- Tayeb, H.T.; Dela Cruz, D.M.; Al-Qahtani, A.; Al-Ahdal, M.N.; Carter, M.J. Enteric viruses in pediatric diarrhea in Saudi Arabia. J. Med. Virol. 2008, 80, 1919–1929. [Google Scholar] [CrossRef]
- Tayeb, H.T.; Al-Ahdal, M.N.; Cartear, M.J.; Al-Qahtani, A.A.; Cruz, D.M.D. Molecular epidemiology of human astrovirus infections in Saudi Arabia pediatric patients. J. Med. Virol. 2010, 82, 2038–2042. [Google Scholar] [CrossRef]
- Nour, I.; Hanif, A.; Zakri, A.M.; Al-Ashkar, I.; Alhetheel, A.; Eifan, S. Human adenovirus molecular characterization in various water environments and seasonal impacts in Riyadh, Saudi Arabia. Int. J. Environ. Res. Public Health 2021, 18, 4773. [Google Scholar] [CrossRef]
- Nour, I.; Hanif, A.; Alanazi, F.; Zakri, A.M.; Al-Ashkar, I.; Alhetheel, A.; Eifan, S. Evaluation of three different concentration and extraction methods for recovery efficiency of human adenovirus and human rotavirus virus A. J. Virol. Met. 2021, 295, 114212. [Google Scholar] [CrossRef]
- Blanco, A.; Abid, I.; Al-Otaibi, N.; Pérez-Rodríguez, F.J.; Fuentes, C.; Guix, S.; Pintó, R.M.; Bosch, A. Glass Wool Concentration Optimization for the Detection of Enveloped and Non-enveloped Waterborne Viruses. Food Environ. Virol. 2019, 11, 184–192. [Google Scholar] [CrossRef]
- Fuentes, C.; Guix, S.; Pérez-Rodriguez, F.J.; Fuster, N.; Carol, M.; Pintó, R.M.; Bosch, A. Standardized Multiplex one-step qRT-PCR for Hepatitis A virus, Norovirus GI and GII Quantification in bivalve Mollusks and Water. Food Microbiol. 2014, 40, 55–63. [Google Scholar] [CrossRef]
- Costafreda, M.I.; Bosch, A.; Pintó, R.M. Development, evaluation, and standardization of a real-time TaqMan Reverse Transcription-PCR Assay for Quantification of Hepatitis A Virus in Clinical and Shellfish Samples. Appl. Environ. Microbiol. 2006, 72, 3846–3855. [Google Scholar] [CrossRef]
- Da Silva, A.K.; Le Saux, J.C.; Parnaudeau, S.; Pommepuy, M.; Elimelech, M.; Le Guyader, F.S. Evaluation of Removal of Noroviruses during Wastewater Treatment, Using Real-Time Reverse Transcription-PCR: Different Behaviors of Genogroups I and II. Appl. Environ. Microbiol. 2007, 73, 7891–7897. [Google Scholar] [CrossRef] [PubMed]
- Loisy, F.; Atmar, R.L.; Guillon, P.; Le Cann, P.; Pommepuy, M.; Le Guyader, F.S. Real-time RT-PCR for Norovirus screening in Shellfish. J. Virol. Met. 2005, 123, 1–7. [Google Scholar] [CrossRef]
- Pinto, R.M.; Costafreda, M.I.; Bosch, A. Risk assessment in shellfish-borne outbreaks of hepatitis A. Appl. Environ. Microbiol. 2009, 75, 7350–7355. [Google Scholar] [CrossRef]
- Moosavi, S.; Makhzoumi, J.; Grose, M. Landscape practice in the Middle East between local and global aspirations. Land. Res. 2016, 41, 265–278. [Google Scholar] [CrossRef]
- Alhamid, A.A.; Alfayzi, S.A.; Hamadto, M.A. A Sustainable Water Resources Management Plan for Wadi Hanifa in Saudi Arabia. J. King Saud Uni. 2007, 19, 209–221. [Google Scholar] [CrossRef]
- Al Dossary, R.A.; Alnafie, A.N.; Aljaroodi, S.A.; Rahman, J.U.; Hunasemarada, B.C.; Alkharsah, K.R. Prevalence of Hepatitis E Virus Infection Among Blood Donors in the Eastern Province of Saudi Arabia. J. Multi. Health 2021, 27, 2381–2390. [Google Scholar] [CrossRef] [PubMed]
- El-Kafrawy, S.A.; Hassan, A.M.; El-Daly, M.M.; Qadri, I.; Tolah, A.M.; Al-Subhi, T.L.; Alzahrani, A.A.; Alsaaidi, G.A.; Al-Abdullah, N.; Kaki, R.M.; et al. Seroprevalence of Dromedary Camel HEV in Domestic and Imported Camels from Saudi Arabia. Viruses 2020, 12, 553. [Google Scholar] [CrossRef]
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Tsang, A.K.; Joseph, M.; Wong, E.Y.; Tang, Y.; Sivakumar, S.; Xie, J.; Bai, R.; et al. New hepatitis E virus genotype in camels, the Middle East. Emerg. Infect. Dis. 2014, 20, 1044–1048. [Google Scholar] [CrossRef]
- El-Kafrawy, S.A.; Hassan, A.M.; El-Daly, M.M.; Al-Hajri, M.; Farag, E.; Elnour, F.A.; Khan, A.; Tolah, A.M.; Alandijany, T.A.; Othman, N.A.; et al. Genetic Diversity of Hepatitis E virus (HEV) in Imported and Domestic Camels in Saudi Arabia. Sci. Rep. 2022, 12, 7005. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus (Nt Position) | Primers |
|---|---|
| Mengovirus (110–209) | Fw: 5′-GCGGGTCCTGCCGAAAGT-3′ |
| Rv: 5′-TGCACGCCATCTTCATTCACA-3′ | |
| Probe (Multiplex): 5′-[VIC]-AGCACGTGGGAGGGCGATCG-[MGB]-3′ | |
| Probe (Monoplex): 5′-[6FAM]-AGCACGTGGGAGGGCGATCG-[MGB]-3′ | |
| Hepatitis A virus (68–240) | Fw: 5′-TCACCGCCGTTTGCCTAG-3′ |
| Rv: 5′-GAGCCCTGGAAGAAAG-3′ | |
| Probe (Multiplex): 5′-[6FAM]-CCTGAACCTGCAGGAATTAA-[MGB]-3′ | |
| Probe (Monoplex): 5′-[6FAM]-CCTGAACCTGCAGGAATTAA-[MGB]-3′ | |
| Norovirus GI (5291–5376) | Fw: 5′-CGCTGGATGCGNTTCCAT-3′ |
| Rv: 5′-CCTTAGACGCCATCATCATTTAC-3′ | |
| Probe (Multiplex): 5′-[TxRED]-TGGACAGGAGAYCGCRATCT-[IBRQ]-3′ | |
| Probe (Monoplex): 5′-[6FAM]-TGGACAGGAGAYCGCRATCT-[TAMRA]-3′ | |
| Norovirus GII (5012–5100) | Fw: 5′-ATGTTCAGRTGGATGAGRTTCTCWGA-3′ |
| Rv: 5′-TCGACGCCATCTTCATTCACA-3′ | |
| Probe (Multiplex): 5′-[ATTO]-AGCACGTGGGAGGGCGATCG-[BHQ]-3′ | |
| Probe (Monoplex): 5′-[6FAM]-AGCACGTGGGAGGGCGATCG-[TAMRA]-3′ | |
| Hepatitis E virus (5261–5330) | Fw: 5′-GGTGGTTTCTGGGGTGAC-3′ |
| Rv: 5′-AGGGGTTGGTTGGATGAA-3′ | |
| Probe: 5′-[6FAM]-TGATTCTCAGCCCTTCGC-[BHQ]-3′ |
| Viral Load (Log10 Mean ± SD) | ||||||
|---|---|---|---|---|---|---|
| Month | Surface Water | Well Water | ||||
| HAV | NoV GI | NoV GII | HAV | NoV GI | NoV GII | |
| December 2014 | 1.63 ± 0.00 | 3.40 ± 0.21 | 3.10 ± 0.99 | <1.69 ± 0.00 | <1.69 ± 0.00 | <1.69 ± 0.00 |
| January 2015 | <1.69 ± 0.00 | 3.36 ± 0.09 | 3.29 ± 0.83 | <1.69 ± 0.00 | NA | 1.75 ± 0.05 |
| February 2015 | 1.20 ± 0.50 | 3.06 ± 0.17 | 2.60 ± 0.75 | <1.69 ± 0.00 | 1.94 ± 0.39 | <1.69 ± 0.00 |
| March 2015 | 1.04 ± 0.65 | 3.45 ± 0.52 | 3.63 ± 0.36 | NA | 1.78 ± 0.08 | NA |
| April 2015 | 1.58 ± 0.11 | 2.46 ± 0.78 | 3.27 ± 0.87 | <1.69 ± 0.00 | <1.69 ± 0.00 | <1.69 ± 0.00 |
| May 2015 | 1.86 ± 0.23 | 2.63 ± 0.70 | 2.50 ± 1.01 | <1.69 ± 0.00 | NA | NA |
| June 2015 | 2.33 ± 0.52 | 3.76 ± 1.04 | 3.41 ± 0.16 | 2.15 ± 0.34 | NA | NA |
| July 2015 | 1.91 ± 0.15 | NA | 1.96 ± 0.27 | 1.66 ± 0.34 | NA | NA |
| August 2015 | 2.57 ± 0.83 | 3.36 ± 0.17 | 3.02 ± 0.97 | 2.72 ± 0.47 | NA | 1.68 ± 0.00 |
| September 2015 | 2.83 ± 0.98 | 1.69 ± 0.00 | 2.02 ± 0.30 | 1.90 ± 0.17 | NA | NA |
| October 2015 | 1.69 ± 0.00 | 2.94 ± 0.00 | 2.37 ± 0.58 | NA | NA | NA |
| November 2015 | 2.78 ± 0.29 | NA | 3.08 ± 0.93 | NA | NA | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abid, I.; Blanco, A.; Al-Otaibi, N.; Guix, S.; Costafreda, M.I.; Pintó, R.M.; Bosch, A. Dynamic and Seasonal Distribution of Enteric Viruses in Surface and Well Water in Riyadh (Saudi Arabia). Pathogens 2023, 12, 1405. https://doi.org/10.3390/pathogens12121405
Abid I, Blanco A, Al-Otaibi N, Guix S, Costafreda MI, Pintó RM, Bosch A. Dynamic and Seasonal Distribution of Enteric Viruses in Surface and Well Water in Riyadh (Saudi Arabia). Pathogens. 2023; 12(12):1405. https://doi.org/10.3390/pathogens12121405
Chicago/Turabian StyleAbid, Islem, Albert Blanco, Nawal Al-Otaibi, Susana Guix, Maria I. Costafreda, Rosa M. Pintó, and Albert Bosch. 2023. "Dynamic and Seasonal Distribution of Enteric Viruses in Surface and Well Water in Riyadh (Saudi Arabia)" Pathogens 12, no. 12: 1405. https://doi.org/10.3390/pathogens12121405