
A Review of Major Patents on Potential Malaria Vaccine Targets

 , ,
, ,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
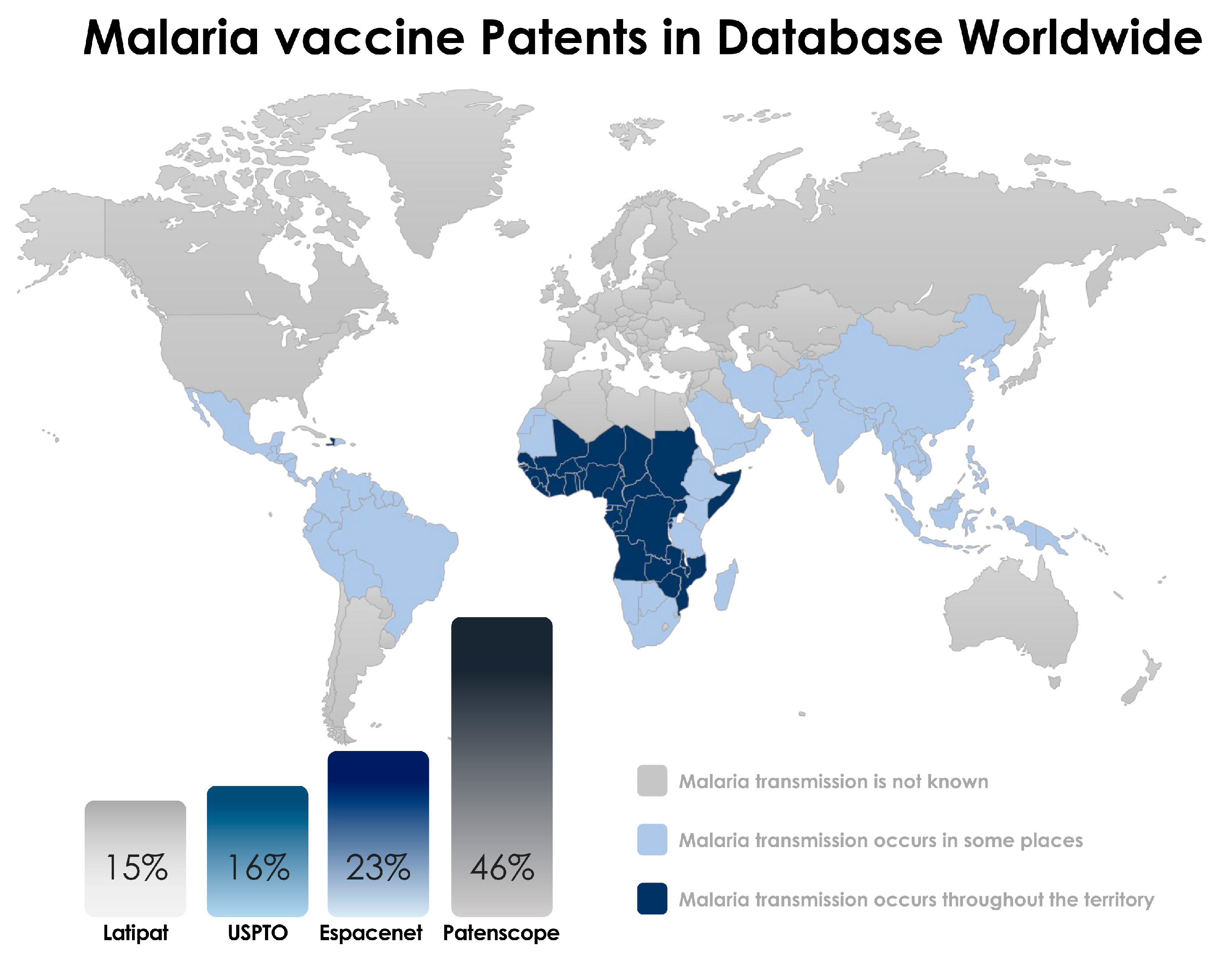
2. Search Strategy and Selection Criteria
3. Results
3.1. Challenge of Developing a Malaria Vaccine
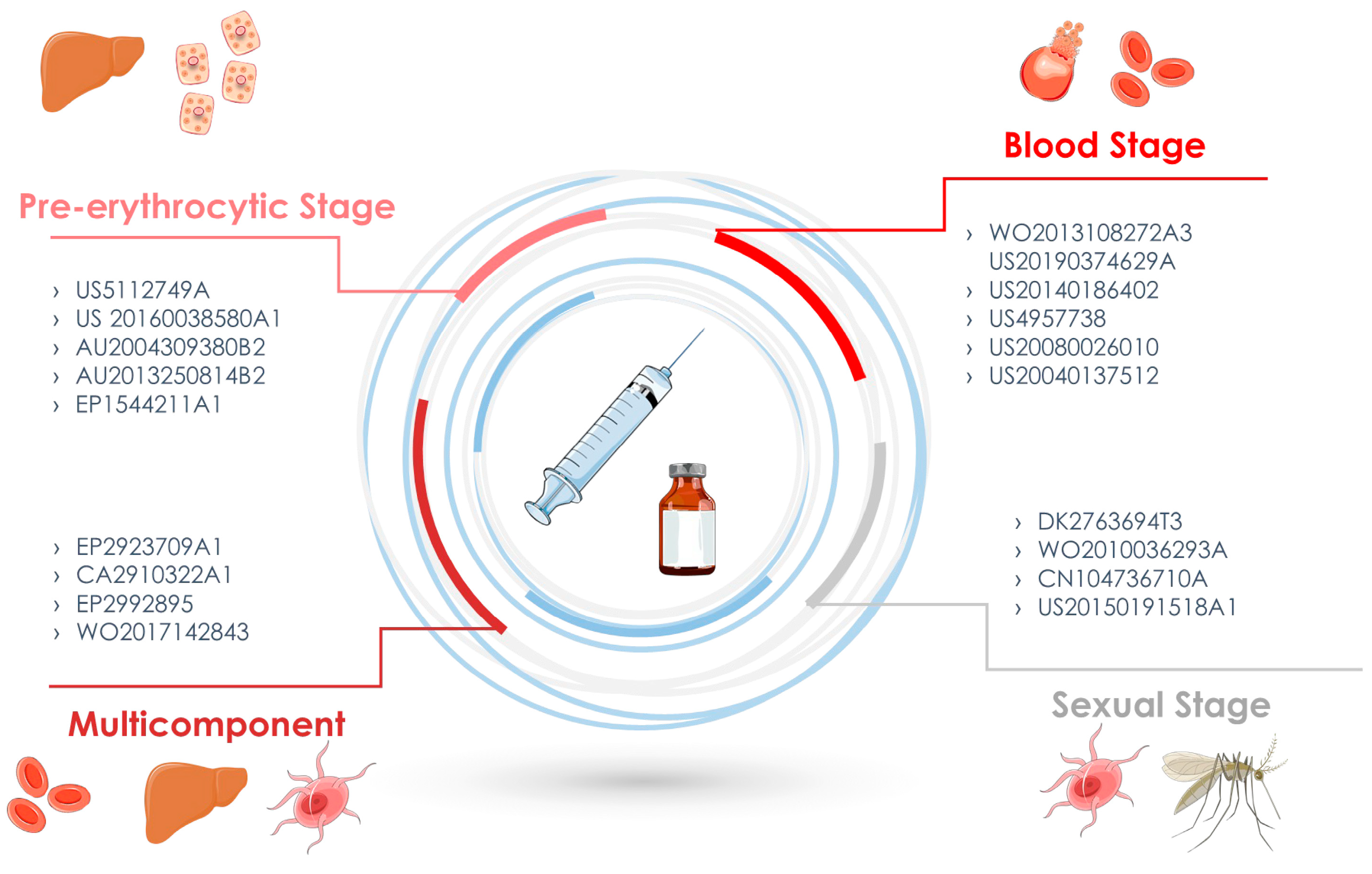
3.2. Pre-Erythrocytic Stage
3.3. Blood Stage
3.4. Sexual Stage
3.5. Multicomponent and/or Various Stage Vaccines
4. New Approaches
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. WORLD MALARIA REPORT. In WHO Library Cataloguing-in-Publication Data; World Health Organization: Geneva, Switzerland, 2018; 210p, ISBN 978 92 4 156565 3. [Google Scholar]
- World Health Organization. WORLD MALARIA REPORT. In WHO Library Cataloguing-in-Publication Data; World Health Organization: Geneva, Switzerland, 2022; 372p, ISBN 978-92-4-006489-8. [Google Scholar]
- Herrera, S.; Corradin, G.; Arévalo-Herrera, M. An update on the search for a Plasmodium vivax vaccine. Trends Parasitol. 2007, 23, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Varo, R.; Chaccour, C.; Bassat, Q. Update on malaria. Med. Clin. 2020, 155, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, S.K.; Childs, L.M.; Buckee, C.; Marti, M. Targeting Human Transmission Biology for Malaria Elimination. PLoS Pathog. 2015, 11, e1004871. [Google Scholar] [CrossRef]
- Meibalan, E.; Marti, M. Biology of Malaria Transmission. Cold Spring Harb. Perspect. Med. 2017, 7, a025452. [Google Scholar] [CrossRef] [PubMed]
- Ramaprasad, A.; Pain, A.; Ravasi, T. Defining the protein interaction network of human malaria parasite Plasmodium falciparum. Genomics 2012, 99, 69–75. [Google Scholar] [CrossRef]
- Coker, M.; Folayan, M.O.; Michelow, I.C.; Oladokun, R.E.; Torbunde, N.; Sam-Agudu, N.A. Things must not fall apart: The ripple effects of the COVID-19 pandemic on children in sub-Saharan Africa. Pediatr. Res. 2021, 89, 1078–1086. [Google Scholar] [CrossRef] [PubMed]
- Sands, P. HIV, tuberculosis, and malaria: How can the impact of COVID-19 be minimised? Lancet Glob. Health 2020, 8, e1102–e1103. [Google Scholar] [CrossRef]
- Gardner, M.J.; Hall, N.; Fung, E.; White, O.; Berriman, M.; Hyman, R.W.; Carlton, J.M.; Pain, A.; Nelson, K.E.; Bowman, S.; et al. Genome sequence of the human malaria parasite Plasmodium falciparum. Nature 2002, 419, 498–511. [Google Scholar] [CrossRef]
- Ballou, W.R. Malaria vaccines in development. Expert Opin. Emerg. Drugs 2005, 10, 489–503. [Google Scholar] [CrossRef]
- Hill, A.V.S. Vaccines against malaria. Phil. Trans. R. Soc. B 2011, 366, 2806–2814. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, C.; Gomes, R. Experimental models in vaccine research: Malaria and leishmaniasis. Braz. J. Med. Biol. Res. 2013, 46, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Ramjanee, S.; Robertson, J.S.; Franke-Fayard, B.; Sinha, R.; Waters, A.P.; Janse, C.J.; Wu, Y.; Blagborough, A.M.; Saul, A.; Sinden, R.E. The use of transgenic Plasmodium berghei expressing the Plasmodium vivax antigen P25 to determine the transmission-blocking activity of sera from malaria vaccine trials. Vaccine 2007, 25, 886–894. [Google Scholar] [CrossRef] [PubMed]
- Vincke, I.H. Experimental transmission of Plasmodium berghei. Indian J. Malariol. 1954, 8, 257–262. [Google Scholar]
- Carlton, J.M.; Hayton, K.; Cravo, P.V.; Walliker, D. Of mice and malaria mutants: Unravelling the genetics of drug resistance using rodent malaria models. Trends Parasitol. 2001, 17, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Stowers, A.W.; Cioce, V.; Shimp, R.L.; Lawson, M.; Hui, G.; Muratova, O.; Kaslow, D.C.; Robinson, R.; Long, C.A.; Miller, L.H. Efficacy of Two Alternate Vaccines Based on Plasmodium falciparum Merozoite Surface Protein 1 in an Aotus Challenge Trial. Infect. Immun. 2001, 69, 1536–1546. [Google Scholar] [CrossRef]
- Webster, K.; Collins, W.E.; Aikawa, M.; Brown, A.; Smith, C.D.; Tegoshi, T.; Howard, R.J.; Hasler, T.H.; Ito, Y.; Perry, G.; et al. A Primate Model for Human Cerebral Malaria: Plasmodium coatneyi-Infected Rhesus Monkeys. Am. J. Trop. Med. Hyg. 1992, 46, 391–397. [Google Scholar] [CrossRef]
- Frimpong, A.; Kusi, K.A.; Ofori, M.F.; Ndifon, W. Novel Strategies for Malaria Vaccine Design. Front. Immunol. 2018, 9, 2769. [Google Scholar] [CrossRef]
- Bojang, K.A.; Milligan, P.J.; Pinder, M.; Vigneron, L.; Alloueche, A.; Kester, K.E.; Ballou, W.R.; Conway, D.J.; Reece, W.H.; Gothard, P.; et al. Efficacy of RTS,S/AS02 malaria vaccine against Plasmodium falciparum infection in semi-immune adult men in The Gambia: A randomised trial. Lancet 2001, 358, 1927–1934. [Google Scholar] [CrossRef] [PubMed]
- Aponte, J.J.; Aide, P.; Renom, M.; Mandomando, I.; Bassat, Q.; Sacarlal, J.; Manaca, M.N.; Lafuente, S.; Barbosa, A.; Leach, A.; et al. Safety of the RTS, S/AS02D candidate malaria vaccine in infants living in a highly endemic area of Mozambique: A double blind randomised controlled phase I/IIb trial. Lancet 2007, 370, 1543–1551. [Google Scholar] [CrossRef]
- Olotu, A.; Fegan, G.; Wambua, J.; Nyangweso, G.; Leach, A.; Lievens, M.; Kaslow, D.C.; Njuguna, P.; Marsh, K.; Bejon, P. Seven-Year Efficacy of RTS,S/AS01 Malaria Vaccine among Young African Children. N. Engl. J. Med. 2016, 374, 2519–2529. [Google Scholar] [CrossRef]
- Adepoju, P. RTS, S malaria vaccine pilots in three African countries. Lancet 2019, 393, 1685. [Google Scholar] [CrossRef] [PubMed]
- Crompton, P.D.; Pierce, S.K.; Miller, L.H. Advances and challenges in malaria vaccine development. J. Clin. Investig. 2010, 120, 4168–4178. [Google Scholar] [CrossRef] [PubMed]
- Suau, R.; Vidal, M.; Aguilar, R.; Ruiz-Olalla, G.; Vázquez-Santiago, M.; Jairoce, C.; Nhabomba, A.J.; Gyan, B.; Dosoo, D.; Asante, K.P.; et al. RTS,S/AS01E malaria vaccine induces IgA responses against CSP and vaccine-unrelated antigens in African children in the phase 3 trial. Vaccine 2021, 39, 687–698. [Google Scholar] [CrossRef]
- Dobaño, C.; Ubillos, I.; Jairoce, C.; Gyan, B.; Vidal, M.; Jiménez, A.; Santano, R.; Dosoo, D.; Nhabomba, A.J.; Ayestaran, A.; et al. RTS,S/AS01E Immunization Increases Antibody Responses to Vaccine-Unrelated Plasmodium Falciparum Antigens Associated with Protection against Clinical Malaria in African Children: A Case-Control Study. BMC Med. 2019, 17, 157. [Google Scholar] [CrossRef]
- Duffy, P.E.; Patrick Gorres, J. Malaria vaccines since 2000: Progress, priorities, products. Npj Vaccines 2020, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Sanaria–Foundation for Mission Statement. Available online: http://www.sanaria.com/index.php?s=38 (accessed on 27 June 2021).
- Hoffman, S.L.; Billingsley, P.F.; James, E.; Richman, A.; Loyevsky, M.; Li, T.; Chakravarty, S.; Gunasekera, A.; Chattopadhyay, R.; Li, M.; et al. Development of a metabolically active, non-replicating sporozoite vaccine to prevent Plasmodium falciparum malaria. Hum. Vaccines 2010, 6, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Epstein, J.E.; Tewari, K.; Lyke, K.E.; Sim, B.K.L.; Billingsley, P.F.; Laurens, M.B.; Gunasekera, A.; Chakravarty, S.; James, E.R.; Sedegah, M.; et al. Live Attenuated Malaria Vaccine Designed to Protect Through Hepatic CD8+ T Cell Immunity. Science 2011, 334, 475–480. [Google Scholar] [CrossRef]
- Plotkin, S.A. Vaccines: Correlates of Vaccine-Induced Immunity. Clin. Infect. Dis. 2008, 47, 401–409. [Google Scholar] [CrossRef]
- McNamara, H.A.; Idris, A.H.; Sutton, H.J.; Vistein, R.; Flynn, B.J.; Cai, Y.; Wiehe, K.; Lyke, K.E.; Chatterjee, D.; Kc, N.; et al. Antibody Feedback Limits the Expansion of B Cell Responses to Malaria Vaccination but Drives Diversification of the Humoral Response. Cell Host Microbe 2020, 28, 572–585.e7. [Google Scholar] [CrossRef]
- Coelho, C.H.; Duffy, P.E. Unwanted Feedback: Malaria Antibodies Hinder Vaccine Boosting. Cell Host Microbe 2020, 28, 504–506. [Google Scholar] [CrossRef]
- Moon, J.E.; Ockenhouse, C.; Regules, J.A.; Vekemans, J.; Lee, C.; Chuang, I.; Traskine, M.; Jongert, E.; Ivinson, K.; Morelle, D.; et al. A Phase IIa Controlled Human Malaria Infection and Immunogenicity Study of RTS,S/AS01E and RTS,S/AS01B Delayed Fractional Dose Regimens in Malaria-Naive Adults. J. Infect. Dis. 2020, 222, 1681–1691. [Google Scholar] [CrossRef] [PubMed]
- Regules, J.A.; Cicatelli, S.B.; Bennett, J.W.; Paolino, K.M.; Twomey, P.S.; Moon, J.E.; Kathcart, A.K.; Hauns, K.D.; Komisar, J.L.; Qabar, A.N.; et al. Fractional Third and Fourth Dose of RTS,S/AS01 Malaria Candidate Vaccine: A Phase 2a Controlled Human Malaria Parasite Infection and Immunogenicity Study. J. Infect. Dis. 2016, 214, 762–771. [Google Scholar] [CrossRef] [PubMed]
- Datoo, M.S.; Natama, H.M.; Somé, A.; Bellamy, D.; Traoré, O.; Rouamba, T.; Tahita, M.C.; Ido, N.F.A.; Yameogo, P.; Valia, D.; et al. Efficacy and Immunogenicity of R21/Matrix-M Vaccine against Clinical Malaria after 2 Years’ Follow-up in Children in Burkina Faso: A Phase 1/2b Randomised Controlled Trial. Lancet Infect. Dis. 2022, 22, 1728–1736. [Google Scholar] [CrossRef]
- Vaughan, A.M.; Aly, A.S.I.; Kappe, S.H.I. Malaria Parasite Pre-Erythrocytic Stage Infection: Gliding and Hiding. Cell Host Microbe 2008, 4, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Roestenberg, M.; McCall, M.; Hopman, J.; Wiersma, J.; Luty, A.J.F.; van Gemert, G.J.; van de Vegte-Bolmer, M.; van Schaijk, B.; Teelen, K.; Arens, T.; et al. Protection against a malaria challenge by sporozoite inoculation. N. Engl. J. Med. 2009, 361, 468–477. [Google Scholar] [CrossRef]
- Abuga, K.M.; Jones-Warner, W.; Hafalla, J.C.R. Immune responses to malaria pre-erythrocytic stages: Implications for vaccine development. Parasite Immunol. 2021, 43, e12795. [Google Scholar] [CrossRef] [PubMed]
- Fidock, D.A.; Pasquetto, V.; Gras, H.; Badell, E.; Eling, W.; Ballou, W.R.; Belghiti, J.; Tartar, A.; Druilhe, P. Plasmodium falciparum sporozoite invasion is inhibited by naturally acquired or experimentally induced polyclonal antibodies to the STARP antigen. Eur. J. Immunol. 1997, 27, 2502–2513. [Google Scholar] [CrossRef]
- John, C.C.; Tande, A.J.; Moormann, A.M.; Sumba, P.O.; Lanar, D.E.; Min, X.M.; Kazura, J.W. Antibodies to Pre-erythrocytic Plasmodium falciparum Antigens and Risk of Clinical Malaria in Kenyan Children. J. Infect. Dis. 2008, 197, 519–526. [Google Scholar] [CrossRef]
- Reece, W.H.H.; Pinder, M.; Gothard, P.K.; Milligan, P.; Bojang, K.; Doherty, T.; Plebanski, M.; Akinwunmi, P.; Everaere, S.; Watkins, K.R.; et al. A CD4+ T-cell immune response to a conserved epitope in the circumsporozoite protein correlates with protection from natural Plasmodium falciparum infection and disease. Nat. Med. 2004, 10, 406–410. [Google Scholar] [CrossRef]
- White, M.T.; Bejon, P.; Olotu, A.; Griffin, J.T.; Riley, E.M.; Kester, K.E.; Ockenhouse, C.F.; Ghani, A.C. The Relationship between RTS,S Vaccine-Induced Antibodies, CD4+ T Cell Responses and Protection against Plasmodium falciparum Infection. PLoS ONE 2013, 8, e61395. [Google Scholar] [CrossRef]
- Weiss, W.R.; Jiang, C.G. Protective CD8+ T lymphocytes in Primates Immunized with Malaria Sporozoites. PLoS ONE 2012, 7, e31247. [Google Scholar] [CrossRef] [PubMed]
- Tarun, A.S.; Dumpit, R.F.; Camargo, N.; Labaied, M.; Liu, P.; Takagi, A.; Wang, R.; Kappe, S.H.I. Protracted Sterile Protection with Plasmodium yoelii Pre-erythrocytic Genetically Attenuated Parasite Malaria Vaccines Is Independent of Significant Liver-Stage Persistence and Is Mediated by CD8 + T Cells. J. Infect. Dis. 2007, 196, 608–616. [Google Scholar] [CrossRef]
- Oakley, M.S.; Verma, N.; Zheng, H.; Anantharaman, V.; Takeda, K.; Gao, Y.; Myers, T.G.; Pham, P.T.; Mahajan, B.; Kumar, N.; et al. Molecular Markers of Radiation Induced Attenuation in Intrahepatic Plasmodium falciparum Parasites. PLoS ONE 2016, 11, e0166814. [Google Scholar] [CrossRef]
- Annoura, T.; Ploemen, I.H.J.; van Schaijk, B.C.L.; Sajid, M.; Vos, M.W.; van Gemert, G.-J.; Chevalley-Maurel, S.; Franke-Fayard, B.M.D.; Hermsen, C.C.; Gego, A.; et al. Assessing the adequacy of attenuation of genetically modified malaria parasite vaccine candidates. Vaccine 2012, 30, 2662–2670. [Google Scholar] [CrossRef] [PubMed]
- Sisquella, X.; Nebl, T.; Thompson, J.K.; Whitehead, L.; Malpede, B.M.; Salinas, N.D.; Rogers, K.; Tolia, N.H.; Fleig, A.; O’Neill, J.; et al. Plasmodium falciparum ligand binding to erythrocytes induce alterations in deformability essential for invasion. eLife 2017, 6, e21083. [Google Scholar] [CrossRef]
- Chen, L.; Lopaticki, S.; Riglar, D.T.; Dekiwadia, C.; Uboldi, A.D.; Tham, W.-H.; O’Neill, M.T.; Richard, D.; Baum, J.; Ralph, S.A.; et al. An EGF-like Protein Forms a Complex with PfRh5 and Is Required for Invasion of Human Erythrocytes by Plasmodium falciparum. PLoS Pathog. 2011, 7, e1002199. [Google Scholar] [CrossRef] [PubMed]
- Campeotto, I.; Goldenzweig, A.; Davey, J.; Barfod, L.; Marshall, J.M.; Silk, S.E.; Wright, K.E.; Draper, S.J.; Higgins, M.K.; Fleishman, S.J. One-Step Design of a Stable Variant of the Malaria Invasion Protein RH5 for Use as a Vaccine Immunogen. Proc. Natl. Acad. Sci. USA 2017, 114, 998–1002. [Google Scholar] [CrossRef]
- El Bissati, K.; Zufferey, R.; Witola, W.H.; Carter, N.S.; Ullman, B.; Ben Mamoun, C. The plasma membrane permease PfNT1 is essential for purine salvage in the human malaria parasite Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 2006, 103, 9286–9291. [Google Scholar] [CrossRef]
- Graciano, R.C.D.; Ribeiro, J.A.T.; Macêdo, A.K.S.; de SLavareda, J.P.; de Oliveira, P.R.; Netto, J.B.; Nogueira, L.M.; Machado, J.M.; Camposda-Paz, M.; Giunchetti, R.C.; et al. Recent Patents Applications in Red Biotechnology: A Mini-Review. BIOT 2019, 13, 170–186. [Google Scholar] [CrossRef]
- Sauerwein, R.W.; Eling, W.M.C. Sexual and Sporogonic Stage Antigens. In Chemical Immunology and Allergy; Perlmann, P., Troye-Blomberg, M., Eds.; KARGER: Basel, Switzerland, 2002; Volume 80, pp. 188–203. [Google Scholar] [CrossRef]
- Saxena, A.K.; Wu, Y.; Garboczi, D.N. Plasmodium P25 and P28 Surface Proteins: Potential Transmission-Blocking Vaccines. Eukaryot. Cell 2007, 6, 1260–1265. [Google Scholar] [CrossRef] [Green Version]
- Pradel, G. Proteins of the malaria parasite sexual stages: Expression, function and potential for transmission blocking strategies. Parasitology 2007, 134, 1911–1929. [Google Scholar] [CrossRef] [PubMed]
- Coutinho-Abreu, I.V.; Ramalho-Ortigao, M. Transmission blocking vaccines to control insect-borne diseases: A review. Mem. Inst. Oswaldo Cruz. 2010, 105, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Eksi, S.; Czesny, B.; van Gemert, G.-J.; Sauerwein, R.W.; Eling, W.; Williamson, K.C. Malaria transmission-blocking antigen, Pfs230, mediates human red blood cell binding to exflagellating male parasites and oocyst production. Mol. Microbiol. 2006, 61, 991–998. [Google Scholar] [CrossRef]
- Nikolaeva, D.; Draper, S.J.; Biswas, S. Toward the development of effective transmission-blocking vaccines for malaria. Expert Rev. Vaccines 2015, 14, 653–680. [Google Scholar] [CrossRef]
- Carter, R. Transmission blocking malaria vaccines. Vaccine 2001, 19, 2309–2314. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, M.R.; Janse, C.J.; Thompson, J.; Waters, A.P.; Braks, J.A.; Dodemont, H.J.; Stunnenberg, H.G.; van Gemert, G.-J.; Sauerwein, R.W.; Eling, W. A central role for P48/45 in malaria parasite male gamete fertility. Cell 2001, 104, 153–164. [Google Scholar] [CrossRef]
- Roeffen, W.; Teelen, K.; van As, J.; vd Vegte-Bolmer, M.; Eling, W.; Sauerwein, R. Plasmodium falciparum: Production and Characterization of Rat Monoclonal Antibodies Specific for the Sexual-Stage Pfs48/45 Antigen. Exp. Parasitol. 2001, 97, 45–49. [Google Scholar] [CrossRef]
- Simon, N.; Lasonder, E.; Scheuermayer, M.; Kuehn, A.; Tews, S.; Fischer, R.; Zipfel, P.F.; Skerka, C.; Pradel, G. Malaria Parasites Co-opt Human Factor H to Prevent Complement-Mediated Lysis in the Mosquito Midgut. Cell Host Microbe 2013, 13, 29–41. [Google Scholar] [CrossRef]
- Talaat, K.R.; Ellis, R.D.; Hurd, J.; Hentrich, A.; Gabriel, E.; Hynes, N.A.; Rausch, K.M.; Zhu, D.; Muratova, O.; Herrera, R.; et al. Safety and Immunogenicity of Pfs25-EPA/Alhydrogel®, a Transmission Blocking Vaccine against Plasmodium falciparum: An Open Label Study in Malaria Naïve Adults. PLoS ONE 2016, 11, e0163144. [Google Scholar] [CrossRef] [PubMed]
- Sagara, I.; Healy, S.A.; Assadou, M.H.; Gabriel, E.E.; Kone, M.; Sissoko, K.; Tembine, I.; Guindo, M.A.; Doucoure, M.; Niaré, K.; et al. Safety and immunogenicity of Pfs25H-EPA/Alhydrogel, a transmission-blocking vaccine against Plasmodium falciparum: A randomised, double-blind, comparator-controlled, dose-escalation study in healthy Malian adults. Lancet Infect. Dis. 2018, 18, 969–982. [Google Scholar] [CrossRef]
- Coelho, C.H.; Rappuoli, R.; Hotez, P.J.; Duffy, P.E. Transmission-Blocking Vaccines for Malaria: Time to Talk about Vaccine Introduction. Trends Parasitol. 2019, 35, 483–486. [Google Scholar] [CrossRef]
- Standaert, B.; Rappuoli, R. 3. How comprehensive can we be in the economic assessment of vaccines? J. Mark. Access Health Policy 2017, 5, 1336044. [Google Scholar] [CrossRef] [PubMed]
- Duffy, P.E.; Sahu, T.; Akue, A.; Milman, N.; Anderson, C. Pre-erythrocytic malaria vaccines: Identifying the targets. Expert Rev. Vaccines 2012, 11, 1261–1280. [Google Scholar] [CrossRef]
- Belachew, E.B. Immune Response and Evasion Mechanisms of Plasmodium falciparum Parasites. J. Immunol. Res. 2018, 2018, 6529681. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.-C.; Shih, T.-P.; Ko, W.-C.; Tang, H.-J.; Hsueh, P.-R. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and coronavirus disease-2019 (COVID-19): The epidemic and the challenges. Int. J. Antimicrob. Agents 2020, 55, 105924. [Google Scholar] [CrossRef] [PubMed]
- Singh, A. Eliciting B cell immunity against infectious diseases using nanovaccines. Nat. Nanotechnol. 2021, 16, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.S.; Hirosue, S.; Raczy, M.M.; Bonilla-Ramirez, L.; Jeanbart, L.; Wang, R.; Kwissa, M.; Franetich, J.-F.; Broggi, M.A.S.; Diaceri, G.; et al. Antigens reversibly conjugated to a polymeric glyco-adjuvant induce protective humoral and cellular immunity. Nat. Mater. 2019, 18, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Kanoi, B.N.; Nagaoka, H.; Morita, M.; Tsuboi, T.; Takashima, E. Leveraging the wheat germ cell-free protein synthesis system to accelerate malaria vaccine development. Parasitol. Int. 2021, 80, 102224. [Google Scholar] [CrossRef]
- Tsuboi, T.; Takeo, S.; Iriko, H.; Jin, L.; Tsuchimochi, M.; Matsuda, S.; Han, E.-T.; Otsuki, H.; Kaneko, O.; Sattabongkot, J.; et al. Wheat Germ Cell-Free System-Based Production of Malaria Proteins for Discovery of Novel Vaccine Candidates. Infect. Immun. 2008, 76, 1702–1708. [Google Scholar] [CrossRef]
- Rui, E.; Fernandez-Becerra, C.; Takeo, S.; Sanz, S.; Lacerda, M.V.; Tsuboi, T.; del Portillo, H.A. Plasmodium vivax: Comparison of immunogenicity among proteins expressed in the cell-free systems of Escherichia coli and wheat germ by suspension array assays. Malar. J. 2011, 10, 192. [Google Scholar] [CrossRef] [Green Version]
- Jaroentomeechai, T.; Stark, J.C.; Natarajan, A.; Glasscock, C.J.; Yates, L.E.; Hsu, K.J.; Mrksich, M.; Jewett, M.C.; DeLisa, M.P. Single-pot glycoprotein biosynthesis using a cell-free transcription-translation system enriched with glycosylation machinery. Nat. Commun. 2018, 9, 2686. [Google Scholar] [CrossRef] [PubMed]
- Avci, F.Y.; Li, X.; Tsuji, M.; Kasper, D.L. Carbohydrates and T cells: A sweet twosome. Semin. Immunol. 2013, 25, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Goddard-Borger, E.D.; Boddey, J.A. Implications of Plasmodium glycosylation on vaccine efficacy and design. Future Microbiol. 2018, 13, 609–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
| Registration Number | Invention Name | Inventors | Target Stage | Target Antigen | Database | Patent Date |
|---|---|---|---|---|---|---|
| US5112749A | Vaccines for the malaria circumsporozoite protein | Brey, Robert et al. | Pre-erythrocytic stage | Circumsporozoite recombinant proteins | USPTO, Espacenet | 12 May 1992 |
| US20160038580A1 | Soluble recombinant P. falciparum circumsporozoite protein, use in vaccines, production methods and uses | Dutta, Sheetij | Pre-erythrocytic stage | Circumsporozoite recombinant proteins | USPTO | 11 February 2016 |
| AU2004309380B2 | Live genetically attenuated malaria vaccine | Stefan, Kappe, et al. | Pre-erythrocytic stage | Live Plasmodium organisms genetically engineered | Espacenet | 29 April 2010 |
| AU2013250814B2 | Rodent Plasmodium parasites as platforms for a whole-organism malaria vaccine | Mendes, Antonio Manuel Barbeiro et al. | Pre-erythrocytic stage | Live Plasmodium organisms genetically engineered | Espacenet | 2 March 2017 |
| EP1544211A1 | LSA-5 pre-erythrocytic stage antigen of P. falciparum, an immunogenic composition comprising said antigen, and vaccines against malaria | Brahimi-Zeghidour, Karima and Druilhe, Pierre | Pre-erythrocytic stage | P. falciparum liver sporozoite antigen | Espacenet | 22 Jun 2005 |
| WO2013108272A3 | Blood-stage malaria vaccine | Gaur, Deepak, et al. | Blood stage | Merozoite antigens | Patent scope, Espacenet | 25 July 2013 |
| US20190374629A | Thermostable variants of P. Falciparum PfRH5 that can be produced in bacterial cells | Draper, Simon, et al. | Blood stage | Modified homologous reticulocyte binding | USPTO, espacenet | 12 December 2019 |
| US20140186402A1 | Blood Stage Malaria Vaccine | Good, Michael Spithill, Terry Lee, Moses | Blood stage | Isolated or purified merozoites | USPTO, espacenet | 3 July 2014 |
| US20080026010 | Use of conditional Plasmodium strains lacking nutrient transporters in malaria vaccination | Ben Mamoun Choukri, El Bissati Kamal | Blood stage | Attenuated malarial parasite | Patent scope, Espacenet, USPTO | 31 January 2008 |
| US4957738 | Protein copolymer malaria vaccine | Patarroyo, Manuel | Blood stage | Circumsporozoite proteins synthetic | USPTO, espacenet | 18 September 1990 |
| US20040137512 | Malaria Plasmodium antigen polypeptide SE36, method of purifying the same and vaccine and diagnosis with the use of the resulting antigen | Horii, Toshihiro | Blood stage | Serine-repeat antigen of P. falciparum | Patent scope, Espacenet, USPTO, Latipat | 15 July 2004 |
| DK2763694T3 | Preparation of a cysteine-rich protein | Andersen, Michael TheisenGorm | Sexual stage | Pfs48/45 recombinant protein | espacenet | 16 April 2018 |
| WO2010036293A1 | Malaria vaccine | Kumar, Nirbhay Angov, Evelina | Sexual stage | Pfs48/45 recombinant protein | Patent scope, Espacenet, | 3 June 2010 |
| CN104736710A | Use of p47 from P. falciparum (pfs47) or P. vivax (pvs47) as a vaccine or drug screening target for the inhibition of human malaria transmission | Barillas-Mury, Carolina Veronica et al. | Sexual stage | P. falciparum P47 (Pfs47) or P. vivax P47 (Pfs47) surface antigens | espacenet | 24 June 2015 |
| US20150191518A1 | Novel malaria transmission-blocking vaccines | Pradel, Gabriele et al. | Sexual stage | P. falciparum gliding-associated protein 50 | USPTO, espacenet | 9 July 2015 |
| EP2923709A1 | Multi-component-multistage malaria vaccine | Boes, Alexander et al. | Various Stage Vaccines | Recombinant fusion proteins | Espacenet | 20 September 2015 |
| CA2910322A1 | Novel vaccines against apicomplexan pathogens | Boes, Alexander et al. | Various Stage Vaccines | Recombinant fusion proteins | Espacenet | 20 October 2014 |
| EP2992895 | Three-component-multistage malaria vaccine | Fischer, Rainer et al. | Various Stage Vaccines | Recombinant fusion proteins | Espacenet | 9 March 2016 |
| WO2017142843 | Novel antigen for use in malaria vaccine | AGUIAR, Joao Carlos | Various Stage Vaccines | Recombinant fusion proteins | Patent scope | 24 August 2017 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mariano, R.M.d.S.; Gonçalves, A.A.M.; Oliveira, D.S.d.; Ribeiro, H.S.; Pereira, D.F.S.; Santos, I.S.; Lair, D.F.; Silva, A.V.d.; Galdino, A.S.; Chávez-Fumagalli, M.A.; et al. A Review of Major Patents on Potential Malaria Vaccine Targets. Pathogens 2023, 12, 247. https://doi.org/10.3390/pathogens12020247
Mariano RMdS, Gonçalves AAM, Oliveira DSd, Ribeiro HS, Pereira DFS, Santos IS, Lair DF, Silva AVd, Galdino AS, Chávez-Fumagalli MA, et al. A Review of Major Patents on Potential Malaria Vaccine Targets. Pathogens. 2023; 12(2):247. https://doi.org/10.3390/pathogens12020247
Chicago/Turabian StyleMariano, Reysla Maria da Silveira, Ana Alice Maia Gonçalves, Diana Souza de Oliveira, Helen Silva Ribeiro, Diogo Fonseca Soares Pereira, Ingrid Soares Santos, Daniel Ferreira Lair, Augusto Ventura da Silva, Alexsandro Sobreira Galdino, Miguel Angel Chávez-Fumagalli, and et al. 2023. "A Review of Major Patents on Potential Malaria Vaccine Targets" Pathogens 12, no. 2: 247. https://doi.org/10.3390/pathogens12020247
APA StyleMariano, R. M. d. S., Gonçalves, A. A. M., Oliveira, D. S. d., Ribeiro, H. S., Pereira, D. F. S., Santos, I. S., Lair, D. F., Silva, A. V. d., Galdino, A. S., Chávez-Fumagalli, M. A., Silveira-Lemos, D. d., Dutra, W. O., & Giunchetti, R. C. (2023). A Review of Major Patents on Potential Malaria Vaccine Targets. Pathogens, 12(2), 247. https://doi.org/10.3390/pathogens12020247