
Disease Progression and Serological Assay Performance in Heritage Breed Pigs following Brucella suis Experimental Challenge as a Model for Naturally Infected Feral Swine

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Bacteria
2.3. Experimental Design
2.4. Clinical Observations
2.5. Culture and Serology
2.6. Statistical Analyses
2.6.1. Differences in Tissues Infected
2.6.2. Diagnostic Assay Performance
2.6.3. Changes in Detection Probability through Time
2.6.4. Optimal Combination of Diagnostic Assays
3. Results
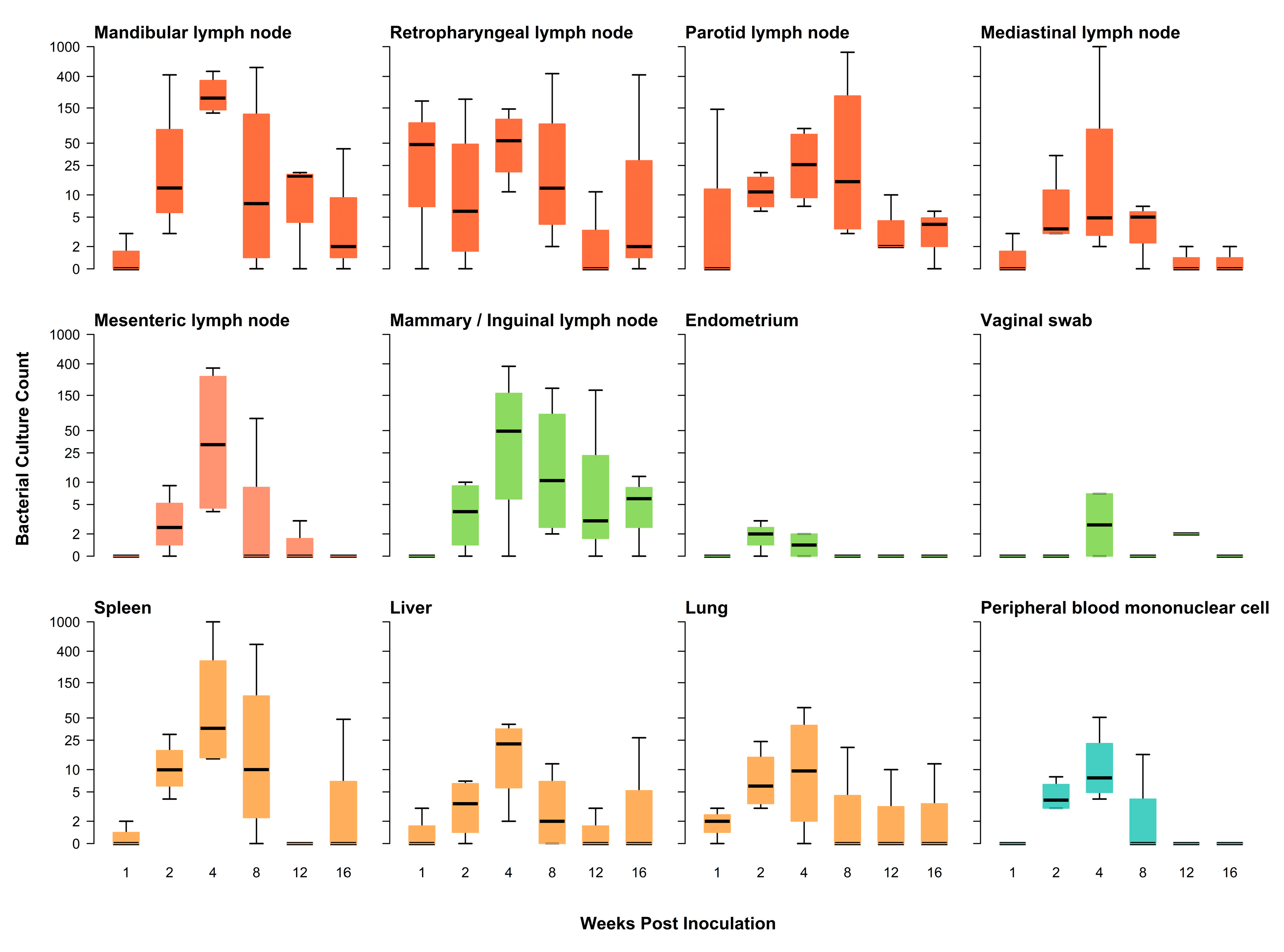
3.1. Differences in Tissues Infected and Serological Results
3.1.1. Clinical Signs and Direct Culture
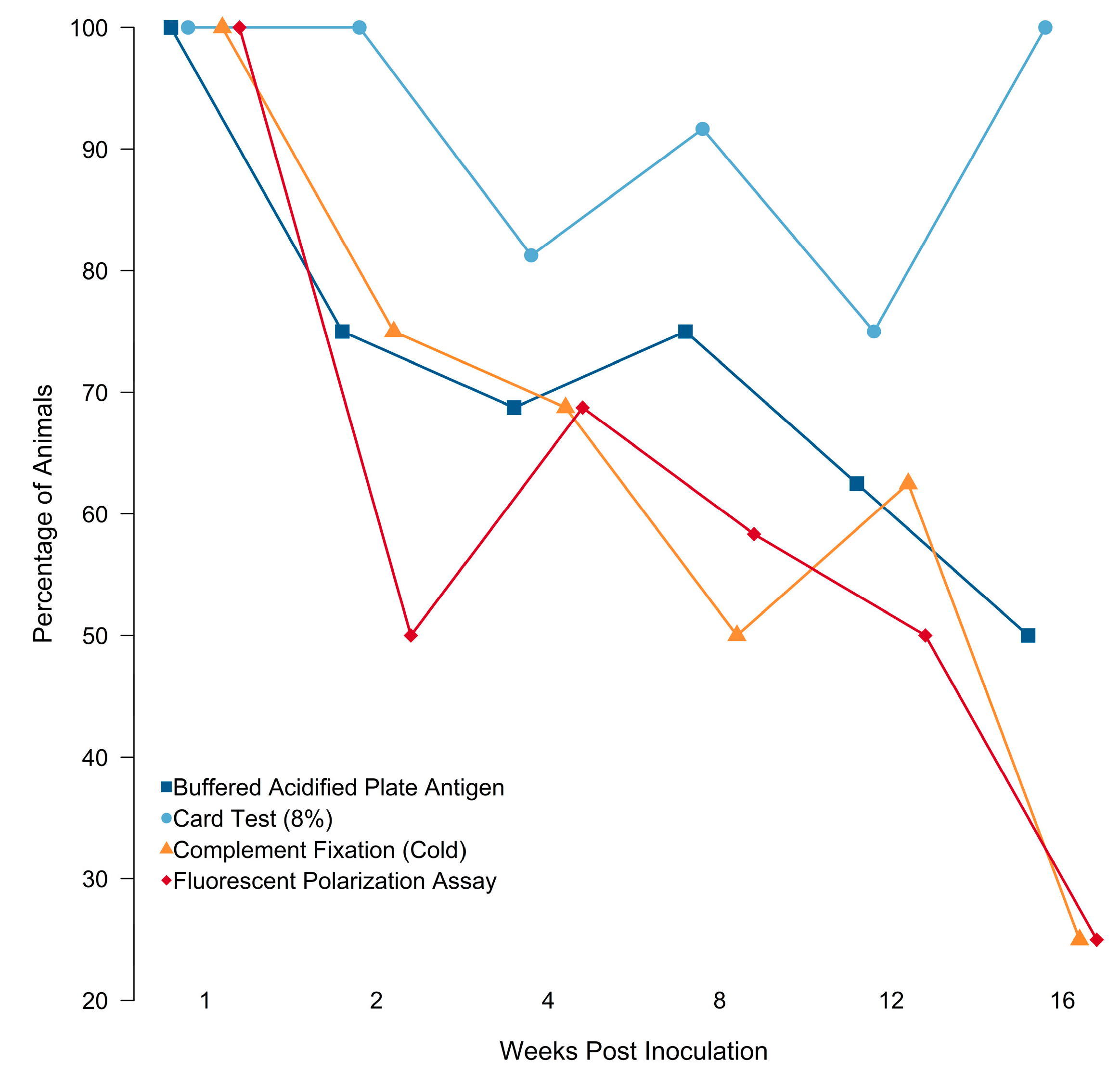
3.1.2. Serology
3.2. Diagnostic Assay Performance
3.2.1. Single Assay Performance
3.2.2. Changes in Detection Probability through Time
3.2.3. Optimal Combination of Diagnostic Assays
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, R.S.; Bevins, S.N.; Cook, G.; Free, R.; Pepin, K.M.; Gidlewski, T.; Brown, V.R. Adaptive risk-based targeted surveillance for foreign animal diseases at the wildlife-livestock interface. Transbound Emerg. Dis. 2022, 69, e2329–e2340. [Google Scholar] [CrossRef]
- Miller, R.S.; Farnsworth, M.L.; Malmberg, J.L. Diseases at the livestock–wildlife interface: Status, challenges, and opportunities in the United States. Prev. Vet. Med. 2013, 110, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Vercauteren, K.C.; Miller, R.S. Characteristics and perspectives of disease at the wildlife-livestock interface in North America. In Diseases at the Wildlife-Livestock Interface; Vicente, J., Gortàzar, C., Eds.; Springer: Cham, Switzerland, 2021; Volume 3, pp. 245–269. [Google Scholar]
- Ryser-Degiorgis, M.-P. Wildlife health investigations: Needs, challenges and recommendations. BMC Vet. Res. 2013, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Stallknecht, D. Impediments to wildlife disease surveillance, research, and diagnostics. In Wildlife and Emerging Zoonotic Diseases: The Biology, Circumstances and Consequences of Cross-Species Transmission, 1st ed.; Childs, J.E., Mackenzie, J.S., Eds.; Springer: Heidelberg, Germany, 2007; pp. 445–461. [Google Scholar] [CrossRef]
- Brown, V.R.; Marlow, M.C.; Gidlewski, T.; Bowen, R.; Bosco-Lauth, A. Perspectives on the past, present, and future of feral swine disease surveillance in the United States. J. Anim. Sci. 2020, 98, skaa256. [Google Scholar] [CrossRef]
- Miller, R.S.; Sweeney, S.J.; Slootmaker, C.; Grear, D.A.; Di Salvo, P.A.; Kiser, D.; Shwiff, S.A. Cross-species transmission potential between wild pigs, livestock, poultry, wildlife, and humans: Implications for disease risk management in North America. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bevins, S.N.; Pedersen, K.; Lutman, M.W.; Gidlewski, T.; Deliberto, T.J. Consequences associated with the recent range expansion of nonnative feral swine. Bioscience 2014, 64, 291–299. [Google Scholar] [CrossRef]
- Brown, V.R.; Bowen, R.A.; Bosco-Lauth, A.M. Zoonotic pathogens from feral swine that pose a significant threat to public health. Transbound Emerg. Dis. 2018, 65, 649–659. [Google Scholar] [CrossRef]
- Hutton, T.; DeLiberto, T.J.; Owen, S.; Morrison, B. Disease risks associated with increasing feral swine numbers and distribution in the United States. Mich. Bov. Tuberc. Bibliogr. Database 2006, 59, 1–15. [Google Scholar]
- Olsen, S.; Tatum, F. Swine brucellosis: Current perspectives. Vet. Med. Res. Rep. 2017, 8, 1. [Google Scholar] [CrossRef]
- Alton, G.G. Brucella suis. In Animal Brucellosis; Neilsen, K., Duncan, J.R., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 411–422. [Google Scholar]
- Alton, G.G.; Jones, L.M.; Angus, R.; Verger, J. Techniques for the brucellosis laboratory. Br. Vet. J. 1990, 142, 118. [Google Scholar]
- Cvetnic, Z.; Spicic, S.; Curic, S.; Jukic, B.; Lojkic, M.; Albert, D.; Thiébaud, M.; Garin-Bastuji, B. Isolation of Brucella suis biovar 3 from horses in Croatia. Vet. Rec. 2005, 156, 584–585. [Google Scholar] [CrossRef]
- Ewalt, D.R.; Payeur, J.B.; Rhyan, J.C.; Geer, P.L. Brucella suis biovar 1 in naturally infected cattle: A bacteriological, serological, and histological study. J. Vet. Diagn. Investig. 1997, 9, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, S.; Woldemeskel, M.; Ligett, A.; Snider, R.; Cobb, R.; Rajeev, S. Brucella suis infection in dogs, Georgia, USA. Emerg. Infect. Dis. 2011, 17, 2386–2387. [Google Scholar] [CrossRef]
- Tessaro, S.V. The existing and potential importance of brucellosis and tuberculosis in Canadian wildlife: A review. Can. Vet. J. 1986, 27, 119–122. [Google Scholar] [PubMed]
- Deyoe, B. Histopathologic changes in male swine with experimental brucellosis. Am. J. Vet. Res. 1968, 29, 1215–1220. [Google Scholar] [PubMed]
- Jungersen, G.; SØrensen, V.; Giese, S.B.; Stack, J.A.; Riber, U. Differentiation between serological responses to Brucella suis and Yersinia enterocolitica serotype O:9 after natural or experimental infection in pigs. Epidemiol. Infect. 2006, 134, 347–357. [Google Scholar] [CrossRef]
- 9 CFR-78.43. Validated Brucellosis-Free States. 9 2011. 9 CFR § 78.43-Validated Brucellosis-Free States. |Electronic Code of Federal Regulations (e-CFR)|US Law|LII/Legal Information Institute (cornell.edu). Available online: https://www.law.cornell.edu/cfr/text/9/78.43 (accessed on 1 March 2023).
- Miller, R.S. Interaction among Societal and Biological Drivers of Policy at the Wildlife-Agricultural Interface. Ph.D. Thesis, Colorado State University, Fort Colins, CO, USA, 2017. [Google Scholar]
- Cvetnić, Z.; Spicić, S.; Toncić, J.; Majnarić, D.; Benić, M.; Albert, D.; Thiébaud, M.; Garin-Bastuji, B. Brucella suis infection in domestic pigs and wild boar in Croatia. Rev. Sci. Tech. 2009, 28, 1057–1067. [Google Scholar] [CrossRef]
- Wu, N.; Abril, C.; Thomann, A.; Grosclaude, E.; Doherr, M.G.; Boujon, P.; Ryser-Degiorgis, M.-P. Risk factors for contacts between wild boar and outdoor pigs in Switzerland and investigations on potential Brucella suis spill-over. BMC Vet. Res. 2012, 8, 1–12. [Google Scholar] [CrossRef]
- Cortina, M.E.; Balzano, R.E.; Serantes, D.A.R.; Caillava, A.J.; Elena, S.; Ferreira, A.; Nicola, A.M.; Ugalde, J.E.; Comerci, D.J.; Ciocchini, A.E. A bacterial glycoengineered antigen for improved serodiagnosis of porcine brucellosis. J. Clin. Micro. 2016, 54, 1448–1455. [Google Scholar] [CrossRef]
- Godfroid, J.; Nielsen, K.; Saegerman, C. Diagnosis of brucellosis in livestock and wildlife. Croat. Med. J. 2010, 51, 296–305. [Google Scholar] [CrossRef]
- Praud, A.; Gimenez, O.; Zanella, G.; Dufour, B.; Pozzi, N.; Antras, V.; Meyer, L.; Garin-Bastuji, B. Estimation of sensitivity and specificity of five serological tests for the diagnosis of porcine brucellosis. Prev. Vet. Med. 2012, 104, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.T.; Garcia-Carreras, B.; Hitchings, M.D.T.; Yang, B.; Katzelnick, L.C.; Rattigan, S.M.; Borgert, B.A.; Moreno, C.A.; Solomon, B.D.; Trimmer-Smith, L.; et al. A systematic review of antibody mediated immunity to coronaviruses: Kinetics, correlates of protection, and association with severity. Nat. Commun. 2020, 11, 4704. [Google Scholar] [CrossRef]
- Sánchez-Cordón, P.J.; Nunez, A.; Neimanis, A.; Wikström-Lassa, E.; Montoya, M.; Crooke, H.; Gavier-Widén, D. African swine fever: Disease dynamics in wild boar experimentally infected with ASFV isolates belonging to genotype I and II. Viruses 2019, 11, 852. [Google Scholar] [CrossRef] [PubMed]
- Mayer, J.J.; Brisbin, I.L. Wild Pigs in the United States: Their History, Comparative Morphology, and Current Status; University of Georgia Press: Athens, GA, USA, 2008. [Google Scholar]
- USDA APHIS. Complement Fixation Test for Detection of Antibodies to Brucella abortus and Brucella suis; USDA APHIS: Riverdale Park, MD, USA, 2014; pp. 1–63.
- USDA APHIS. Card Test for Detection of Antibodies to Brucella abortus/suis; USDA APHIS: Riverdale Park, MD, USA, 2015; pp. 1–5.
- USDA APHIS. Buffered Acidified Plate Antigen Test for Detection of Antibodies to Brucella; USDA APHIS: Riverdale Park, MD, USA, 2020; pp. 1–5.
- USDA APHIS. Brucella Fluorescence Polarization Assay (FPA) Using Tubes; USDA APHIS: Riverdale Park, MD, USA, 2020; pp. 1–5.
- Hinić, V.; Brodard, I.; Thomann, A.; Cvetnić, Z.; Makaya, P.V.; Frey, J.; Abril, C. Novel identification and differentiation of Brucella melitensis, B. abortus, B. suis, B. ovis, B. canis, and B. neotomae suitable for both conventional and real-time PCR systems. J. Microbiol. Methods 2008, 75, 375–378. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria. Available online: https://www.r-project.org (accessed on 15 June 2022).
- Rubin, D.B. The bayesian bootstrap. Ann. Stat. 1981, 130–134. [Google Scholar] [CrossRef]
- bayesboot: An Implementation of Rubin’s (1981) Bayesian Bootstrap, Version 0.2; Rasmus Bååth 2018. Available online: https://cran.r-project.org/web/packages/bayesboot/bayesboot.pdf (accessed on 15 June 2022).
- Gelman, A.; Hill, J. Data Analysis Using Regression and Multilevel/Hierarchical Models; Cambridge University Press: Cambrifge, UK, 2007. [Google Scholar]
- Geweke, J.F. Evaluating the Accuracy of Sampling-Based Approaches to the Calculation of Posterior Moments; Staff Report 148; Federal Reserve Bank of Minneapolis: Minneapolis, MN, USA, 1991. [Google Scholar]
- Gelman, A.; Rubin, D.B. Inference from iterative simulation using multiple sequences. Stat. Sci. 1992, 7, 457–472. [Google Scholar] [CrossRef]
- Vehtari, A.; Gelman, A.; Gabry, J. Practical Bayesian model evaluation using leave-one-out cross-validation and WAIC. Stat. Comput. 2017, 27, 1413–1432. [Google Scholar] [CrossRef]
- Gelman, A.; Goodrich, B.; Gabry, J.; Vehtari, A. R-squared for Bayesian regression models. Am. Stat. 2019, 73, 307–309. [Google Scholar] [CrossRef]
- Goodrich, B.; Gabry, J.; Ali, I.; Brilleman, S. rstanarm: Bayesian applied regression modeling via Stan. R package version 2018, 2, 1758. [Google Scholar]
- Hanley, J.A.; McNeil, B.J. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 1982, 143, 29–36. [Google Scholar] [CrossRef]
- Hajian-Tilaki, K. Receiver operating characteristic (ROC) curve analysis for medical diagnostic test evaluation. Caspian. J. Intern. Med. 2013, 4, 627. [Google Scholar]
- Khan, M.R.A.A. ROCit-An R Package for Performance Assessment of Binary Classifier with Visualization; South Dakota State University: Brookings, SD, USA, 2019. [Google Scholar]
- Tabak, M.A.; Pedersen, K.; Miller, R.S. Detection error influences both temporal seroprevalence predictions and risk factors associations in wildlife disease models. Ecol. Evol. 2019, 9, 10404–10414. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, K.; Bauer, N.; Olsen, S.; Arenas-Gamboa, A.; Henry, A.; Sibley, T.; Gidlewski, T. Identification of Brucella spp. in feral swine (Sus scrofa) at abattoirs in Texas, USA. Zoonoses Public Health 2017, 64, 647–654. [Google Scholar] [CrossRef] [PubMed]
- USDA APHIS. What Is Swine Brucellosis? Available online: https://www.aphis.usda.gov/aphis/ourfocus/animalhealth/animal-disease-information/swine-disease-information/swine-brucellosis (accessed on 15 June 2022).
- Van Der Leek, M.; Becker, H.; Humphrey, P.; Adams, C.; Belden, R.; Frankenberger, W.; Nicoletti, P. Prevalence of Brucella sp. antibodies in feral swine in Florida. J. Wildl. Dis. 1993, 29, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Stoffregen, W.C.; Olsen, S.C.; Wheeler, C.J.; Bricker, B.J.; Palmer, M.V.; Jensen, A.E.; Halling, S.M.; Alt, D.P. Diagnostic Characterization of a Feral Swine Herd Enzootically Infected with Brucella. J. Vet. Diagn. Invest. 2007, 19, 227–237. [Google Scholar] [CrossRef]
- Garin-Bastuji, B.; Blasco, J.M. Brucellosis (infection with Brucella abortus, B. melitensis, and B. suis). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2019; OIE: Paris, France, 2016; pp. 355–398. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pig #e | 774 | 776 | 777 | 731 | 771 | 879 | 880 | 768 | 878 | 728 | 737 | 881 | 885 | 735 | 775 | 733 | 770 | 729 | 772 | 730 | 732 | 773 | 736 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Wpi Euth b | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 4 | 4 | 4 | 4 | 8 | 8 | 8 | 8 | 12 | 12 | 12 | 12 | 16 | 16 | 16 | 16 | |
| Sex c | M | M | F | F | F | M | F | F | M | F | M | F | M | F | M | F | M | F | M | M | F | M | F | |
| Age d | A | - | A | A | - | A | A | A | A | A | A | A | A | A | SA | A | A | A | SA | J | - | SA | J | |
| Strain a | 294 | 1330 | 1330 | 1330 | 1330 | 294 | 294 | 294 | 294 | 1330 | 1330 | 294 | 294 | 1330 | 1330 | 294 | 294 | 1330 | 1330 | 294 | 294 | 1330 | 1330 | Percent Animals |
| PBMC | 0 | 0 | 0 | + | + | + | + | + | + | + | + | 0 | 0 | 0 | + | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 39 |
| Vaginal Swab | 0 | 0 | 0 | 0 | + | 0 | 0 | 0 | + | 0 | 0 | 0 | 17 | |||||||||||
| Endometrium | 0 | + | + | 0 | 0 | + | 0 | 0 | 0 | 0 | 0 | 0 | 25 | |||||||||||
| Testis | 0 | 0 | ||||||||||||||||||||||
| Mandibular LN | + f | 0 | 0 | + | ++ | + | + | ++ | ++ | ++ | ++ | 0 | + | + | +++ | + | 0 | 0 | + | + | 0 | + | 0 | 65 |
| Retropharyngeal LN | ++ | 0 | + | + | ++ | 0 | + | ++ | + | + | + | + | + | + | ++ | + | 0 | 0 | 0 | + | 0 | ++ | 0 | 70 |
| Parotid LN | ++ | 0 | 0 | + | + | + | + | + | + | + | + | + | + | + | +++ | + | + | 0 | + | 0 | 0 | + | + | 78 |
| Mediastinal LN | + | 0 | 0 | + | + | + | + | +++ | + | + | + | 0 | + | + | + | 0 | 0 | 0 | + | 0 | 0 | + | 0 | 61 |
| Mesenteric LN | 0 | 0 | 0 | 0 | + | + | + | ++ | ++ | + | + | 0 | 0 | 0 | + | + | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 39 |
| Mammary/Inguinal LN | 0 | 0 | 0 | + | + | + | 0 | ++ | + | 0 | + | + | + | + | ++ | ++ | 0 | 0 | + | 0 | 0 | + | + | 61 |
| Spleen | 0 | + | 0 | + | + | + | + | +++ | + | + | + | + | 0 | + | ++ | 0 | 0 | 0 | 0 | 0 | 0 | + | 0 | 57 |
| Liver | 0 | + | 0 | + | 0 | + | + | + | + | + | + | 0 | 0 | + | + | 0 | 0 | 0 | + | 0 | 0 | + | 0 | 52 |
| Lung | 0 | + | + | + | + | + | + | + | + | 0 | + | 0 | 0 | 0 | + | 0 | 0 | 0 | + | 0 | 0 | + | 0 | 52 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brown, V.R.; Miller, R.S.; Bowden, C.F.; Smyser, T.J.; Ledesma, N.A.; Hartwig, A.; Gordy, P.; Anderson, A.M.; Porter, S.M.; Alexander, K.; et al. Disease Progression and Serological Assay Performance in Heritage Breed Pigs following Brucella suis Experimental Challenge as a Model for Naturally Infected Feral Swine. Pathogens 2023, 12, 638. https://doi.org/10.3390/pathogens12050638
Brown VR, Miller RS, Bowden CF, Smyser TJ, Ledesma NA, Hartwig A, Gordy P, Anderson AM, Porter SM, Alexander K, et al. Disease Progression and Serological Assay Performance in Heritage Breed Pigs following Brucella suis Experimental Challenge as a Model for Naturally Infected Feral Swine. Pathogens. 2023; 12(5):638. https://doi.org/10.3390/pathogens12050638
Chicago/Turabian StyleBrown, Vienna R., Ryan S. Miller, Courtney F. Bowden, Timothy J. Smyser, Nicholas A. Ledesma, Airn Hartwig, Paul Gordy, Aaron M. Anderson, Stephanie M. Porter, Kate Alexander, and et al. 2023. "Disease Progression and Serological Assay Performance in Heritage Breed Pigs following Brucella suis Experimental Challenge as a Model for Naturally Infected Feral Swine" Pathogens 12, no. 5: 638. https://doi.org/10.3390/pathogens12050638
APA StyleBrown, V. R., Miller, R. S., Bowden, C. F., Smyser, T. J., Ledesma, N. A., Hartwig, A., Gordy, P., Anderson, A. M., Porter, S. M., Alexander, K., Gouker, Z., Gidlewski, T., Bowen, R. A., & Bosco-Lauth, A. M. (2023). Disease Progression and Serological Assay Performance in Heritage Breed Pigs following Brucella suis Experimental Challenge as a Model for Naturally Infected Feral Swine. Pathogens, 12(5), 638. https://doi.org/10.3390/pathogens12050638