

The Humanized Mouse Model: What Added Value Does It Offer for HIV Research?

{kind=link}
Abstract
:1. Introduction
2. What Are the Major Questions in HIV Research?
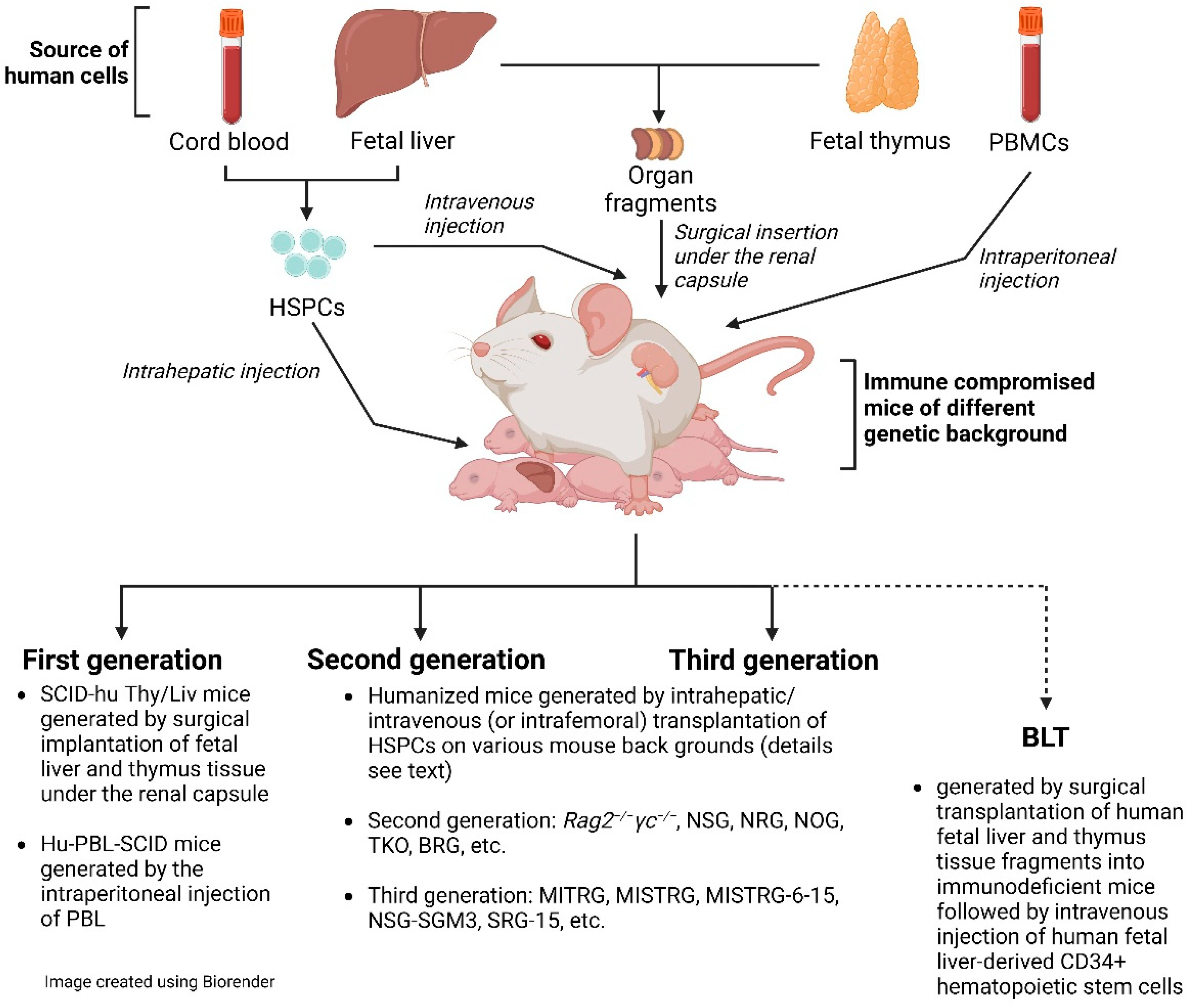
3. What Models Are Available for Biomedical HIV Research?
4. What Are the Attributes of a Good HIV Mouse Model?
5. Of All the Novel Models Reported, Is There a Model That Should Preferably Be Used to Study HIV Infection?
5.1. Ad: Which Humanized Mouse Models Are Suitable for Testing Novel anti-HIV Compounds and anti-HIV bNAbs?
5.2. Ad: Development of Cell and Gene Therapy to Cure HIV
5.3. Ad: Exploring Experimental Strategies to Eradicate Dormant HIV
5.4. Ad: Prophylactic or Therapeutic Vaccination
5.5. Ad: HIV Infection via the Vaginal Route
5.6. Ad: Pathogenesis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AIDS | Acquired immunodeficiency syndrome |
| ART | Antiretroviral therapy |
| BLT | Bone marrow thymus liver |
| bNAbs | Broadly neutralizing antibodies |
| BRG | Rag2−/−/IL-2Rγ−/− in BALB/c background |
| cART | Combined antiretroviral therapy |
| CCR5 | C-C chemokine receptor 5 |
| COVID-19 | Coronavirus disease 19 |
| CXCR4 | C-X-C chemokine receptor type 4 |
| DARPins | Designed ankyrin repeat proteins |
| DMPA | Depot medroxyprogesterone acetate |
| Env | Envelope protein |
| FITC | Fluorescein isothiocyanate |
| GM-CSF | Granulocyte–macrophage colony stimulating factor |
| GvHD | Graft versus host disease |
| HIV | Human immunodeficiency virus |
| HLA | Human leukocyte antigen |
| HSPCs | Hematopoietic stem and progenitor cells |
| hu | Humanized |
| Hu-PBL-SCID | Humanized peripheral blood leukocytes severe combined Immunodeficiency |
| IgA | Immunoglobulin A |
| IgG | Immunoglobulin G |
| IL | Interleukin |
| MHC | Major histocompatibility complex |
| MISTRG | M-CSFh/h IL-3/GM-CSFh/h SIRP-αh/m TPOh/h RAG2−/−IL-2Rγ−/− |
| MITRG | M-CSFh/h IL-3/GM-CSFh/h TPOh/h RAG2−/−IL-2Rγ−/− |
| NK cells | Natural killer cells |
| NOD | Non-obese diabetic |
| NRG | Rag1−/−/IL-2Rγ−/− in NOD background |
| NSG | SCID/IL-2Rγ−/− in NOD background |
| NSG-SGM3 | NSG-Human Stem cell factor–GM-CSF–IL-3 |
| NYVAC-KC | Replication-competent, attenuated recombinant of the vaccinia virus strain NYVAC |
| PBMCs | Peripheral blood mononuclear cells |
| PD-1 | Programmed cell death protein-1 |
| Rag2−/−γc−/− | Rag2/Il2rγ compound mutant mice |
| RNA | Ribonucleic acid |
| SCID-hu Thy/Liv | Severe combined immunodeficiency–human thymus and liver |
| SIRP-α | Signal recognition protein alpha |
| SIV | Simian immunodeficiency virus |
| SRG-15 | Human SIRP-α and human IL-15 knock-in |
| TKO | Triple knockout |
| TLR | Toll-like receptor |
| UNAIDS | Joined United Nations Programme on HIV/AIDS |
References
- Campbell, M.; Gandhi, R.T. Update on Human Immunodeficiency Virus (HIV)-2 Infection. Clin. Infect. Dis. 2011, 52, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Veazey, R.S.; Lackner, A.A. Nonhuman Primate Models and Understanding the Pathogenesis of HIV Infection and AIDS. ILAR J. 2017, 58, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Nischang, M.; Gers-Huber, G.; Audigé, A.; Akkina, R.; Speck, R.F. Modeling HIV infection and therapies in humanized mice. Swiss Med. Wkly. 2012, 142, w13618. [Google Scholar] [CrossRef] [PubMed]
- Martinov, T.; McKenna, K.M.; Tan, W.H.; Collins, E.J.; Kehret, A.R.; Linton, J.D.; Olsen, T.M.; Shobaki, N.; Rongvaux, A. Building the Next Generation of Humanized Hemato-Lymphoid System Mice. Front. Immunol. 2021, 12, 643852. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liao, S.; Xiao, Z.; Pan, Q.; Wang, X.; Shen, K.; Wang, S.; Yang, L.; Guo, F.; Liu, H.-F.; et al. The development and improvement of immunodeficient mice and humanized immune system mouse models. Front. Immunol. 2022, 13, 7579. [Google Scholar] [CrossRef]
- Ye, W.; Chen, Q. Potential Applications and Perspectives of Humanized Mouse Models. Annu. Rev. Anim. Biosci. 2022, 10, 395–417. [Google Scholar] [CrossRef]
- Terahara, K.; Iwabuchi, R.; Tsunetsugu-Yokota, Y. Perspectives on Non-BLT Humanized Mouse Models for Studying HIV Pathogenesis and Therapy. Viruses 2021, 13, 776. [Google Scholar] [CrossRef]
- Saag, M.S.; Gandhi, R.T.; Hoy, J.F.; Landovitz, R.J.; Thompson, M.A.; Sax, P.E.; Smith, D.M.; Benson, C.A.; Buchbinder, S.P.; Del Rio, C.; et al. Antiretroviral Drugs for Treatment and Prevention of HIV Infection in Adults. JAMA 2020, 324, 1651–1669. [Google Scholar] [CrossRef]
- Mediouni, S.; Lyu, S.; Schader, S.M.; Valente, S.T. Forging a Functional Cure for HIV: Transcription Regulators and Inhibitors. Viruses 2022, 14, 1980. [Google Scholar] [CrossRef]
- Plückthun, A. Designed Ankyrin Repeat Proteins (DARPins): Binding Proteins for Research, Diagnostics, and Therapy. Annu. Rev. Pharmacol. Toxicol. 2015, 55, 489–511. [Google Scholar] [CrossRef]
- Ding, J.; Liu, Y.; Lai, Y. Knowledge From London and Berlin: Finding Threads to a Functional HIV Cure. Front. Immunol. 2021, 12, 8747. [Google Scholar] [CrossRef] [PubMed]
- Pauza, C.D.; Huang, K.; Bordon, J. Advances in cell and gene therapy for HIV disease: It is good to be specific. Curr. Opin. HIV AIDS 2021, 16, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Michels, A.; Ho, N.; Buchholz, C.J. Precision medicine: In vivo CAR therapy as a showcase for receptor-targeted vector platforms. Mol. Ther. 2022, 30, 2401–2415. [Google Scholar] [CrossRef] [PubMed]
- Palella, F.J., Jr.; Delaney, K.M.; Moorman, A.C.; Loveless, M.O.; Fuhrer, J.; Satten, G.A.; Aschman, D.J.; Holmberg, S.D.; the HIV Outpatient Study Investigators. Declining Morbidity and Mortality among Patients with Advanced Human Immunodeficiency Virus Infection. N. Engl. J. Med. 1998, 338, 853–860. [Google Scholar] [CrossRef] [PubMed]
- Ta, T.M.; Malik, S.; Anderson, E.M.; Jones, A.D.; Perchik, J.; Freylikh, M.; Sardo, L.; Klase, Z.A.; Izumi, T. Insights Into Persistent HIV-1 Infection and Functional Cure: Novel Capabilities and Strategies. Front. Microbiol. 2022, 13, 2270. [Google Scholar] [CrossRef] [PubMed]
- Darcis, G.; Berkhout, B.; Pasternak, A.O. The Quest for Cellular Markers of HIV Reservoirs: Any Color You Like. Front. Immunol. 2019, 10, 2251. [Google Scholar] [CrossRef] [PubMed]
- Umotoy, J.C.; de Taeye, S.W. Antibody Conjugates for Targeted Therapy Against HIV-1 as an Emerging Tool for HIV-1 Cure. Front. Immunol. 2021, 12, 8806. [Google Scholar] [CrossRef]
- Deeks, S.G. Shock and kill. Nature 2012, 487, 439–440. [Google Scholar] [CrossRef]
- Campbell, G.R.; Spector, S.A. Current strategies to induce selective killing of HIV-1-infected cells. J. Leukoc. Biol. 2022, 112, 1273–1284. [Google Scholar] [CrossRef]
- Crosby, B.; Deas, C.M. Repurposing medications for use in treating HIV infection: A focus on valproic acid as a latency-reversing agent. J. Clin. Pharm. Ther. 2018, 43, 740–745. [Google Scholar] [CrossRef]
- Custodio, M.M.; Sparks, J.; Long, T.E. Disulfiram: A repurposed drug in preclinical and clinical development for the treatment of infectious diseases. Anti-Infect. Agents 2022, 20, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Zerbato, J.M.; Purves, H.V.; Lewin, S.R.; Rasmussen, T.A. Between a shock and a hard place: Challenges and developments in HIV latency reversal. Curr. Opin. Virol. 2019, 38, 1–9. [Google Scholar] [CrossRef] [PubMed]
- van Heuvel, Y.; Schatz, S.; Rosengarten, J.F.; Stitz, J. Infectious RNA: Human Immunodeficiency Virus (HIV) Biology, Therapeutic Intervention, and the Quest for a Vaccine. Toxins 2022, 14, 138. [Google Scholar] [CrossRef] [PubMed]
- Sneller, M.C.; Blazkova, J.; Justement, J.S.; Shi, V.; Kennedy, B.D.; Gittens, K.; Tolstenko, J.; McCormack, G.; Whitehead, E.J.; Schneck, R.F.; et al. Combination anti-HIV antibodies provide sustained virological suppression. Nature 2022, 606, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Leggat, D.J.; Cohen, K.W.; Willis, J.R.; Fulp, W.J.; Decamp, A.C.; Kalyuzhniy, O.; Cottrell, C.A.; Menis, S.; Finak, G.; Ballweber-Fleming, L.; et al. Vaccination induces HIV broadly neutralizing antibody precursors in humans. Science 2022, 378, eadd6502. [Google Scholar] [CrossRef] [PubMed]
- Haynes, B.F.; Kelsoe, G.; Harrison, S.C.; Kepler, T. B-cell–lineage immunogen design in vaccine development with HIV-1 as a case study. Nat. Biotechnol. 2012, 30, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Julg, B. Therapeutic Vaccines for the Treatment of HIV. Transl. Res. 2020, 223, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Packard, T.A.; Schwarzer, R.; Herzig, E.; Rao, D.; Luo, X.; Egedal, J.H.; Hsiao, F.; Widera, M.; Hultquist, J.F.; Grimmett, Z.W.; et al. CCL2: A Chemokine Potentially Promoting Early Seeding of the Latent HIV Reservoir. mBio 2022, 13, e01891-22. [Google Scholar] [CrossRef] [PubMed]
- Al-Ani, A.; Toms, D.; Kondro, D.; Thundathil, J.; Yu, Y.; Ungrin, M. Oxygenation in cell culture: Critical parameters for reproducibility are routinely not reported. PLoS ONE 2018, 13, e0204269. [Google Scholar] [CrossRef]
- Klatt, N.R.; Silvestri, G.; Hirsch, V. Nonpathogenic Simian Immunodeficiency Virus Infections. Cold Spring Harb. Perspect. Med. 2011, 2, a007153. [Google Scholar] [CrossRef]
- Williams, K.C.; Burdo, T.H. HIV and SIV infection: The role of cellular restriction and immune responses in viral replication and pathogenesis. Apmis 2009, 117, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Van Rompay, K.K.A. Tackling HIV and AIDS: Contributions by non-human primate models. Lab Anim. 2017, 46, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Witvrouw, M.; Pannecouque, C.; Switzer, W.M.; Folks, T.M.; De Clercq, E.; Heneine, W. Susceptibility of HIV-2, Siv and Shiv to Various Anti-HIV-1 Compounds: Implications for Treatment and Postexposure Prophylaxis. Antivir. Ther. 2004, 9, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Traggiai, E.; Chicha, L.; Mazzucchelli, L.; Bronz, L.; Piffaretti, J.-C.; Lanzavecchia, A.; Manz, M.G. Development of a Human Adaptive Immune System in Cord Blood Cell-Transplanted Mice. Science 2004, 304, 104–107. [Google Scholar] [CrossRef]
- Baenziger, S.; Tussiwand, R.; Schlaepfer, E.; Mazzucchelli, L.; Heikenwalder, M.; Kurrer, M.O.; Behnke, S.; Frey, J.; Oxenius, A.; Joller, H.; et al. Disseminated and sustained HIV infection in CD34+ cord blood cell-transplanted Rag2-/-gamma c-/- mice. Proc. Natl. Acad. Sci. USA 2006, 103, 15951–15956. [Google Scholar] [CrossRef]
- Nischang, M.; Sutmuller, R.; Gers-Huber, G.; Audigé, A.; Li, D.; Rochat, M.-A.; Baenziger, S.; Hofer, U.; Schlaepfer, E.; Regenass, S.; et al. Humanized Mice Recapitulate Key Features of HIV-1 Infection: A Novel Concept Using Long-Acting Anti-Retroviral Drugs for Treating HIV-1. PLoS ONE 2012, 7, e38853. [Google Scholar] [CrossRef]
- Berges, B.K.; Wheat, W.H.; Palmer, B.E.; Connick, E.; Akkina, R. HIV-1 infection and CD4 T cell depletion in the humanized Rag2-/-gamma c-/- (RAG-hu) mouse model. Retrovirology 2006, 3, 76. [Google Scholar] [CrossRef]
- Watanabe, S.; Ohta, S.; Yajima, M.; Terashima, K.; Ito, M.; Mugishima, H.; Fujiwara, S.; Shimizu, K.; Honda, M.; Shimizu, N.; et al. Humanized NOD/SCID/IL2Rgamma(null) mice transplanted with hematopoietic stem cells under nonmyeloablative conditions show prolonged life spans and allow detailed analysis of human immunodeficiency virus type 1 pathogenesis. J. Virol. 2007, 81, 13259–13264. (In English) [Google Scholar] [CrossRef]
- Watanabe, S.; Terashima, K.; Ohta, S.; Horibata, S.; Yajima, M.; Shiozawa, Y.; Dewan, M.Z.; Yu, Z.; Ito, M.; Morio, T.; et al. Hematopoietic stem cell-engrafted NOD/SCID/IL2Rgamma null mice develop human lymphoid systems and induce long-lasting HIV-1 infection with specific humoral immune responses. Blood 2007, 109, 212–218. [Google Scholar] [CrossRef]
- Ince, W.L.; Zhang, L.; Jiang, Q.; Arrildt, K.; Su, L.; Swanstrom, R. Evolution of the HIV-1 env gene in the Rag2-/- gammaC-/- humanized mouse model. J. Virol. 2010, 84, 2740–2752. (In English) [Google Scholar] [CrossRef]
- Hernandez, M.M.; Fahrny, A.; Jayaprakash, A.; Gers-Huber, G.; Dillon-White, M.; Audigé, A.; Mulder, L.C.F.; Sachidanandam, R.; Speck, R.F.; Simon, V. Impact of Suboptimal APOBEC3G Neutralization on the Emergence of HIV Drug Resistance in Humanized Mice. J. Virol. 2020, 94, e01543-19. [Google Scholar] [CrossRef] [PubMed]
- Klein, F.; Halper-Stromberg, A.; Horwitz, J.A.; Gruell, H.; Scheid, J.F.; Bournazos, S.; Mouquet, H.; Spatz, L.A.; Diskin, R.; Abadir, A.; et al. HIV therapy by a combination of broadly neutralizing antibodies in humanized mice. Nature 2012, 492, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Marsden, M.D.; Kovochich, M.; Suree, N.; Shimizu, S.; Mehta, R.; Cortado, R.; Bristol, G.; An, D.S.; Zack, J.A. HIV Latency in the Humanized BLT Mouse. J. Virol. 2012, 86, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Hess, N.J.; Brown, M.E.; Capitini, C.M. GVHD Pathogenesis, Prevention and Treatment: Lessons From Humanized Mouse Transplant Models. Front. Immunol. 2021, 12, 3544. [Google Scholar] [CrossRef] [PubMed]
- Greenblatt, M.B.; Vbranac, V.; Tivey, T.; Tsang, K.; Tager, A.M.; Aliprantis, A.O. Graft versus Host Disease in the Bone Marrow, Liver and Thymus Humanized Mouse Model. PLoS ONE 2012, 7, e44664. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Cao, F.L.; Zheng, S.G. Update of humanized animal disease models in studying Graft-versus-host disease. Hum. Vaccines Immunother. 2018, 14, 1–6. [Google Scholar] [CrossRef]
- McCune, J.M.; Namikawa, R.; Kaneshima, H.; Shultz, L.D.; Lieberman, M.; Weissman, I.L. The SCID-hu mouse: Murine model for the analysis of human hematolymphoid differentiation and function. Science 1988, 241, 1632–1639. [Google Scholar] [CrossRef]
- Tary-Lehmann, M.; Lehmann, P.V.; Schols, D.; Roncarolo, M.G.; Saxon, A. Anti-SCID mouse reactivity shapes the human CD4+ T cell repertoire in hu-PBL-SCID chimeras. J. Exp. Med. 1994, 180, 1817–1827. [Google Scholar] [CrossRef]
- Ye, C.; Wang, W.; Cheng, L.; Li, G.; Wen, M.; Wang, Q.; Zhang, Q.; Li, D.; Zhou, P.; Su, L. Glycosylphosphatidylinositol-Anchored Anti-HIV scFv Efficiently Protects CD4 T Cells from HIV-1 Infection and Deletion in hu-PBL Mice. J. Virol. 2017, 91, e01389-16. [Google Scholar] [CrossRef]
- Ibarra, G.S.R.; Paul, B.; Sather, B.D.; Younan, P.M.; Sommer, K.; Kowalski, J.P.; Hale, M.; Stoddard, B.; Jarjour, J.; Astrakhan, A.; et al. Efficient Modification of the CCR5 Locus in Primary Human T Cells With megaTAL Nuclease Establishes HIV-1 Resistance. Mol. Ther. Nucleic Acids 2016, 5, e352. [Google Scholar] [CrossRef]
- Asao, H.; Kondo, M.; Tanaka, N.; Ishii, N.; Ohbo, K.; Nakamura, M.; Takeshita, T. The Interleukin-2 Receptor γ Chain: Its Role in the Multiple Cytokine Receptor Complexes and T Cell Development in XSCID. Annu. Rev. Immunol. 1996, 14, 179–205. [Google Scholar] [CrossRef]
- Ma, A.; Koka, R.; Burkett, P. Diverse functions of IL-2, IL-15, and IL-7 in lymphoid homeostasis. Annu. Rev. Immunol. 2006, 24, 657–679. [Google Scholar] [CrossRef]
- Rongvaux, A.; Takizawa, H.; Strowig, T.; Willinger, T.; Eynon, E.E.; Flavell, R.A.; Manz, M.G. Human Hemato-Lymphoid System Mice: Current Use and Future Potential for Medicine. Annu. Rev. Immunol. 2013, 31, 635–674. [Google Scholar] [CrossRef] [PubMed]
- Baenziger, S.; Ziegler, P.; Mazzucchelli, L.; Bronz, L.; Speck, R.F.; Manz, M.G. Human T Cell Development and HIV Infection in Human Hemato-Lymphoid System Mice. Poxviruses 2008, 324, 125–131. [Google Scholar] [CrossRef]
- Gillgrass, A.; Wessels, J.M.; Yang, J.X.; Kaushic, C. Advances in Humanized Mouse Models to Improve Understanding of HIV-1 Pathogenesis and Immune Responses. Front. Immunol. 2021, 11, 7516. [Google Scholar] [CrossRef]
- Lepard, M.; Yang, J.X.; Afkhami, S.; Nazli, A.; Zganiacz, A.; Tang, S.; Choi, M.W.Y.; Vahedi, F.; Deshiere, A.; Tremblay, M.J.; et al. Comparing Current and Next-Generation Humanized Mouse Models for Advancing HIV and HIV/Mtb Co-Infection Studies. Viruses 2022, 14, 1927. [Google Scholar] [CrossRef]
- Lan, P.; Tonomura, N.; Shimizu, A.; Wang, S.; Yang, Y.-G. Reconstitution of a functional human immune system in immunodeficient mice through combined human fetal thymus/liver and CD34+ cell transplantation. Blood 2006, 108, 487–492. [Google Scholar] [CrossRef]
- Namikawa, R.; Kaneshima, H.; Lieberman, M.; Weissman, I.L.; McCune, J.M. Infection of the SCID-hu mouse by HIV-1. Science 1988, 242, 1684–1686. [Google Scholar] [CrossRef]
- Brainard, D.M.; Seung, E.; Frahm, N.; Cariappa, A.; Bailey, C.C.; Hart, W.K.; Shin, H.-S.; Brooks, S.F.; Knight, H.L.; Eichbaum, Q.; et al. Induction of Robust Cellular and Humoral Virus-Specific Adaptive Immune Responses in Human Immunodeficiency Virus-Infected Humanized BLT Mice. J. Virol. 2009, 83, 7305–7321. [Google Scholar] [CrossRef]
- Kawasaki, H.; Yamada, T.; Wada, T.; Kosugi, S. Current status and legal/ethical problems in the research use of the tissues of aborted human fetuses in Japan. Congenit. Anom. 2020, 60, 166–174. [Google Scholar] [CrossRef]
- Lavender, K.J.; Pang, W.W.; Messer, R.J.; Duley, A.K.; Race, B.; Phillips, K.; Scott, D.P.; Peterson, K.E.; Chan, C.K.; Dittmer, U.; et al. BLT-humanized C57BL/6 Rag2−/−γc−/−CD47−/− mice are resistant to GVHD and develop B- and T-cell immunity to HIV infection. Blood 2013, 122, 4013–4020. [Google Scholar] [CrossRef] [PubMed]
- Stripecke, R.; Münz, C.; Schuringa, J.J.; Bissig, K.; Soper, B.; Meeham, T.; Yao, L.; Di Santo, J.P.; Brehm, M.; Rodriguez, E.; et al. Innovations, challenges, and minimal information for standardization of humanized mice. EMBO Mol. Med. 2020, 12, e8662. [Google Scholar] [CrossRef] [PubMed]
- Volk, V.; Reppas, A.I.; Robert, P.A.; Spineli, L.M.; Sundarasetty, B.S.; Theobald, S.J.; Schneider, A.; Gerasch, L.; Roth, C.D.; Klöss, S.; et al. Multidimensional Analysis Integrating Human T-Cell Signatures in Lymphatic Tissues with Sex of Humanized Mice for Prediction of Responses after Dendritic Cell Immunization. Front. Immunol. 2017, 8, 1709. [Google Scholar] [CrossRef]
- Notta, F.; Doulatov, S.; Laurenti, E.; Poeppl, A.; Jurisica, I.; Dick, J.E. Isolation of Single Human Hematopoietic Stem Cells Capable of Long-Term Multilineage Engraftment. Science 2011, 333, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Christopher, A.C.; Venkatesan, V.; Karuppusamy, K.V.; Srinivasan, S.; Babu, P.; Azhagiri, M.K.K.; Chambayil, K.; Bagchi, A.; Rajendiran, V.; Ravi, N.S.; et al. Preferential Expansion of Human CD34+CD133+CD90+ Hematopoietic Stem Cells Enhances Gene-Modified Cell Frequency for Gene Therapy. Hum. Gene Ther. 2022, 33, 188–201. [Google Scholar] [CrossRef]
- Brehm, M.A.; Kenney, L.L.; Wiles, M.V.; Low, B.E.; Tisch, R.M.; Burzenski, L.; Mueller, C.; Greiner, D.L.; Shultz, L.D. Lack of acute xenogeneic graft- versus -host disease, but retention of T-cell function following engraftment of human peripheral blood mononuclear cells in NSG mice deficient in MHC class I and II expression. FASEB J. 2018, 33, 3137–3151. [Google Scholar] [CrossRef]
- Yaguchi, T.; Kobayashi, A.; Inozume, T.; Morii, K.; Nagumo, H.; Nishio, H.; Iwata, T.; Ka, Y.; Katano, I.; Ito, R.; et al. Human PBMC-transferred murine MHC class I/II-deficient NOG mice enable long-term evaluation of human immune responses. Cell. Mol. Immunol. 2017, 15, 953–962. [Google Scholar] [CrossRef]
- Cheng, L.; Ma, J.; Li, G.; Su, L. Humanized Mice Engrafted With Human HSC Only or HSC and Thymus Support Comparable HIV-1 Replication, Immunopathology, and Responses to ART and Immune Therapy. Front. Immunol. 2018, 9, 817. [Google Scholar] [CrossRef]
- Hur, E.M.; Patel, S.N.; Shimizu, S.; Rao, D.S.; Gnanapragasam, P.N.P.; An, N.S.; Yang, L.; Baltimore, D. Inhibitory effect of HIV-specific neutralizing IgA on mucosal transmission of HIV in humanized mice. Blood 2012, 120, 4571–4582. [Google Scholar] [CrossRef]
- Holguin, L.; Echavarria, L.; Burnett, J.C. Novel Humanized Peripheral Blood Mononuclear Cell Mouse Model with Delayed Onset of Graft-versus-Host Disease for Preclinical HIV Research. J. Virol. 2022, 96, e01394-21. [Google Scholar] [CrossRef]
- Ivic, S.; Rochat, M.-A.; Li, D.; Audigé, A.; Schlaepfer, E.; Münz, C.; Manz, M.G.; Speck, R.F. Differential Dynamics of HIV Infection in Humanized MISTRG versus MITRG Mice. Immunohorizons 2017, 1, 162–175. [Google Scholar] [CrossRef]
- Rongvaux, A.; Willinger, T.; Martinek, J.; Strowig, T.; Gearty, S.; Teichmann, L.L.; Saito, Y.; Marches, F.; Halene, S.; Palucka, A.K.; et al. Development and function of human innate immune cells in a humanized mouse model. Nat. Biotechnol. 2014, 32, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Oronsky, B.; Carter, C.; Reid, T.; Brinkhaus, F.; Knox, S.J. Just eat it: A review of CD47 and SIRP-α antagonism. Semin. Oncol. 2020, 47, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Kang, G.; Prathipati, P.K.; Fan, W.; Li, Q.; Destache, C.J. Long-acting parenteral combination antiretroviral loaded nano-drug delivery system to treat chronic HIV-1 infection: A humanized mouse model study. Antivir. Res. 2018, 156, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Kessing, C.F.; Nixon, C.C.; Li, C.; Tsai, P.; Takata, H.; Mousseau, G.; Ho, P.T.; Honeycutt, J.B.; Fallahi, M.; Trautmann, L.; et al. In Vivo Suppression of HIV Rebound by Didehydro-Cortistatin A, a “Block-and-Lock” Strategy for HIV-1 Treatment. Cell Rep. 2017, 21, 600–611. [Google Scholar] [CrossRef]
- Weichseldorfer, M.; Affram, Y.; Heredia, A.; Rikhtegaran-Tehrani, Z.; Sajadi, M.M.; Williams, S.P.; Tagaya, Y.; Benedetti, F.; Ramadhani, H.O.; Denaro, F.; et al. Combined cART including Tenofovir Disoproxil, Emtricitabine, and Dolutegravir has potent therapeutic effects in HIV-1 infected humanized mice. J. Transl. Med. 2021, 19, 1–10. [Google Scholar] [CrossRef]
- Satheesan, S.; Li, H.; Burnett, J.C.; Takahashi, M.; Li, S.; Wu, S.X.; Synold, T.; Rossi, J.J.; Zhou, J. HIV Replication and Latency in a Humanized NSG Mouse Model during Suppressive Oral Combinational Antiretroviral Therapy. J. Virol. 2018, 92, e02118-17. [Google Scholar] [CrossRef]
- Li, Q.; Tso, F.Y.; Kang, G.; Lu, W.; Li, Y.; Fan, W.; Yuan, Z.; Destache, C.; Wood, C. Early Initiation of Antiretroviral Therapy Can Functionally Control Productive HIV-1 Infection in Humanized-BLT Mice. Am. J. Ther. 2015, 69, 519–527. [Google Scholar] [CrossRef]
- Kumar, P.; Ban, H.-S.; Kim, S.-S.; Wu, H.; Pearson, T.; Greiner, D.L.; Laouar, A.; Yao, J.; Haridas, V.; Habiro, K.; et al. T Cell-Specific siRNA Delivery Suppresses HIV-1 Infection in Humanized Mice. Cell 2008, 134, 577–586. [Google Scholar] [CrossRef]
- Beloor, J.; Kudalkar, S.N.; Buzzelli, G.; Yang, F.; Mandl, H.K.; Rajashekar, J.K.; Spasov, K.A.; Jorgensen, W.L.; Saltzman, W.M.; Anderson, K.S.; et al. Long-acting and extended-release implant and nanoformulations with a synergistic antiretroviral two-drug combination controls HIV-1 infection in a humanized mouse model. Bioeng. Transl. Med. 2021, 7, e10237. [Google Scholar] [CrossRef]
- Hoffmann-Fezer, G.; Gall, C.; Zengerle, U.; Kranz, B.; Thierfelder, S. Immunohistology and immunocytology of human T-cell chimerism and graft-versus-host disease in SCID mice. Blood 1993, 81, 3440–3448. [Google Scholar] [CrossRef] [PubMed]
- Burgunder, E.; Fallon, J.K.; White, N.; Schauer, A.P.; Sykes, C.; Remling-Mulder, L.; Kovarova, M.; Adamson, L.; Luciw, P.; Garcia, J.V.; et al. Antiretroviral Drug Concentrations in Lymph Nodes: A Cross-Species Comparison of the Effect of Drug Transporter Expression, Viral Infection, and Sex in Humanized Mice, Nonhuman Primates, and Humans. Experiment 2019, 370, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Devanathan, A.S.; Pirone, J.R.; Akkina, R.; Remling-Mulder, L.; Luciw, P.; Adamson, L.; Garcia, J.V.; Kovarova, M.; White, N.R.; Schauer, A.P.; et al. Antiretroviral Penetration across Three Preclinical Animal Models and Humans in Eight Putative HIV Viral Reservoirs. Antimicrob. Agents Chemother. 2019, 64, e01639-19. [Google Scholar] [CrossRef] [PubMed]
- Grüll, H.; Klein, F. Progress in HIV-1 antibody research using humanized mice. Curr. Opin. HIV AIDS 2017, 12, 285–293. [Google Scholar] [CrossRef]
- Horwitz, J.A.; Halper-Stromberg, A.; Mouquet, H.; Gitlin, A.D.; Tretiakova, A.; Eisenreich, T.R.; Malbec, M.; Gravemann, S.; Billerbeck, E.; Dorner, M.; et al. HIV-1 suppression and durable control by combining single broadly neutralizing antibodies and antiretroviral drugs in humanized mice. Proc. Natl. Acad. Sci. USA 2013, 110, 16538–16543. [Google Scholar] [CrossRef] [PubMed]
- Schommers, P.; Gruell, H.; Abernathy, M.E.; Tran, M.-K.; Dingens, A.S.; Gristick, H.B.; Barnes, C.O.; Schoofs, T.; Schlotz, M.; Vanshylla, K.; et al. Restriction of HIV-1 Escape by a Highly Broad and Potent Neutralizing Antibody. Cell 2020, 180, 471–489.e22. [Google Scholar] [CrossRef] [PubMed]
- Freund, N.T.; Horwitz, J.A.; Nogueira, L.; Sievers, S.A.; Scharf, L.; Scheid, J.F.; Gazumyan, A.; Liu, C.; Velinzon, K.; Goldenthal, A.; et al. A New Glycan-Dependent CD4-Binding Site Neutralizing Antibody Exerts Pressure on HIV-1 In Vivo. PLoS Pathog. 2015, 11, e1005238. [Google Scholar] [CrossRef]
- Bournazos, S.; Gazumyan, A.; Seaman, M.S.; Nussenzweig, M.C.; Ravetch, J.V. Bispecific Anti-HIV-1 Antibodies with Enhanced Breadth and Potency. Cell 2016, 165, 1609–1620. [Google Scholar] [CrossRef]
- Klein, F.; Nogueira, L.; Nishimura, Y.; Phad, G.E.; West, A.P.; Halper-Stromberg, A.; Horwitz, J.A.; Gazumyan, A.; Liu, C.; Eisenreich, T.R.; et al. Enhanced HIV-1 immunotherapy by commonly arising antibodies that target virus escape variants. J. Exp. Med. 2014, 211, 2361–2372. [Google Scholar] [CrossRef]
- Halper-Stromberg, A.; Lu, C.-L.; Klein, F.; Horwitz, J.A.; Bournazos, S.; Nogueira, L.; Eisenreich, T.R.; Liu, C.; Gazumyan, A.; Schaefer, U.; et al. Broadly Neutralizing Antibodies and Viral Inducers Decrease Rebound from HIV-1 Latent Reservoirs in Humanized Mice. Cell 2014, 158, 989–999. [Google Scholar] [CrossRef]
- Haneberg, B.; Kendall, D.; Apter, F.M.; Neutra, M.R. Distribution of Monoclonal Antibodies in Intestinal and Urogenital Secretions of Mice Bearing Hybridoma ‘Backpack’ Tumours. Scand. J. Immunol. 1997, 45, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.M.; Lei, M.Y.Y.; Feidi, R.A.; West, A.P.; Balazs, A.B.; Bjorkman, P.J.; Yang, L.; Baltimore, D. Dimeric 2G12 as a Potent Protection against HIV-1. PLoS Pathog. 2010, 6, e1001225. [Google Scholar] [CrossRef] [PubMed]
- Balazs, A.B.; Chen, J.; Hong, C.M.; Rao, D.S.; Yang, L.; Baltimore, D. Antibody-based protection against HIV infection by vectored immunoprophylaxis. Nature 2011, 481, 81–84. [Google Scholar] [CrossRef] [PubMed]
- van der Velden, Y.U.; Villaudy, J.; Siteur-van Rijnstra, E.; van der Linden, C.A.; Frankin, E.; Weijer, K.; Schermer, E.; Vink, M.A.; Berkhout, B.; Sanders, R.W.; et al. Short Communication: Protective Efficacy of Broadly Neutralizing Antibody PGDM1400 Against HIV-1 Challenge in Humanized Mice. AIDS Res. Hum. Retrovir. 2018, 34, 790–793. [Google Scholar] [CrossRef] [PubMed]
- Stoddart, C.A.; Galkina, S.A.; Joshi, P.; Kosikova, G.; Long, B.R.; Maidji, E.; Moreno, M.E.; Rivera, J.M.; Sanford, U.R.; Sloan, B.; et al. Efficacy of broadly neutralizing monoclonal antibody PG16 in HIV-infected humanized mice. Virology 2014, 462–463, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Herndler-Brandstetter, D.; Shan, L.; Yao, Y.; Stecher, C.; Plajer, V.; Lietzenmayer, M.; Strowig, T.; de Zoete, M.R.; Palm, N.W.; Chen, J.; et al. Humanized mouse model supports development, function, and tissue residency of human natural killer cells. Proc. Natl. Acad. Sci. USA 2017, 114, E9626–E9634. [Google Scholar] [CrossRef]
- Rajashekar, J.K.; Richard, J.; Beloor, J.; Prévost, J.; Anand, S.P.; Beaudoin-Bussières, G.; Shan, L.; Herndler-Brandstetter, D.; Gendron-Lepage, G.; Medjahed, H.; et al. Modulating HIV-1 envelope glycoprotein conformation to decrease the HIV-1 reservoir. Cell Host Microbe 2021, 29, 904–916.e6. [Google Scholar] [CrossRef]
- Holt, N.; Wang, J.; Kim, K.; Friedman, G.; Wang, X.; Taupin, V.; Crooks, G.M.; Kohn, D.B.; Gregory, P.D.; Holmes, M.C.; et al. Human hematopoietic stem/progenitor cells modified by zinc-finger nucleases targeted to CCR5 control HIV-1 in vivo. Nat. Biotechnol. 2010, 28, 839–847. [Google Scholar] [CrossRef]
- Myburgh, R.; Ivic, S.; Pepper, M.S.; Gers-Huber, G.; Li, D.; Audigé, A.; Rochat, M.-A.; Jaquet, V.; Regenass, S.; Manz, M.; et al. Lentivector Knockdown of CCR5 in Hematopoietic Stem and Progenitor Cells Confers Functional and Persistent HIV-1 Resistance in Humanized Mice. J. Virol. 2015, 89, 6761–6772. [Google Scholar] [CrossRef]
- Falkenhagen, A.; Singh, J.; Asad, S.; Leontyev, D.; Read, S.; Zúñiga-Pflücker, J.C.; Joshi, S. Control of HIV Infection In Vivo Using Gene Therapy with a Secreted Entry Inhibitor. Mol. Ther. Nucleic Acids 2017, 9, 132–144. [Google Scholar] [CrossRef]
- Khamaikawin, W.; Shimizu, S.; Kamata, M.; Cortado, R.; Jung, Y.; Lam, J.; Wen, J.; Kim, P.; Xie, Y.; Kim, S.; et al. Modeling Anti-HIV-1 HSPC-Based Gene Therapy in Humanized Mice Previously Infected with HIV-1. Mol. Ther. Methods Clin. Dev. 2017, 9, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Petit, N.Y.; Baillou, C.; Burlion, A.; Dorgham, K.; Levacher, B.; Amiel, C.; Schneider, V.; Lemoine, F.M.; Gorochov, G.; Marodon, G. Gene transfer of two entry inhibitors protects CD4+ T cell from HIV-1 infection in humanized mice. Gene Ther. 2015, 23, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yang, H.; Gao, Y.; Chen, Z.; Xie, L.; Liu, Y.; Liu, Y.; Wang, X.; Li, H.; Lai, W.; et al. CRISPR/Cas9-Mediated CCR5 Ablation in Human Hematopoietic Stem/Progenitor Cells Confers HIV-1 Resistance In Vivo. Mol. Ther. 2017, 25, 1782–1789. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M.W.; Guo, L.; Xin, F.; Yang, X.; Riley, J.L. Stabilized Human TRIM5α Protects Human T Cells From HIV-1 Infection. Mol. Ther. 2014, 22, 1084–1095. [Google Scholar] [CrossRef] [PubMed]
- Didigu, C.A.; Wilen, C.B.; Wang, J.; Duong, J.; Secreto, A.J.; Danet-Desnoyers, G.A.; Riley, J.L.; Gregory, P.D.; June, C.H.; Holmes, M.C.; et al. Simultaneous zinc-finger nuclease editing of the HIV coreceptors ccr5 and cxcr4 protects CD4+ T cells from HIV-1 infection. Blood 2014, 123, 61–69. [Google Scholar] [CrossRef]
- Maldini, C.R.; Claiborne, D.T.; Okawa, K.; Chen, T.; Dopkin, D.L.; Shan, X.; Power, K.A.; Trifonova, R.T.; Krupp, K.; Phelps, M.; et al. Dual CD4-based CAR T cells with distinct costimulatory domains mitigate HIV pathogenesis in vivo. Nat. Med. 2020, 26, 1776–1787. [Google Scholar] [CrossRef]
- Kitchen, S.G.; Levin, B.R.; Bristol, G.; Rezek, V.; Kim, S.; Aguilera-Sandoval, C.; Balamurugan, A.; Yang, O.O.; Zack, J.A. In Vivo Suppression of HIV by Antigen Specific T Cells Derived from Engineered Hematopoietic Stem Cells. PLoS Pathog. 2012, 8, e1002649. [Google Scholar] [CrossRef]
- Ringpis, G.-E.E.; Shimizu, S.; Arokium, H.; Camba-Colón, J.; Carroll, M.V.; Cortado, R.; Xie, Y.; Kim, P.Y.; Sahakyan, A.; Lowe, E.L.; et al. Engineering HIV-1-Resistant T-Cells from Short-Hairpin RNA-Expressing Hematopoietic Stem/Progenitor Cells in Humanized BLT Mice. PLoS ONE 2012, 7, e53492. [Google Scholar] [CrossRef]
- Shimizu, S.; Ringpis, G.-E.; Marsden, M.D.; Cortado, R.V.; Wilhalme, H.M.; Elashoff, D.; Zack, J.A.; Chen, I.S.Y.; An, D.S. RNAi-Mediated CCR5 Knockdown Provides HIV-1 Resistance to Memory T Cells in Humanized BLT Mice. Mol. Ther. Nucleic Acids 2015, 4, e227. [Google Scholar] [CrossRef]
- Hauber, I.; Hofmann-Sieber, H.; Chemnitz, J.; Dubrau, D.; Chusainow, J.; Stucka, R.; Hartjen, P.; Schambach, A.; Ziegler, P.; Hackmann, K.; et al. Highly Significant Antiviral Activity of HIV-1 LTR-Specific Tre-Recombinase in Humanized Mice. PLoS Pathog. 2013, 9, e1003587. [Google Scholar] [CrossRef]
- Richter, M.; Saydaminova, K.; Yumul, R.; Krishnan, R.; Liu, J.; Nagy, E.-E.; Singh, M.; Izsvak, Z.; Cattaneo, R.; Uckert, W.; et al. In vivo transduction of primitive mobilized hematopoietic stem cells after intravenous injection of integrating adenovirus vectors. Blood 2016, 128, 2206–2217. [Google Scholar] [CrossRef] [PubMed]
- Brendel, C.; Rio, P.; Verhoeyen, E. Humanized mice are precious tools for evaluation of hematopoietic gene therapies and preclinical modeling to move towards a clinical trial. Biochem. Pharmacol. 2019, 174, 113711. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Liu, W.; Bauch, T.; Graviss, E.A.; Arduino, R.C.; Kimata, J.T.; Chen, M.; Wang, J. Clearance of HIV infection by selective elimination of host cells capable of producing HIV. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Nixon, C.C.; Mavigner, M.; Sampey, G.C.; Brooks, A.D.; Spagnuolo, R.A.; Irlbeck, D.M.; Mattingly, C.; Ho, P.T.; Schoof, N.; Cammon, C.G.; et al. Systemic HIV and SIV latency reversal via non-canonical NF-κB signalling in vivo. Nature 2020, 578, 160–165. [Google Scholar] [CrossRef] [PubMed]
- McBrien, J.B.; Mavigner, M.; Franchitti, L.; Smith, S.A.; White, E.; Tharp, G.K.; Walum, H.; Busman-Sahay, K.; Aguilera-Sandoval, C.R.; Thayer, W.O.; et al. Robust and persistent reactivation of SIV and HIV by N-803 and depletion of CD8+ cells. Nature 2020, 578, 154–159. [Google Scholar] [CrossRef]
- Tsai, P.; Wu, G.; Baker, C.E.; Thayer, W.O.; Spagnuolo, R.A.; Sanchez, R.; Barrett, S.; Howell, B.; Margolis, D.; Hazuda, D.J.; et al. In vivo analysis of the effect of panobinostat on cell-associated HIV RNA and DNA levels and latent HIV infection. Retrovirology 2016, 13, 1–12. [Google Scholar] [CrossRef]
- Marsden, M.D.; Zhang, T.-H.; Du, Y.; Dimapasoc, M.; Soliman, M.S.; Wu, X.; Kim, J.T.; Shimizu, A.; Schrier, A.; Wender, P.A.; et al. Tracking HIV Rebound following Latency Reversal Using Barcoded HIV. Cell Rep. Med. 2020, 1, 100162. [Google Scholar] [CrossRef]
- Pache, L.; Marsden, M.D.; Teriete, P.; Portillo, A.J.; Heimann, D.; Kim, J.T.; Soliman, M.S.; Dimapasoc, M.; Carmona, C.; Celeridad, M.; et al. Pharmacological Activation of Non-canonical NF-κB Signaling Activates Latent HIV-1 Reservoirs In Vivo. Cell Rep. Med. 2020, 1, 100037. [Google Scholar] [CrossRef]
- Lavender, K.J.; Pace, C.; Sutter, K.; Messer, R.J.; Pouncey, D.; Cummins, N.W.; Natesampillai, S.; Zheng, J.; Goldsmith, J.; Widera, M.; et al. An advanced BLT-humanized mouse model for extended HIV-1 cure studies. Aids 2018, 32, 1–10. [Google Scholar] [CrossRef]
- Llewellyn, G.N.; Seclén, E.; Wietgrefe, S.; Liu, S.; Chateau, M.; Pei, H.; Perkey, K.; Marsden, M.D.; Hinkley, S.J.; Paschon, D.E.; et al. Humanized Mouse Model of HIV-1 Latency with Enrichment of Latent Virus in PD-1+ and TIGIT+ CD4 T Cells. J. Virol. 2019, 93, e02086-18. [Google Scholar] [CrossRef]
- Gao, H.; Ozantürk, A.N.; Wang, Q.; Harlan, G.H.; Schmitz, A.J.; Presti, R.M.; Deng, K.; Shan, L. Evaluation of HIV-1 Latency Reversal and Antibody-Dependent Viral Clearance by Quantification of Singly Spliced HIV-1 vpu/env mRNA. J. Virol. 2021, 95, e02124-20. [Google Scholar] [CrossRef] [PubMed]
- Sungur, C.M.; Wang, Q.; Ozantürk, A.N.; Gao, H.; Schmitz, A.J.; Cella, M.; Yokoyama, W.M.; Shan, L. Human NK cells confer protection against HIV-1 infection in humanized mice. J. Clin. Investig. 2022, 132. [Google Scholar] [CrossRef] [PubMed]
- Flerin, N.C.; Bardhi, A.; Zheng, J.H.; Korom, M.; Folkvord, J.; Kovacs, C.; Benko, E.; Truong, R.; Mota, T.; Connick, E.; et al. Establishment of a Novel Humanized Mouse Model To Investigate In Vivo Activation and Depletion of Patient-Derived HIV Latent Reservoirs. J. Virol. 2019, 93, e02051-18. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Nie, C.; Misawa, N.; Tanaka, Y.; Ito, M.; Koyanagi, Y. Dynamics of memory and naive CD8+ T lymphocytes in humanized NOD/SCID/IL-2Rgammanull mice infected with CCR5-tropic HIV-1. Vaccine 2010, 28 (Suppl. S2), B32–B37. (In English) [Google Scholar] [CrossRef] [PubMed]
- Gorantla, S.; Makarov, E.; Finke-Dwyer, J.; Gebhart, C.L.; Domm, W.; Dewhurst, S.; Gendelman, H.E.; Poluektova, L.Y. CD8+ Cell Depletion Accelerates HIV-1 Immunopathology in Humanized Mice. J. Immunol. 2010, 184, 7082–7091. [Google Scholar] [CrossRef]
- Majji, S.; Wijayalath, W.; Shashikumar, S.; Pow-Sang, L.; Villasante, E.; Brumeanu, T.D.; Casares, S. Differential effect of HLA class-I versus class-II transgenes on human T and B cell reconstitution and function in NRG mice. Sci. Rep. 2016, 6, 28093. [Google Scholar] [CrossRef]
- Ollerton, M.T.; Folkvord, J.M.; Peachman, K.K.; Shashikumar, S.; Morrison, E.B.; Jagodzinski, L.L.; Peel, S.A.; Khreiss, M.; D’aquila, R.T.; Casares, S.; et al. HIV-1 infected humanized DRAGA mice develop HIV-specific antibodies despite lack of canonical germinal centers in secondary lymphoid tissues. Front. Immunol. 2022, 13, 1047277. [Google Scholar] [CrossRef]
- Godot, V.; Tcherakian, C.; Gil, L.; Cervera-Marzal, I.; Li, G.; Cheng, L.; Ortonne, N.; Lelièvre, J.-D.; Pantaleo, G.; Fenwick, C.; et al. TLR-9 agonist and CD40-targeting vaccination induces HIV-1 envelope-specific B cells with a diversified immunoglobulin repertoire in humanized mice. PLoS Pathog. 2020, 16, e1009025. [Google Scholar] [CrossRef]
- Li, Y.; Masse-Ranson, G.; Garcia, Z.; Bruel, T.; Kök, A.; Strick-Marchand, H.; Jouvion, G.; Serafini, N.; Lim, A.I.; Dusseaux, M.; et al. A human immune system mouse model with robust lymph node development. Nat. Methods 2018, 15, 623–630. [Google Scholar] [CrossRef]
- Takahashi, T.; Katano, I.; Ito, R.; Goto, M.; Abe, H.; Mizuno, S.; Kawai, K.; Sugiyama, F.; Ito, M. Enhanced Antibody Responses in a Novel NOG Transgenic Mouse with Restored Lymph Node Organogenesis. Front. Immunol. 2018, 8, 2017. [Google Scholar] [CrossRef]
- Dudek, T.E.; No, D.C.; Seung, E.; Vrbanac, V.D.; Fadda, L.; Bhoumik, P.; Boutwell, C.L.; Power, K.A.; Gladden, A.D.; Battis, L.; et al. Rapid Evolution of HIV-1 to Functional CD8 + T Cell Responses in Humanized BLT Mice. Sci. Transl. Med. 2012, 4, 143ra98. [Google Scholar] [CrossRef] [PubMed]
- Seung, E.; Dudek, T.E.; Allen, T.; Freeman, G.J.; Luster, A.D.; Tager, A.M. PD-1 Blockade in Chronically HIV-1-Infected Humanized Mice Suppresses Viral Loads. PLoS ONE 2013, 8, e77780. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Wang, Q.; Li, G.; Banga, R.; Ma, J.; Yu, H.; Yasui, F.; Zhang, Z.; Pantaleo, G.; Perreau, M.; et al. TLR3 agonist and CD40-targeting vaccination induces immune responses and reduces HIV-1 reservoirs. J. Clin. Investig. 2018, 128, 4387–4396. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Chang, H.; Sarkis, P.T.N.; Fikrig, E.; Zhu, Q.; Marasco, W.A. Humoral immune responses in humanized BLT mice immunized with West Nile virus and HIV-1 envelope proteins are largely mediated via human CD5+ B cells. Immunology 2011, 134, 419–433. [Google Scholar] [CrossRef]
- Garcia-Beltran, W.F.; Claiborne, D.T.; Maldini, C.R.; Phelps, M.; Vrbanac, V.; Karpel, M.E.; Krupp, K.L.; Power, K.A.; Boutwell, C.L.; Balazs, A.B.; et al. Innate Immune Reconstitution in Humanized Bone Marrow-Liver-Thymus (HuBLT) Mice Governs Adaptive Cellular Immune Function and Responses to HIV-1 Infection. Front. Immunol. 2021, 12, 667393. [Google Scholar] [CrossRef]
- Wang, H.; Madariaga, M.L.; Wang, S.; Van Rooijen, N.; Oldenborg, P.-A.; Yang, Y.-G. Lack of CD47 on nonhematopoietic cells induces split macrophage tolerance to CD47 null cells. Proc. Natl. Acad. Sci. USA 2007, 104, 13744–13749. [Google Scholar] [CrossRef]
- Rutishauser, R.L.; Trautmann, L. CD8+ T-cell responses in HIV controllers: Potential implications for novel HIV remission strategies. Curr. Opin. HIV AIDS 2022, 17, 315–324. [Google Scholar] [CrossRef]
- Rosengarten, J.F.; Schatz, S.; Wolf, T.; Barbe, S.; Stitz, J. Components of a HIV-1 vaccine mediate virus-like particle (VLP)-formation and display of envelope proteins exposing broadly neutralizing epitopes. Virology 2022, 568, 41–48. [Google Scholar] [CrossRef]
- Bailon, L.; Alarcón-Soto, Y.; Benet, S. Challenges of HIV therapeutic vaccines clinical trials design. Curr. Opin. HIV AIDS 2022, 17, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Macatangay, B.J.C.; Landay, A.L.; Garcia, F.; Rinaldo, C.R. Editorial: Advances in T Cell Therapeutic Vaccines for HIV. Front. Immunol. 2022, 13, 905836. [Google Scholar] [CrossRef]
- Deruaz, M.; Luster, A.D. BLT Humanized Mice as Model to Study HIV Vaginal Transmission. J. Infect. Dis. 2013, 208, S131–S136. [Google Scholar] [CrossRef] [PubMed]
- Denton, P.; Estes, J.D.; Sun, Z.; Othieno, F.A.; Wei, B.L.; Wege, A.K.; Powell, D.; Payne, D.; Haase, A.T.; Garcia, J.V. Antiretroviral Pre-exposure Prophylaxis Prevents Vaginal Transmission of HIV-1 in Humanized BLT Mice. PLoS Med. 2008, 5, e16. [Google Scholar] [CrossRef] [PubMed]
- Denton, P.W.; Othieno, F.; Martinez-Torres, F.; Zou, W.; Krisko, J.F.; Fleming, E.; Zein, S.; Powell, D.A.; Wahl, A.; Kwak, Y.T.; et al. One Percent Tenofovir Applied Topically to Humanized BLT Mice and Used According to the CAPRISA 004 Experimental Design Demonstrates Partial Protection from Vaginal HIV Infection, Validating the BLT Model for Evaluation of New Microbicide Candidates. J. Virol. 2011, 85, 7582–7593. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, L.A.; Trifonova, R.; Vrbanac, V.; Basar, E.; McKernan, S.; Xu, Z.; Seung, E.; Deruaz, M.; Dudek, T.; Einarsson, J.I.; et al. Inhibition of HIV transmission in human cervicovaginal explants and humanized mice using CD4 aptamer-siRNA chimeras. J. Clin. Investig. 2011, 121, 2401–2412. [Google Scholar] [CrossRef] [PubMed]
- Berges, B.K.; Akkina, S.R.; Folkvord, J.M.; Connick, E.; Akkina, R. Mucosal transmission of R5 and X4 tropic HIV-1 via vaginal and rectal routes in humanized Rag2-/- gammac-/- (RAG-hu) mice. Virology 2008, 373, 342–351. (In English) [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Prathipati, P.K.; Kang, G.; Zhou, Y.; Yuan, Z.; Fan, W.; Li, Q.; Destache, C.J. Tenofovir alafenamide and elvitegravir loaded nanoparticles for long-acting prevention of HIV-1 vaginal transmission. Aids 2017, 31, 469–476. [Google Scholar] [CrossRef]
- Deruaz, M.; Moldt, B.; Le, K.M.; Power, K.A.; Vrbanac, V.D.; Tanno, S.; Ghebremichael, M.S.; Allen, T.M.; Tager, A.M.; Burton, D.R.; et al. Protection of Humanized Mice From Repeated Intravaginal HIV Challenge by Passive Immunization: A Model for Studying the Efficacy of Neutralizing Antibodies In Vivo. J. Infect. Dis. 2016, 214, 612–616. [Google Scholar] [CrossRef]
- Brady, J.M.; Phelps, M.; MacDonald, S.W.; Lam, E.C.; Nitido, A.; Parsons, D.; Boutros, C.L.; Deal, C.E.; Garcia-Beltran, W.F.; Tanno, S.; et al. Antibody-mediated prevention of vaginal HIV transmission is dictated by IgG subclass in humanized mice. Sci. Transl. Med. 2022, 14. [Google Scholar] [CrossRef]
- Olesen, R.; Swanson, M.D.; Kovarova, M.; Nochi, T.; Chateau, M.; Honeycutt, J.; Long, J.M.; Denton, P.; Hudgens, M.G.; Richardson, A.; et al. ART influences HIV persistence in the female reproductive tract and cervicovaginal secretions. J. Clin. Investig. 2016, 126, 892–904. [Google Scholar] [CrossRef]
- Hofer, U.; Baenziger, S.; Heikenwalder, M.; Schlaepfer, E.; Gehre, N.; Regenass, S.; Brunner, T.; Speck, R.F. RAG2-/- gamma(c)-/- mice transplanted with CD34+ cells from human cord blood show low levels of intestinal engraftment and are resistant to rectal transmission of human immunodeficiency virus. J. Virol. 2008, 82, 12145–12153. (In English) [Google Scholar] [CrossRef]
- Nguyen, P.V.; Wessels, J.M.; Mueller, K.; Vahedi, F.; Anipindi, V.; Verschoor, C.P.; Chew, M.; Deshiere, A.; Karniychuk, U.; Mazzulli, T.; et al. Frequency of Human CD45+ Target Cells is a Key Determinant of Intravaginal HIV-1 Infection in Humanized Mice. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef]
- Maidji, C.A.S.E.; Galkina, S.A.; Kosikova, G.; Rivera, J.M.; Moreno, M.E.; Sloan, B.; Joshi, P.; Longa, B.R. Superior human leukocyte reconstitution and susceptibility to vaginal HIV transmission in humanized NOD-scid IL-2Rgamma(-/-) (NSG) BLT mice. Virology 2011, 417, 154–160. (In English) [Google Scholar] [CrossRef]
- Veselinovic, M.; Neff, C.P.; Mulder, L.R.; Akkina, R. Topical gel formulation of broadly neutralizing anti-HIV-1 monoclonal antibody VRC01 confers protection against HIV-1 vaginal challenge in a humanized mouse model. Virology 2012, 432, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Wessels, J.M.; Nguyen, P.V.; Vitali, D.; Mueller, K.; Vahedi, F.; Felker, A.M.; Dupont, H.A.; Bagri, P.; Verschoor, C.P.; Deshiere, A.; et al. Depot medroxyprogesterone acetate (DMPA) enhances susceptibility and increases the window of vulnerability to HIV-1 in humanized mice. Sci. Rep. 2021, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Veselinovic, M.; Charlins, P.; Akkina, R. Modeling HIV-1 Mucosal Transmission and Prevention in Humanized Mice. Methods Mol. Biol. 2016, 1354, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Denton, P.; Krisko, J.F.; Powell, D.; Mathias, M.; Kwak, Y.T.; Martinez-Torres, F.; Zou, W.; Payne, D.A.; Estes, J.D.; Garcia, J.V. Systemic Administration of Antiretrovirals Prior to Exposure Prevents Rectal and Intravenous HIV-1 Transmission in Humanized BLT Mice. PLoS ONE 2010, 5, e8829. [Google Scholar] [CrossRef]
- Hioe, C.E.; Li, G.; Liu, X.; Tsahouridis, O.; He, X.; Funaki, M.; Klingler, J.; Tang, A.F.; Feyznezhad, R.; Heindel, D.W.; et al. Non-neutralizing antibodies targeting the immunogenic regions of HIV-1 envelope reduce mucosal infection and virus burden in humanized mice. PLoS Pathog. 2022, 18, e1010183. [Google Scholar] [CrossRef]
- Vanshylla, K.; Held, K.; Eser, T.M.; Gruell, H.; Kleipass, F.; Stumpf, R.; Jain, K.; Weiland, D.; Münch, J.; Grüttner, B.; et al. CD34T+ Humanized Mouse Model to Study Mucosal HIV-1 Transmission and Prevention. Vaccines 2021, 9, 198. [Google Scholar] [CrossRef]
- Boily, M.-C.; Baggaley, R.F.; Wang, L.; Masse, B.; White, R.G.; Hayes, R.J.; Alary, M. Heterosexual risk of HIV-1 infection per sexual act: Systematic review and meta-analysis of observational studies. Lancet Infect. Dis. 2009, 9, 118–129. [Google Scholar] [CrossRef]
- Andrews, S.M.; Rowland-Jones, S. Recent advances in understanding HIV evolution. F1000Research 2017, 6, 597. [Google Scholar] [CrossRef]
- Cheng, L.; Yu, H.; Wrobel, J.A.; Li, G.; Liu, P.; Hu, Z.; Xu, X.-N.; Su, L. Identification of pathogenic TRAIL-expressing innate immune cells during HIV-1 infection in humanized mice by scRNA-seq. J. Clin. Investig. 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Adoro, S.; Cubillos-Ruiz, J.R.; Chen, X.; Deruaz, M.; Vrbanac, V.D.; Song, M.; Park, S.; Murooka, T.T.; Dudek, T.E.; Luster, A.D.; et al. IL-21 induces antiviral microRNA-29 in CD4 T cells to limit HIV-1 infection. Nat. Commun. 2015, 6, 7562. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Wang, Q.; Ghneim, K.; Wang, L.; Rampanelli, E.; Holley-Guthrie, E.; Cheng, L.; Garrido, C.; Margolis, D.M.; Eller, L.A.; et al. Multi-omics analyses reveal that HIV-1 alters CD4+ T cell immunometabolism to fuel virus replication. Nat. Immunol. 2021, 22, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Aryee, K.; Burzenski, L.M.; Yao, L.; Keck, J.G.; Greiner, D.L.; Shultz, L.D.; Brehm, M.A. Enhanced development of functional human NK cells in NOD- scid-IL2rgnull mice expressing human IL15. FASEB J. 2022, 36, e22476. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; Ono, R.; Iyoda, T.; Endo, T.; Iwasaki, M.; Tomizawa-Murasawa, M.; Saito, Y.; Kaneko, A.; Shimizu, K.; Yamada, D.; et al. Human NK cell development in hIL-7 and hIL-15 knockin NOD/SCID/IL2rgKO mice. Life Sci. Alliance 2019, 2, e201800195. [Google Scholar] [CrossRef]
- Zhang, J.; Lohani, S.C.; Cheng, Y.; Wang, T.; Guo, L.; Kim, W.-K.; Gorantla, S.; Li, Q. Human Microglia Extensively Reconstitute in Humanized-BLT Mice With Human Interleukin-34 Transgene and Support HIV-1 Brain Infection. Front. Immunol. 2021, 12, 672415. [Google Scholar] [CrossRef]
- Honeycutt, J.B.; Wahl, A.; Archin, N.; Choudhary, S.; Margolis, D.; Garcia, J.V. HIV-1 infection, response to treatment and establishment of viral latency in a novel humanized T cell-only mouse (TOM) model. Retrovirology 2013, 10, 121. [Google Scholar] [CrossRef]
- Honeycutt, J.B.; Wahl, A.; Baker, C.; Spagnuolo, R.A.; Foster, J.; Zakharova, O.; Wietgrefe, S.; Caro-Vegas, C.; Madden, V.; Sharpe, G.; et al. Macrophages sustain HIV replication in vivo independently of T cells. J. Clin. Investig. 2016, 126, 1353–1366. [Google Scholar] [CrossRef]
- Honeycutt, J.B.; Thayer, W.O.; Baker, C.E.; Ribeiro, R.M.; Lada, S.M.; Cao, Y.; Cleary, R.A.; Hudgens, M.G.; Richman, D.D.; Garcia, J.V. HIV persistence in tissue macrophages of humanized myeloid-only mice during antiretroviral therapy. Nat. Med. 2017, 23, 638–643. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baroncini, L.; Bredl, S.; Nicole, K.P.; Speck, R.F. The Humanized Mouse Model: What Added Value Does It Offer for HIV Research? Pathogens 2023, 12, 608. https://doi.org/10.3390/pathogens12040608
Baroncini L, Bredl S, Nicole KP, Speck RF. The Humanized Mouse Model: What Added Value Does It Offer for HIV Research? Pathogens. 2023; 12(4):608. https://doi.org/10.3390/pathogens12040608
Chicago/Turabian StyleBaroncini, Luca, Simon Bredl, Kadzioch P. Nicole, and Roberto F. Speck. 2023. "The Humanized Mouse Model: What Added Value Does It Offer for HIV Research?" Pathogens 12, no. 4: 608. https://doi.org/10.3390/pathogens12040608
APA StyleBaroncini, L., Bredl, S., Nicole, K. P., & Speck, R. F. (2023). The Humanized Mouse Model: What Added Value Does It Offer for HIV Research? Pathogens, 12(4), 608. https://doi.org/10.3390/pathogens12040608