
COVID-19 Vaccination and Alcohol Consumption: Justification of Risks


Abstract
:1. Introduction
2. The Impacts of Alcohol Consumption on the Immune System
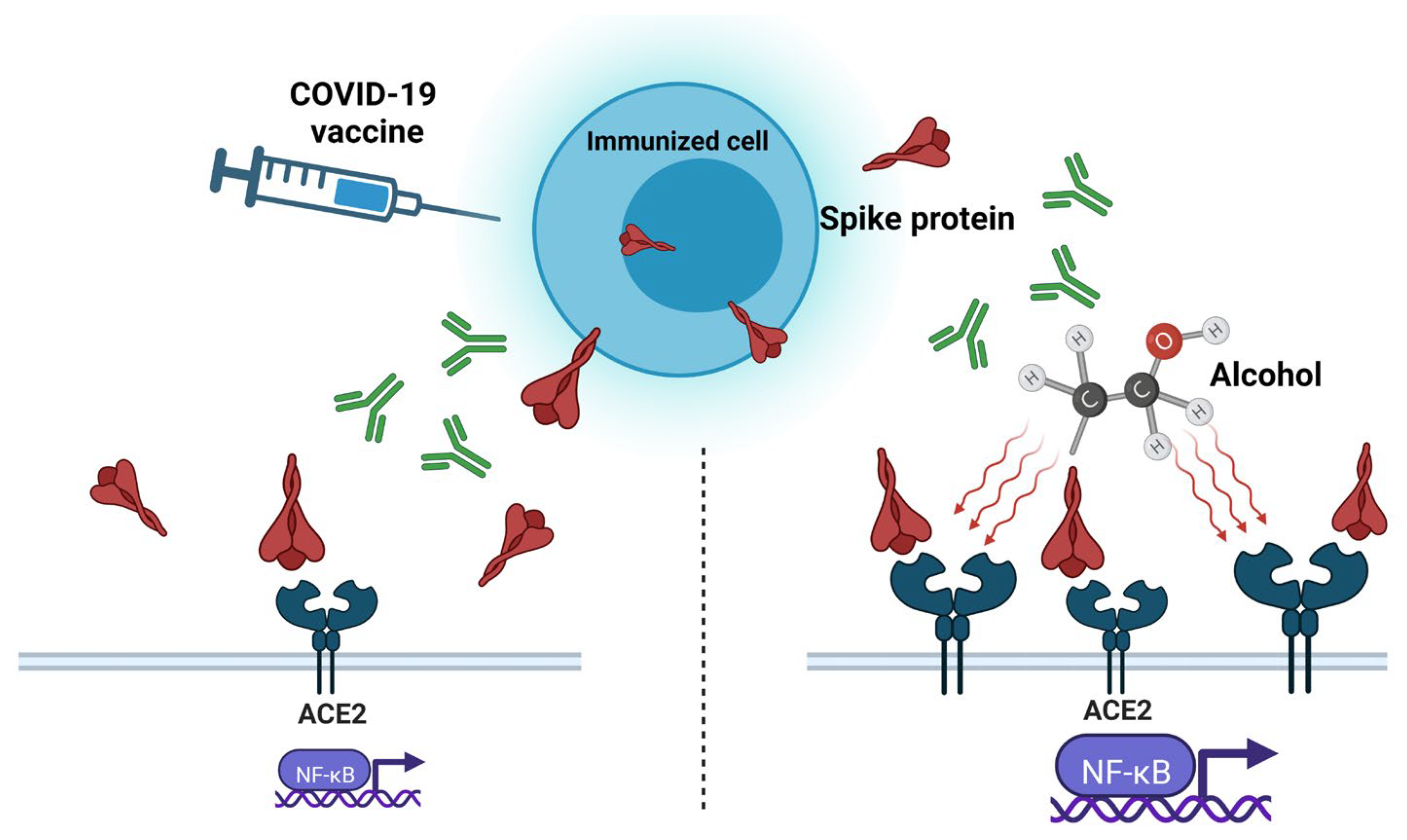
3. “Spike Effect” of COVID-19 Vaccines and Alcohol
4. Protein Subunit Vaccines and Alcohol
5. Inactivated Whole-Virus Vaccines and Alcohol
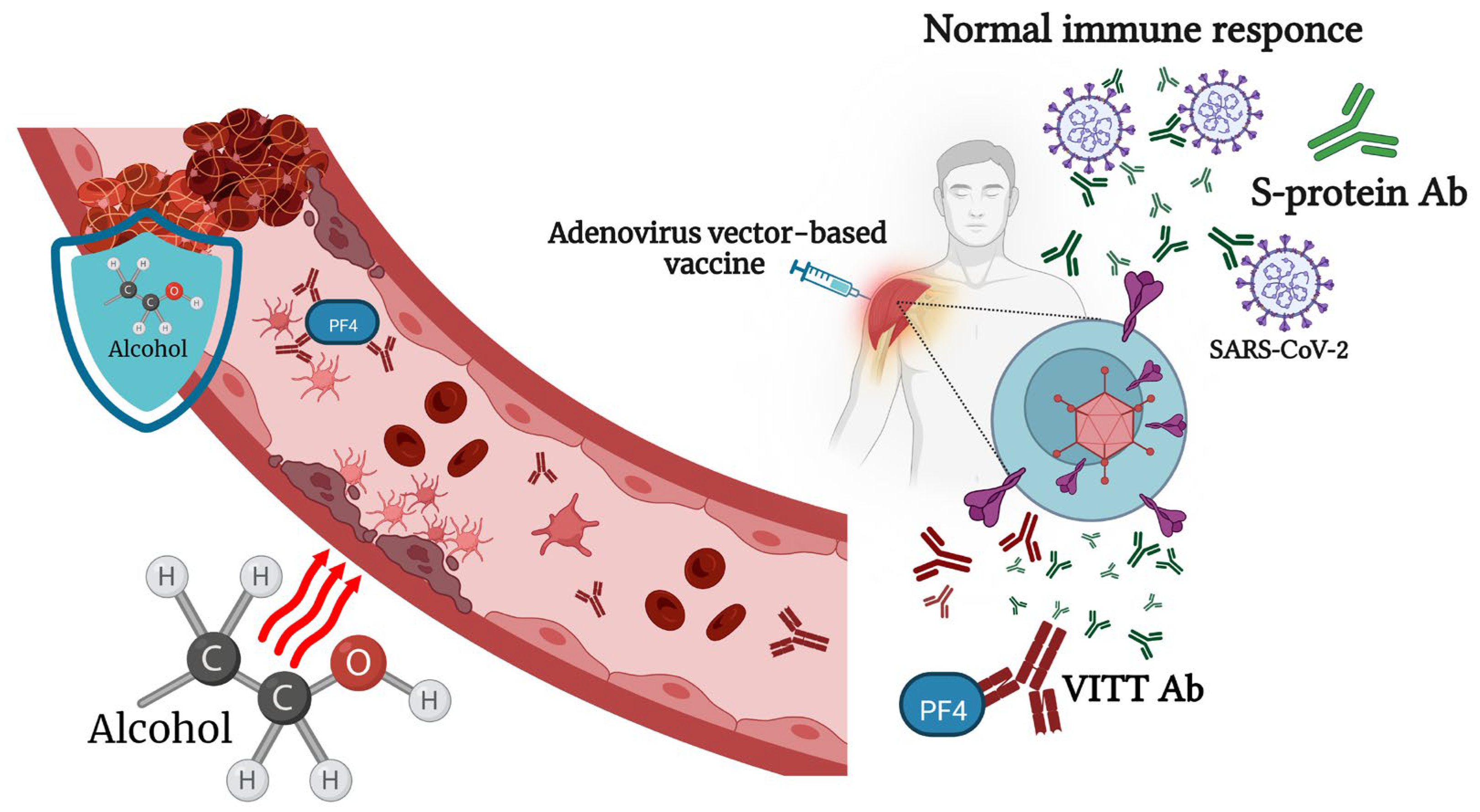
6. Viral-Vector-Based COVID-19 Vaccines and Alcohol
7. mRNA-Based Vaccines and Alcohol Consumption
8. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Messaoudi, I.; Pasala, S.; Grant, K. Could moderate alcohol intake be recommended to improve vaccine responses? Expert. Rev. Vaccines 2014, 13, 817–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, D.; Jung, M.K.; Wang, H.J. Alcohol and the Immune System. Alcohol Res. Curr. Rev. 2015, 37, 153–155. [Google Scholar]
- Alraiyes, A.H.; Shaheen, K.; Alraies, M.C. Alcoholic leukopenic pneumococcal sepsis. Avicenna J. Med. 2013, 3, 53–55. [Google Scholar] [CrossRef] [PubMed]
- Trannesen, H.; Andersen, J.R.; Pedersen, A.E.; Kaiser, A.H. Lymphopenia in heavy drinkers--reversibility and relation to the duration of drinking episodes. Ann. Med. 1990, 22, 229–231. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Quintela, A.; Alende, R.; Gude, F.; Campos-Franco, J.; Rey, J.; Meijide, L.M.; Fernandez-Merino, C.; Vidal, C. Serum levels of immunoglobulins (IgG, IgA, IgM) in a general adult population and their relationship with alcohol consumption, smoking and common metabolic abnormalities. Clin. Exp. Immunol. 2008, 151, 42–50. [Google Scholar] [CrossRef]
- Mili, F.; Flanders, W.D.; Boring, J.R.; Annest, J.L.; DeStefano, F. The associations of alcohol drinking and drinking cessation to measures of the immune system in middle-aged men. Alcohol. Clin. Exp. Res. 1992, 16, 688–694. [Google Scholar] [CrossRef]
- Barr, T.; Helms, C.; Grant, K.; Messaoudi, I. Opposing effects of alcohol on the immune system. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2016, 65, 242–251. [Google Scholar] [CrossRef] [Green Version]
- Meadows, G.G.; Zhang, H. Effects of Alcohol on Tumor Growth, Metastasis, Immune Response, and Host Survival. Alcohol Res. Curr. Rev. 2015, 37, 311–322. [Google Scholar]
- Fortner, A.; Schumacher, D. First COVID-19 Vaccines Receiving the US FDA and EMA Emergency Use Authorization. Discoveries 2021, 9, e122. [Google Scholar] [CrossRef]
- Castaldelli-Maia, J.M.; Segura, L.E.; Martins, S.S. The concerning increasing trend of alcohol beverage sales in the U.S. during the COVID-19 pandemic. Alcohol 2021, 96, 37–42. [Google Scholar] [CrossRef]
- Nadkarni, A.; Kapoor, A.; Pathare, S. COVID-19 and forced alcohol abstinence in India: The dilemmas around ethics and rights. Int. J. Law Psychiatry 2020, 71, 101579. [Google Scholar] [CrossRef] [PubMed]
- Pedrosa, A.L.; Bitencourt, L.; Fróes, A.C.F.; Cazumbá, M.L.B.; Campos, R.G.B.; de Brito, S.B.C.S.; Simões e Silva, A.C. Emotional, Behavioral, and Psychological Impact of the COVID-19 Pandemic. Front. Psychol. 2020, 11, 566212. [Google Scholar] [CrossRef] [PubMed]
- Anker, J.J.; Kushner, M.G. Co-Occurring Alcohol Use Disorder and Anxiety: Bridging Psychiatric, Psychological, and Neurobiological Perspectives. Alcohol Res. 2019, 40. [Google Scholar] [CrossRef]
- Aghababaeian, H.; Hamdanieh, L.; Ostadtaghizadeh, A. Alcohol intake in an attempt to fight COVID-19: A medical myth in Iran. Alcohol 2020, 88, 29–32. [Google Scholar] [CrossRef]
- Awijen, H.; Ben Zaied, Y.; Nguyen, D.K. Covid-19 vaccination, fear and anxiety: Evidence from Google search trends. Soc. Sci. Med. 2022, 297, 114820. [Google Scholar] [CrossRef]
- Bendau, A.; Plag, J.; Petzold, M.B.; Ströhle, A. COVID-19 vaccine hesitancy and related fears and anxiety. Int. Immunopharmacol. 2021, 97, 107724. [Google Scholar] [CrossRef]
- Chen, S.; Aruldass, A.R.; Cardinal, R.N. Mental health outcomes after SARS-CoV-2 vaccination in the United States: A national cross-sectional study. J. Affect. Disord. 2022, 298, 396–399. [Google Scholar] [CrossRef] [PubMed]
- McNeil, A.; Purdon, C. Anxiety disorders, COVID-19 fear, and vaccine hesitancy. J. Anxiety Disord. 2022, 90, 102598. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Q.; Davis, P.B.; Volkow, N.D.; Xu, R. Increased risk for COVID-19 breakthrough infection in fully vaccinated patients with substance use disorders in the United States between December 2020 and August 2021. World Psychiatry 2022, 21, 124–132. [Google Scholar] [CrossRef]
- Solopov, P.; Colunga Biancatelli, R.; Sharlow, E.; Lazo, J.; Catravas, J. Single intratracheal exposure to SARS-CoV-2 S1 spike protein induces acute lung injury in K18-hACE2 transgenic mice. FASEB J. 2021, 35. [Google Scholar] [CrossRef]
- Solopov, P.A.; Biancatelli, R.M.L.C.; Catravas, J.D. Alcohol Increases Lung Angiotensin-Converting Enzyme 2 Expression and Exacerbates Severe Acute Respiratory Syndrome Coronavirus 2 Spike Protein Subunit 1-Induced Acute Lung Injury in K18-hACE2 Transgenic Mice. Am. J. Pathol. 2022, 192, 990–1000. [Google Scholar] [CrossRef] [PubMed]
- Bautista, A.P. Free radicals, chemokines, and cell injury in HIV-1 and SIV infections and alcoholic hepatitis. Free Radic. Biol. Med. 2001, 31, 1527–1532. [Google Scholar] [CrossRef] [PubMed]
- Daniluk, J.; Szuster-Ciesielska, A.; Drabko, J.; Kandefer-Szerszeń, M. Serum cytokine levels in alcohol-related liver cirrhosis. Alcohol 2001, 23, 29–34. [Google Scholar] [CrossRef] [PubMed]
- González-Quintela, A.; Dominguez-Santalla, M.; Pérez, L.; Vidal, C.; Lojo, S.; Barrio, E. Influence of acute alcohol intake and alcohol withdrawal on circulating levels of IL-6, IL-8, IL-10 and IL-12. Cytokine 2000, 12, 1437–1440. [Google Scholar] [CrossRef]
- Zakhari, S. Overview: How is alcohol metabolized by the body? Alcohol Res. Health 2006, 29, 245–254. [Google Scholar]
- Yeligar, S.M.; Chen, M.M.; Kovacs, E.J.; Sisson, J.H.; Burnham, E.L.; Brown, L.A.S. Alcohol and lung injury and immunity. Alcohol 2016, 55, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Awaya, T.; Moroi, M.; Enomoto, Y.; Kunimasa, T.; Nakamura, M. What Should We Do after the COVID-19 Vaccination? Vaccine-Associated Diseases and Precautionary Measures against Adverse Reactions. Vaccines 2022, 10, 866. [Google Scholar] [CrossRef]
- Gentilello, L.M.; Cobean, R.A.; Walker, A.P.; Moore, E.E.; Wertz, M.J.; Dellinger, E.P. Acute ethanol intoxication increases the risk of infection following penetrating abdominal trauma. J. Trauma Inj. Infect. Crit. Care 1993, 34, 669–675. [Google Scholar] [CrossRef]
- Wyatt, T.A.; Gentry-Nielsen, M.J.; Pavlik, J.A.; Sisson, J.H. Desensitization of PKA-stimulated ciliary beat frequency in an ethanol-fed rat model of cigarette smoke exposure. Alcohol. Clin. Exp. Res. 2004, 28, 998–1004. [Google Scholar] [CrossRef] [Green Version]
- Cohen, S.; Tyrrell, D.A.; Russell, M.A.; Jarvis, M.J.; Smith, A. Smoking, alcohol consumption, and susceptibility to the common cold. Am. J. Public Health 1993, 83, 1277–1283. [Google Scholar] [CrossRef] [Green Version]
- Quesada-Molina, M.; Muñoz-Garach, A.; Tinahones, F.J.; Moreno-Indias, I. A New Perspective on the Health Benefits of Moderate Beer Consumption: Involvement of the Gut Microbiota. Metabolites 2019, 9, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, L.E.; Montero, A.; Gonzalez-Gross, M.; Vallejo, A.I.; Romeo, J.; Marcos, A. Influence of alcohol consumption on immunological status: A review. Eur. J. Clin. Nutr. 2002, 56, S50–S53. [Google Scholar] [CrossRef] [PubMed]
- Magrone, T.; Candore, G.; Caruso, C.; Jirillo, E.; Covelli, V. Polyphenols from Red Wine Modulate Immune Responsiveness: Biological and Clinical Significance. Curr. Pharm. Des. 2008, 14, 2733–2748. [Google Scholar] [CrossRef] [PubMed]
- Romeo, J.; Wärnberg, J.; Nova, E.; Díaz, L.E.; González-Gross, M.; Marcos, A. Changes in the immune system after moderate beer consumption. Ann. Nutr. Metab. 2007, 51, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Solopov, P.; Biancatelli, R.M.L.C.; Dimitropoulou, C.; Catravas, J.D. Dietary Phytoestrogens Ameliorate Hydrochloric Acid-Induced Chronic Lung Injury and Pulmonary Fibrosis in Mice. Nutrients 2021, 13, 3599. [Google Scholar] [CrossRef]
- Romeo, J.; Wärnberg, J.; Nova, E.; Díaz, L.E.; Gómez-Martinez, S.; Marcos, A. Moderate alcohol consumption and the immune system: A review. Br. J. Nutr. 2007, 98, S111–S115. [Google Scholar] [CrossRef]
- Van de Loo, A.; Raasveld, S.; Hogewoning, A.; Zeeuw, R.; Bosma, E.; Bouwmeester, N.; Lukkes, M.; Knipping, K.; Mackus, M.; Kraneveld, A.; et al. Immune Responses after Heavy Alcohol Consumption: Cytokine Concentrations in Hangover-Sensitive and Hangover-Resistant Drinkers. Healthcare 2021, 9, 395. [Google Scholar] [CrossRef]
- Mittal, A.; Manjunath, K.; Ranjan, R.K.; Kaushik, S.; Kumar, S.; Verma, V. COVID-19 pandemic: Insights into structure, function, and hACE2 receptor recognition by SARS-CoV-2. PLoS Pathog. 2020, 16, e1008762. [Google Scholar] [CrossRef]
- Almehdi, A.M.; Khoder, G.; Alchakee, A.S.; Alsayyid, A.T.; Sarg, N.H.; Soliman, S.S. SARS-CoV-2 spike protein: Pathogenesis, vaccines, and potential therapies. Infection 2021, 49, 855–876. [Google Scholar] [CrossRef]
- Martínez-Flores, D.; Zepeda-Cervantes, J.; Cruz-Reséndiz, A.; Aguirre-Sampieri, S.; Sampieri, A.; Vaca, L. SARS-CoV-2 Vaccines Based on the Spike Glycoprotein and Implications of New Viral Variants. Front. Immunol. 2021, 12, 701501. [Google Scholar] [CrossRef]
- Angeli, F.; Reboldi, G.; Trapasso, M.; Zappa, M.; Spanevello, A.; Verdecchia, P. COVID-19, vaccines and deficiency of ACE(2) and other angiotensinases. Closing the loop on the “Spike effect”. Eur. J. Intern. Med. 2022, 103, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Cognetti, J.S.; Miller, B.L. Monitoring Serum Spike Protein with Disposable Photonic Biosensors Following SARS-CoV-2 Vaccination. Sensors 2021, 21, 5857. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C. Could SARS-CoV-2 Spike Protein Be Responsible for Long-COVID Syndrome? Mol. Neurobiol. 2022, 59, 1850–1861. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.S.; Jeon, M.T.; Kim, K.S.; Lee, S.; Kim, S.; Kim, D.G. Spike Proteins of SARS-CoV-2 Induce Pathological Changes in Molecular Delivery and Metabolic Function in the Brain Endothelial Cells. Viruses 2021, 13, 2021. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, P.; Yang, N.; Huang, J.; Ou, J.; Xu, T.; Zhao, X.; Liu, T.; Huang, X.; Wang, Q.; et al. SARS-CoV-2 spike promotes inflammation and apoptosis through autophagy by ROS-suppressed PI3K/AKT/mTOR signaling. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166260. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Shafiei, M.; Longoria, C.; Schoggins, J.; Savani, R.; Zaki, H. SARS-CoV-2 spike protein induces inflammation via TLR2-dependent activation of the NF-κB pathway. Elife 2021, 10, e68563. [Google Scholar] [CrossRef]
- Biancatelli, R.M.L.C.; Solopov, P.A.; Gregory, B.; Khodour, Y.; Catravas, J.D. HSP90 Inhibitors Modulate SARS-CoV-2 Spike Protein Subunit 1-Induced Human Pulmonary Microvascular Endothelial Activation and Barrier Dysfunction. Front. Physiol. 2022, 13, 812199. [Google Scholar] [CrossRef]
- Biancatelli, R.M.C.; Solopov, P.A.; Sharlow, E.R.; Lazo, J.S.; Marik, P.E.; Catravas, J.D. The SARS-CoV-2 spike protein subunit S1 induces COVID-19-like acute lung injury in Κ18-hACE2 transgenic mice and barrier dysfunction in human endothelial cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 2021, 321, L477–L484. [Google Scholar] [CrossRef]
- Lei, Y.; Zhang, J.; Schiavon, C.R.; He, M.; Chen, L.; Shen, H.; Zhang, Y.; Yin, Q.; Cho, Y.; Andrade, L.; et al. SARS-CoV-2 Spike Protein Impairs Endothelial Function via Downregulation of ACE 2. Circ. Res. 2021, 128, 1323–1326. [Google Scholar] [CrossRef]
- Petruk, G.; Puthia, M.; Petrlova, J.; Samsudin, F.; Strömdahl, A.; Cerps, S.; Uller, L.; Kjellström, S.; Bond, P.; Schmidtchen, A. SARS-CoV-2 spike protein binds to bacterial lipopolysaccharide and boosts proinflammatory activity. J. Mol. Cell Biol. 2020, 12, 916–932. [Google Scholar] [CrossRef]
- Zhu, G.; Lyu, L.; Yang, H.; Liu, G.; Yang, S.; Gu, C.; Wang, L.; Yan, H.; Hu, M.; Che, C. SARS-CoV-2 spike protein-induced host inflammatory response signature in human corneal epithelial cells. Mol. Med. Rep. 2021, 24, 584. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.K.; Sozmen, E.G.; Dixit, K.; Montano, M.; Matsui, Y.; Liu, Y.E.; Helmy, T.J.; Deerinck, Z.; Yan, R.; Schuck, R.M.; et al. SARS-CoV-2 spike protein induces abnormal inflammatory blood clots neutralized by fibrin immunotherapy. bioRxiv 2021. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhao, J.; Li, J.; Guo, Z.; Sheng, J.; Ye, X.; Jin, G.; Wang, C.; Chai, W.; Yan, J.; et al. SARS-CoV-2 spike protein causes blood coagulation and thrombosis by competitive binding to heparan sulfate. Int. J. Biol. Macromol. 2021, 193, 1124–1129. [Google Scholar] [CrossRef] [PubMed]
- Boschi, C.; Scheim, D.E.; Bancod, A.; Militello, M.; Le Bideau, M.; Colson, P.; Fantini, J.; La Scola, B. SARS-CoV-2 Spike Protein Induces Hemagglutination: Implications for COVID-19 Morbidities and Therapeutics and for Vaccine Adverse Effects. Int. J. Mol. Sci. 2022, 23, 15480. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, N.; James, T.D.; Pushpavathi, S.G.; Marcinkiewcz, C.A. Repeated ethanol exposure and withdrawal alters ACE2 expression in discrete brain regions: Implications for SARS-CoV-2 infection. bioRxiv 2022. [Google Scholar] [CrossRef]
- Wang, F.; Yang, J.L.; Yu, K.K.; Xu, M.; Xu, Y.Z.; Chen, L.; Lu, Y.-m.; Fang, H.-s.; Wang, X.-y.; Hu, Z.-q.; et al. Wang, Activation of the NF-κB pathway as a mechanism of alcohol enhanced progression and metastasis of human hepatocellular carcinoma. Mol. Cancer 2015, 14, 10. [Google Scholar] [CrossRef] [Green Version]
- Vandenbulcke, M.; Janssens, J. Acute axonal polyneuropathy in chronic alcoholism and malnutrition. Acta Neurol. Belg. 1999, 99, 198–201. [Google Scholar]
- Worner, T.M. Guillain-Barré’s syndrome in alcoholics. Drug Alcohol Depend. 1989, 23, 93. [Google Scholar] [CrossRef]
- Kim, S.Y.; Oh, D.J.; Park, B.; Choi, H.G. Bell’s palsy and obesity, alcohol consumption and smoking: A nested case-control study using a national health screening cohort. Sci. Rep. 2020, 10, 4248. [Google Scholar] [CrossRef] [Green Version]
- Hansson, M.; Nygren, P.A.K.; Ståhl, S. Design and production of recombinant subunit vaccines. Biotechnol. Appl. Biochem. 2000, 32, 95–107. [Google Scholar] [CrossRef]
- Heidary, M.; Kaviar, V.H.; Shirani, M.; Ghanavati, R.; Motahar, M.; Sholeh, M.; Ghahramanpour, H.; Khoshnood, S. A Comprehensive Review of the Protein Subunit Vaccines Against COVID-19. Front. Microbiol. 2022, 13, 927306. [Google Scholar] [CrossRef] [PubMed]
- Heath, P.T.; Galiza, E.P.; Baxter, D.; Boffito, M.; Browne, D.; Burns, F.; Chadwick, D.; Clark, R.; Cosgrove, C.; Galloway, J.; et al. Safety and Efficacy of the NVX-CoV2373 COVID-19 Vaccine at Completion of the Placebo-Controlled Phase of a Randomized Controlled Trial. Clin. Infect Dis. 2022, ciac803. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.H.; Patel, N.; Haupt, R.; Zhou, H.; Weston, S.; Hammond, H.; Logue, J.; Portnoff, A.; Norton, J.; Guebre-Xabier, M.; et al. SARS-CoV-2 spike glycoprotein vaccine candidate NVX-CoV2373 immunogenicity in baboons and protection in mice. Nat. Commun. 2021, 12, 372. [Google Scholar] [CrossRef] [PubMed]
- Rydyznski Moderbacher, C.; Kim, C.; Mateus, J.; Plested, J.; Zhu, M.; Cloney-Clark, S.; Weiskopf, D.; Sette, A.; Fries, L.; Glenn, G.; et al. NVX-CoV2373 vaccination induces functional SARS-CoV-2-specific CD4+ and CD8+ T cell responses. J. Clin. Investig. 2022, 132, e160898. [Google Scholar] [CrossRef]
- Oh, J.; Cho, W.H.; Barcelon, E.; Kim, K.H.; Hong, J.; Lee, S.J. SARS-CoV-2 spike protein induces cognitive deficit and anx-iety-like behavior in mouse via non-cell autonomous hippocampal neuronal death. Sci. Rep. 2022, 12, 5496. [Google Scholar] [CrossRef]
- Barrett, P.N.; Mundt, W.; Kistner, O.; Howard, M.K. Vero cell platform in vaccine production: Moving towards cell cul-ture-based viral vaccines. Expert Rev. Vaccines 2009, 8, 607–618. [Google Scholar] [CrossRef]
- Sanders, B.P.; Koldijk, M.; Schuitemaker, H. Inactivated viral vaccines. In Vaccine Analysis: Strategies, Principles, and Control; Springer: Berlin/Heidelberg, Germany, 2015; pp. 45–80. [Google Scholar] [CrossRef]
- Liang, Z.; Zhu, H.; Wang, X.; Jing, B.; Li, Z.; Xia, X.; Sun, H.; Yang, Y.; Zhang, W.; Shi, L.; et al. Adjuvants for Coronavirus Vaccines. Front. Immunol. 2020, 11, 589833. [Google Scholar] [CrossRef]
- Facciolà, A.; Visalli, G.; Laganà, A.; Di Pietro, A. An Overview of Vaccine Adjuvants: Current Evidence and Future Per-spectives. Vaccines 2022, 10, 819. [Google Scholar] [CrossRef]
- Wang, C.; Chen, L.Y.; Lu, Q.B.; Cui, F. Vaccination with the Inactivated Vaccine (Sinopharm BBIBP-CorV) Ensures Pro-tection against SARS-CoV-2 Related Disease. Vaccines 2022, 10, 920. [Google Scholar] [CrossRef]
- Gao, Q.; Bao, L.; Mao, H.; Wang, L.; Xu, K.; Yang, M.; Li, Y.; Zhu, L.; Wang, N.; Lv, Z.; et al. Development of an inactivated vaccine candidate for SARS-CoV-2. Science 2020, 369, 77–81. [Google Scholar] [CrossRef]
- Zhang, Y.; Zeng, G.; Pan, H.; Li, C.; Hu, Y.; Chu, K.; Han, W.; Chen, Z.; Tang, R.; Yin, W.; et al. Safety, tolerability, and immunogenicity of an inactivated SARS-CoV-2 vaccine in healthy adults aged 18-59 years: A randomised, double-blind, placebo-controlled, phase 1/2 clinical trial. Lancet Infect. Dis. 2021, 21, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Ahi, M.; Farahani, R.H.; Basiri, P.; Rahjerdi, A.K.; Sheidaei, A.; Gohari, K.; Rahimi, Z.; Gholami, F.; Moradi, M.; Naeeni, F.G.; et al. Com-parison of the Safety and Immunogenicity of FAKHRAVAC and BBIBP-CorV Vaccines when Administrated as Booster Dose: A Parallel Two Arms, Randomized, Double Blind Clinical Trial. Vaccines 2022, 10, 1800. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.; Wang, J.; Liu, D.; Xiang, H.; Guo, Y.; Lv, J.; Zhang, Q.; Li, J.; Zhang, X.; Li, Q.; et al. Safety and Immunogenicity of SARS-CoV-2 Vaccines in Patients With Chronic Liver Diseases (CHESS-NMCID 2101): A Multicenter Study. Clin. Gastroenterol. Hepatol. 2022, 20, 1516–1524.e2. [Google Scholar] [CrossRef] [PubMed]
- Jackson, D.A.; Symons, R.H.; Berg, P. Biochemical Method for Inserting New Genetic Information into DNA of Simian Virus 40: Circular SV40 DNA Molecules Containing Lambda Phage Genes and the Galactose Operon of Escherichia coli. Proc. Natl. Acad. Sci. USA 1972, 69, 2904–2909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackett, M.; Smith, G.L.; Moss, B. Vaccinia virus: A selectable eukaryotic cloning and expression vector. Proc. Natl. Acad. Sci. USA 1982, 79, 7415–7419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ura, T.; Okuda, K.; Shimada, M. Developments in Viral Vector-Based Vaccines. Vaccines 2014, 2, 624–641. [Google Scholar] [CrossRef] [Green Version]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell. Microbiol. 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Dal-Ré, R.; Launay, O. Public trust on regulatory decisions: The European Medicines Agency and the AstraZeneca COVID-19 vaccine label. Vaccine 2021, 39, 4029–4031. [Google Scholar] [CrossRef]
- Franchini, M.; Liumbruno, G.M.; Pezzo, M. COVID-19 vaccine-associated immune thrombosis and thrombocytopenia (VITT): Diagnostic and therapeutic recommendations for a new syndrome. Eur. J. Haematol. 2021, 107, 173–180. [Google Scholar] [CrossRef]
- Kelton, J.G.; Arnold, D.M.; Nazy, I. Lessons from vaccine-induced immune thrombotic thrombocytopenia. Nat. Rev. Immunol. 2021, 21, 753–755. [Google Scholar] [CrossRef]
- Gordon, S.F.; Clothier, H.J.; Morgan, H.; Buttery, J.P.; Phuong, L.K.; Monagle, P.; Chunilal, S.; Wood, E.M.; Tran, H.; Szer, J.; et al. Immune thrombocytopenia following immunisation with Vaxzevria ChadOx1-S (AstraZeneca) vaccine, Victoria, Australia. Vaccine 2021, 39, 7052–7057. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.J.; Kao, C.H.; Hsu, T.Y.; Chen, C.Y.; Lin, C.L.; Shih, H.M. Effect of alcohol intoxication on the risk of venous thromboembolism: A nationwide retrospective cohort study. Medicine 2017, 96, e8041. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, A.; Puri, K.; Liangpunsakul, S. Deep vein thrombosis and pulmonary embolism in cirrhotic patients: Systematic review. World J. Gastroenterol. 2014, 20, 5737–5745. [Google Scholar] [CrossRef] [PubMed]
- Pahor, M.; Guralnik, J.M.; Havlik, R.J.; Carbonin, P.; Salive, M.E.; Ferrucci, L.; Corti, M.-C.; Hennekens, C.H. Alcohol Con-sumption and Risk of Deep Venous Thrombosis and Pulmonary Embolism in Older Persons. J. Am. Geriatr. Soc. 1996, 44, 1030–1037. [Google Scholar] [CrossRef]
- Kasuda, S.; Sakurai, Y.; Shima, M.; Morimura, Y.; Kudo, R.; Hatake, K.; Yoshioka, A. Ethanol inhibits microaggregate for-mation of platelets in human whole blood. HAEMA 2004, 7, 200–204. [Google Scholar]
- Abolmaali, M.; Rezania, F.; Behnagh, A.; Hamidabad, N.; Gorji, A.; Mirzaasgari, Z. Guillain-Barré syndrome in asso-ciation with COVID-19 vaccination: A systematic review. Immunol. Res. 2022, 70, 752–764. [Google Scholar] [CrossRef]
- Shay, D.K.; Gee, J.; Su, J.; Myers, T.; Marquez, P.; Liu, R.; Zhang, B.; Licata, C.; Clark, T.; Shimabukuro, T. Safety Monitoring of the Janssen (Johnson & Johnson) COVID-19 Vaccine—United States, March-April 2021. MMWR Morb. Mortal. Wkly. Rep. 2021, 70, 680–684. [Google Scholar]
- Idiculla, P.S.; Gurala, D.; Palanisamy, M.; Vijayakumar, R.; Dhandapani, S.; Nagarajan, E. Cerebral Venous Thrombosis: A Comprehensive Review. Eur. Neurol. 2020, 83, 369–379. [Google Scholar] [CrossRef]
- MacNeil, J.R.; Su, J.R.; Broder, K.R.; Guh, A.Y.; Gargano, J.W.; Wallace, M.; Hadler, S.C.; Scobie, H.M.; Blain, A.E.; Moulia, D.; et al. Updated Recommendations from the Advisory Committee on Immunization Practices for Use of the Janssen (Johnson & Johnson) COVID-19 Vaccine After Reports of Thrombosis with Thrombocytopenia Syndrome Among Vaccine Recipients—United States, April 2021. MMWR Morb. Mortal. Wkly. Rep. 2021, 70, 651–656. [Google Scholar] [CrossRef]
- Oliver, S.E.; Wallace, M.; See, I.; Mbaeyi, S.; Godfrey, M.; Hadler, S.; Jatlaoui, T.; Twentyman, E.; Hughes, M.; Rao, A.; et al. Use of the Janssen (Johnson & Johnson) COVID-19 Vaccine: Updated Interim Rec-ommendations from the Advisory Committee on Immunization Practices—United States, December 2021. MMWR Morb. Mortal. Wkly. Rep. 2022, 71, 90–95. [Google Scholar]
- Logunov, D.Y.; Dolzhikova, I.V.; Zubkova, O.V.; Tukhvatullin, A.I.; Shcheblyakov, D.V.; Dzharullaeva, A.S.; Grousova, D.M.; Erokhova, A.S.; Kovyrshina, A.V.; Botikov, A.G.; et al. Safety and immunogenicity of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine in two formulations: Two open, non-randomised phase 1/2 studies from Russia. Lancet 2020, 396, 887–897. [Google Scholar] [CrossRef]
- Cazzola, M.; Rogliani, P.; Mazzeo, F.; Matera, M.G. Controversy surrounding the Sputnik V vaccine. Respir. Med. 2021, 187, 106569. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.P. Approaches for Optimal Use of Different COVID-19 Vaccines: Issues of Viral Variants and Vaccine Efficacy. JAMA 2021, 325, 1251–1252. [Google Scholar] [CrossRef]
- Perreau, M.; Pantaleo, G.; Kremer, E.J. Activation of a dendritic cell-T cell axis by Ad5 immune complexes creates an im-proved environment for replication of HIV in T cells. J. Exp. Med. 2008, 205, 2717–2725. [Google Scholar] [CrossRef] [PubMed]
- Tumban, E. Lead SARS-CoV-2 Candidate Vaccines: Expectations from Phase III Trials and Recommendations Post-Vaccine Approval. Viruses 2020, 13, 54. [Google Scholar] [CrossRef]
- Heinz, R.; Waltenbaugh, C. Ethanol Consumption Modifies Dendritic Cell Antigen Presentation in Mice. Alcohol Clin. Exp. Res. 2007, 31, 1759–1771. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.G.; Navarro, F.; Chitsike, L.; Ramirez, L.; Kovacs, E.J.; Watkins, S.K. Alcohol exposure differentially effects anti-tumor immunity in females by altering dendritic cell function. Alcohol 2016, 57, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Polina Nikolskaya, A.O.; Reuters. Don’t Mix Sputnik Vaccine with Alcohol, Says Russian Official. Some Recoil. Available online: https://www.reuters.com/article/health-coronavirus-russia-vaccine-alcoho-idUSKBN28J239 (accessed on 19 December 2022).
- Wolff, J.A.; Malone, R.W.; Williams, P.; Chong, W.; Acsadi, G.; Jani, A.; Felgner, P.L. Direct gene transfer into mouse muscle in vivo. Science 1990, 247, 1465–1468. [Google Scholar] [CrossRef]
- Chaudhary, N.; Weissman, D.; Whitehead, K.A. mRNA vaccines for infectious diseases: Principles, delivery and clinical translation. Nat. Rev. Drug Discov. 2021, 20, 817–838. [Google Scholar] [CrossRef]
- Park, J.W.; Lagniton, P.N.; Liu, Y.; Xu, R.-H. mRNA vaccines for COVID-19: What, why and how. Int. J. Biol. Sci. 2021, 17, 1446–1460. [Google Scholar] [CrossRef]
- Chirumbolo, S. Vaccination hesitancy and the “myth” on mRNA-based vaccines in Italy in the COVID-19 era: Does urgency meet major safety criteria? J. Med. Virol. 2021, 93, 4049–4053. [Google Scholar] [CrossRef] [PubMed]
- Trougakos, I.P.; Terpos, E.; Alexopoulos, H.; Politou, M.; Paraskevis, D.; Scorilas, A.; Kastritis, E.; Andreakos, E.; Dimopoulos, M.A. Adverse effects of COVID-19 mRNA vaccines: The spike hypothesis. Trends Mol. Med. 2022, 28, 542–554. [Google Scholar] [CrossRef] [PubMed]
- Ndeupen, S.; Qin, Z.; Jacobsen, S.; Bouteau, A.; Estanbouli, H.; Igyártó, B.Z. The mRNA-LNP platform’s lipid nanoparticle component used in preclinical vaccine studies is highly inflammatory. iScience 2021, 24, 103479. [Google Scholar] [CrossRef]
- Yamamoto, S.; Tanaka, A.; Ohmagari, N.; Yamaguchi, K.; Ishitsuka, K.; Morisaki, N.; Kojima, M.; Nishikimi, A.; Tokuda, H.; Inoue, M.; et al. Use of heated tobacco products, moderate alcohol drinking, and anti-SARS-CoV-2 IgG antibody titers after BNT162b2 vaccination among Japanese healthcare workers. Prev. Med. 2022, 161, 107123. [Google Scholar] [CrossRef]
- Bozkurt, B.; Kamat, I.; Hotez, P.J. Myocarditis With COVID-19 mRNA Vaccines. Circulation 2021, 144, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Patone, M.; Mei, X.W.; Handunnetthi, L.; Dixon, S.; Zaccardi, F.; Shankar-Hari, M.; Watkinson, P.; Khunti, K.; Harnden, A.; Coupland, C.; et al. Risk of Myocarditis After Sequential Doses of COVID-19 Vaccine and SARS-CoV-2 Infection by Age and Sex. Circulation 2022, 146, 743–754. [Google Scholar] [CrossRef]
- Mirijello, A.; Tarli, C.; Vassallo, G.A.; Sestito, L.; Antonelli, M.; D’Angelo, C.; Ferrulli, A.; De Cosmo, S.; Gasbarrini, A.; Addolorato, G. Alcoholic cardiomyopathy: What is known and what is not known. Eur. J. Intern. Med. 2017, 43, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Wilke, A.; Kaiser, A.; Ferency, I.; Maisch, B. Alcohol and myocarditis. Herz 1996, 21, 248–257. [Google Scholar]
- George, A.; Figueredo, V.M. Alcoholic Cardiomyopathy: A Review. J. Card. Fail. 2011, 17, 844–849. [Google Scholar] [CrossRef] [Green Version]
- Awaya, T.; Moroi, M.; Nakamura, F.; Toi, S.; Wakiya, M.; Enomoto, Y.; Kunimasa, T.; Nakamura, M. A Possibility of Vaso-spastic Angina after mRNA COVID-19 Vaccination. Vaccines 2022, 10, 1998. [Google Scholar] [CrossRef]
- Mulligan, M.J.; Lyke, K.E.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Raabe, V.; Bailey, R.; Swanson, K.A.; et al. Phase I/II study of COVID-19 RNA vaccine BNT162b1 in adults. Nature 2020, 586, 589–593. [Google Scholar] [CrossRef] [PubMed]
- John, B.V.; Deng, Y.; Scheinberg, A.; Mahmud, N.; Taddei, T.H.; Kaplan, D.; Labrada, M.; Baracco, G.; Dahman, B. Association of BNT162b2 mRNA and mRNA-1273 Vaccines With COVID-19 Infection and Hospitalization Among Patients With Cirrhosis. JAMA Intern. Med. 2021, 181, 1306–1314. [Google Scholar] [CrossRef] [PubMed]
- COVID-19 Vaccine Tracker. 18 August 2022. Available online: https://www.covid-19vaccinetracker.org/#Top-of-Page (accessed on 19 December 2022).
- Rosenblum, H.G.; Gee, J.; Liu, R.; Marquez, P.L.; Zhang, B.; Strid, P.; Abara, W.E.; McNeil, M.M.; Myers, T.R.; Hause, A.M.; et al. Safety of mRNA vaccines administered during the initial 6 months of the US COVID-19 vaccination programme: An observational study of reports to the Vaccine Adverse Event Reporting System and v-safe. Lancet Infect. Dis. 2022, 22, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.X.; Wu, R.X.; Shen, M.Y.; Huang, J.J.; Li, T.T.; Hu, C.; Luo, F.; Song, S.; Mu, S.; Hao, Y.; et al. Extended SARS-CoV-2 RBD booster vaccination induces humoral and cellular immune tolerance in mice. iScience 2022, 25, 105479. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Vaccine Name | Vaccine Type | Health Conditions and Potential Alcohol-Associated Risks | Reference |
|---|---|---|---|
| Sputnik V | Recombinant adenovirus | Low immune response (warning by health officials) | [99] |
| (Gamaleya National Research Centre for Epidemiology and Microbiology) | Possible reduced efficacy, especially in populations with high anti-Ad5 antibody titers from previous infections | [96] | |
| JNJ-78435735 | Recombinant adenovirus | Thrombosis and thrombocytopenia | [83,84,85,86,87] |
| (Johnson and Johnson/Beth Israel Deaconess Medical Center) | |||
| Covishield, Vaxzevria | Recombinant adenovirus | Thrombosis and thrombocytopenia | [83,84,85,86,87] |
| (Oxford/AstraZeneca) | |||
| BBIBP-CorV | Inactivated whole-virus vaccines | Low immune response | [74] |
| (Sinopharm) | |||
| CoronaVac | Inactivated whole-virus vaccines | Low immune response | [74] |
| (Sinovac) | |||
| NVX-CoV2373 | Recombinant subunit | Low immune response | [22] |
| (Novavax) | High risk of “spike effect” | [21] | |
| BNT162b2 | mRNA | Cardiomyopathy | [109,110,111] |
| (Pfizer/BioNTech) | Low immune response | [106] | |
| Vasospastic angina | [112] | ||
| mRNA-1273 | mRNA | Cardiomyopathy | [107,108,109] |
| (Moderna) | Low immune response | [106] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solopov, P.A. COVID-19 Vaccination and Alcohol Consumption: Justification of Risks. Pathogens 2023, 12, 163. https://doi.org/10.3390/pathogens12020163
Solopov PA. COVID-19 Vaccination and Alcohol Consumption: Justification of Risks. Pathogens. 2023; 12(2):163. https://doi.org/10.3390/pathogens12020163
Chicago/Turabian StyleSolopov, Pavel A. 2023. "COVID-19 Vaccination and Alcohol Consumption: Justification of Risks" Pathogens 12, no. 2: 163. https://doi.org/10.3390/pathogens12020163
APA StyleSolopov, P. A. (2023). COVID-19 Vaccination and Alcohol Consumption: Justification of Risks. Pathogens, 12(2), 163. https://doi.org/10.3390/pathogens12020163