




HSV-1 Triggers an Antiviral Transcriptional Response during Viral Replication That Is Completely Abrogated in PKR−/− Cells



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Virus and Viral Infections
2.3. Design of PKR siRNAs and Detection of the Transcript Levels in HEp-2 Cells
2.4. Western Blot Analysis
2.5. DNA Extraction
2.6. RNA Extraction, Reverse Transcription, and Real-Time PCR
2.7. Statistical Analysis
3. Results
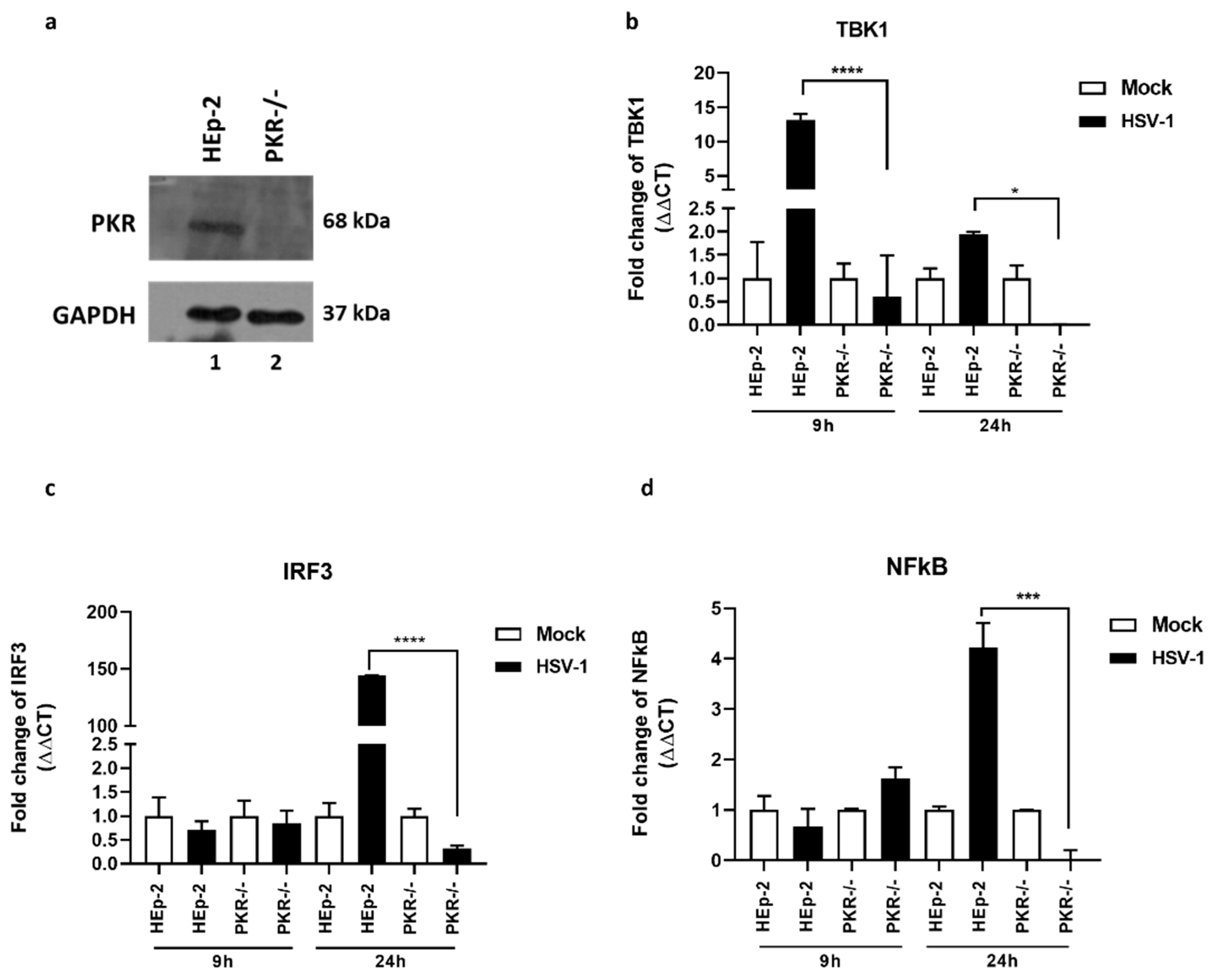
3.1. Impact of PKR Knockout in the TBK1-IRF3-NF-κB Network
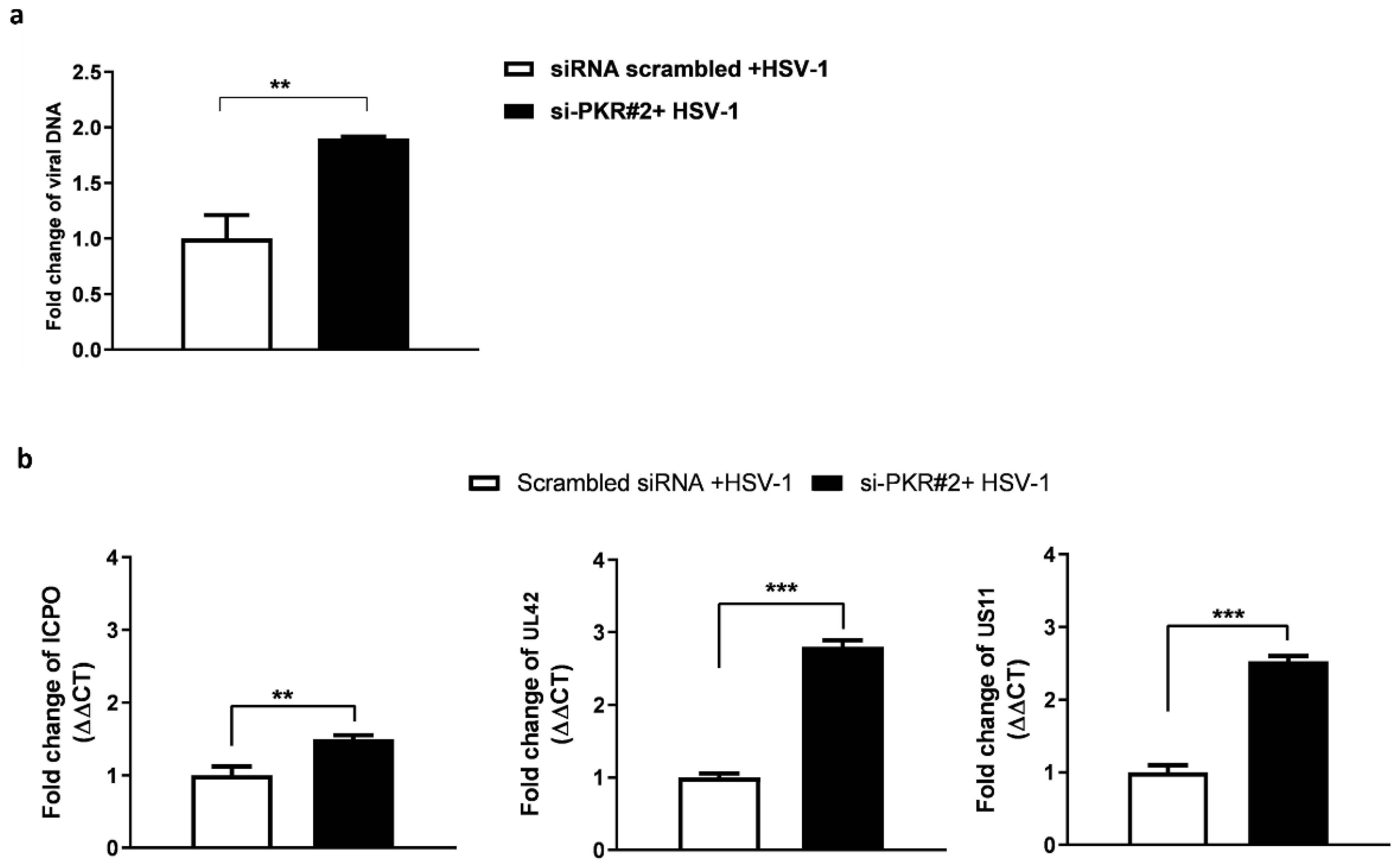
3.2. Depletion of PKR Uniquely Improves Viral DNA Accumulation and HSV-1 Gene Expression
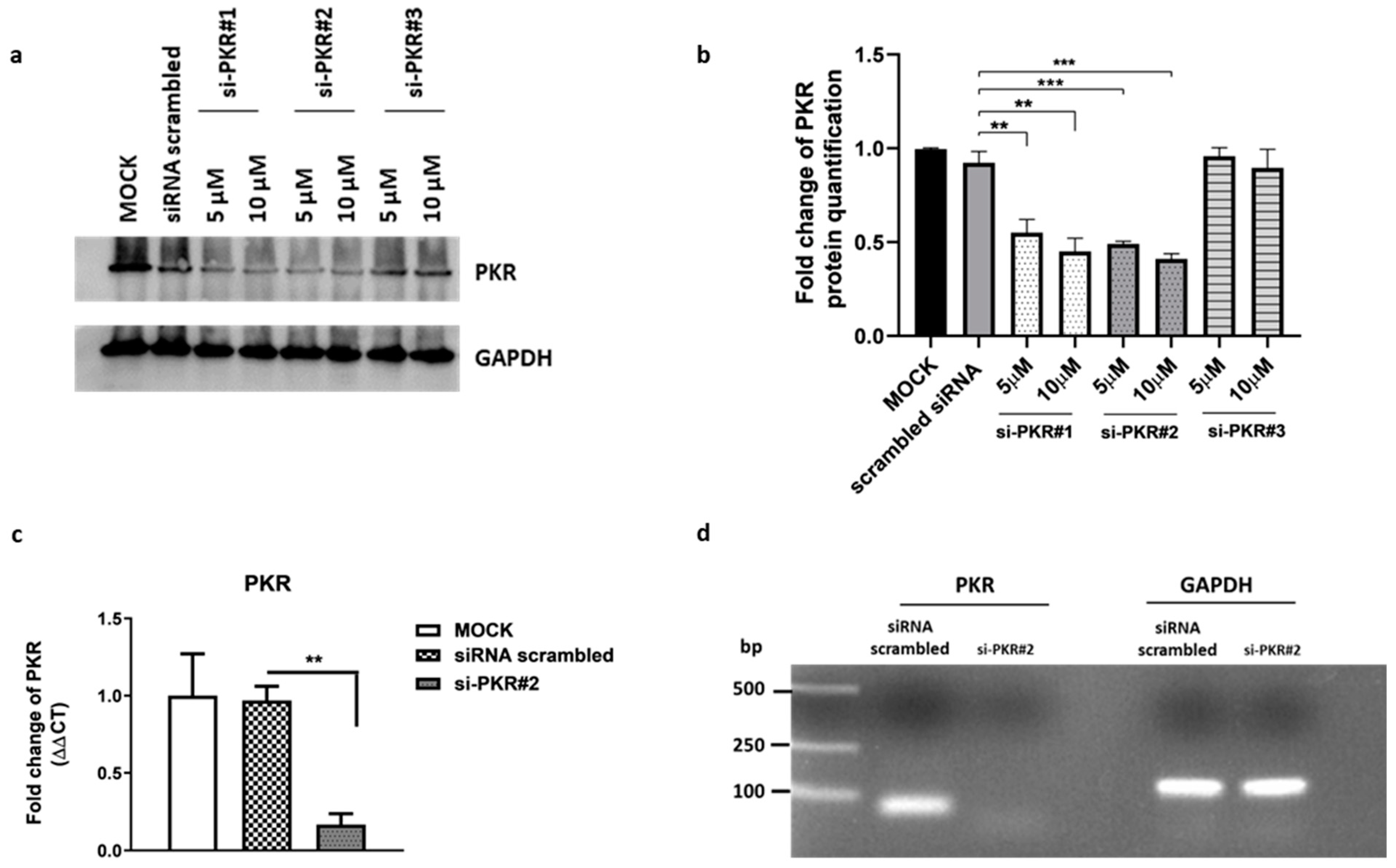
3.3. Effect of PKR-Silencing during HSV-1 Replication
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Honess, R.W.; Roizman, B. Regulation of herpesvirus macromolecular synthesis. I. Cascade regulation of the synthesis of three groups of viral proteins. J. Virol. 1974, 14, 8–19. [Google Scholar] [CrossRef]
- Honess, R.W.; Roizman, B. Regulation of herpesvirus macromolecular synthesis: Sequential transition of polypeptide synthesis requires functional viral polypeptides. Proc. Natl. Acad. Sci. USA 1975, 72, 1276–1280. [Google Scholar] [CrossRef] [PubMed]
- Batterson, W.; Roizman, B. Characterization of the herpes simplex virion-associated factor responsible for the induction of alpha genes. J. Virol. 1983, 46, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Stelitano, D.; Franci, G.; Chianese, A.; Galdiero, S.; Morelli, G.; Galdiero, M. HSV membrane glycoproteins, their function in viral entry and their use in vaccine studies. In Amino Acids, Peptides and Proteins; Ryadnov, M., Hudecz, F., Eds.; The Royal Society of Chemistry: Cambridge, UK, 2019; Volume 43, pp. 14–43. [Google Scholar]
- Zhao, J.; Qin, C.; Liu, Y.; Rao, Y.; Feng, P. Herpes Simplex Virus and Pattern Recognition Receptors: An Arms Race. Front. Immunol. 2021, 11, 613799. [Google Scholar] [CrossRef]
- Tognarelli, E.I.; Palomino, T.F.; Corrales, N.; Bueno, S.M.; Kalergis, A.M.; Gonzalez, P.A. Herpes Simplex Virus Evasion of Early Host Antiviral Responses. Front. Cell Infect. Microbiol. 2019, 9, 127. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Rao, Y.; Tian, M.; Zhang, S.; Feng, P. Modulation of Innate Immune Signaling Pathways by Herpesviruses. Viruses 2019, 11, 572. [Google Scholar] [CrossRef]
- Paludan, S.R.; Bowie, A.G.; Horan, K.A.; Fitzgerald, K.A. Recognition of herpesviruses by the innate immune system. Nat. Rev. Immunol. 2011, 11, 143–154. [Google Scholar] [CrossRef]
- Alandijany, T. Host Intrinsic and Innate Intracellular Immunity During Herpes Simplex Virus Type 1 (HSV-1) Infection. Front. Microbiol. 2019, 10, 2611. [Google Scholar] [CrossRef]
- Chen, N.; Xia, P.; Li, S.; Zhang, T.; Wang, T.T.; Zhu, J. RNA sensors of the innate immune system and their detection of pathogens. IUBMB Life 2017, 69, 297–304. [Google Scholar] [CrossRef]
- Lin, Y.; Zheng, C. A Tug of War: DNA-Sensing Antiviral Innate Immunity and Herpes Simplex Virus Type I Infection. Front. Microbiol. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Yum, S.; Li, M.; Fang, Y.; Chen, Z.J. TBK1 recruitment to STING activates both IRF3 and NF-κB that mediate immune defense against tumors and viral infections. Proc. Natl. Acad. Sci. USA 2021, 118, e2100225118. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Heylbroeck, C.; Pitha, P.M.; Hiscott, J. Virus-dependent phosphorylation of the IRF-3 transcription factor regulates nuclear translocation, transactivation potential, and proteasome-mediated degradation. Mol. Cell. Biol. 1998, 18, 2986–2996. [Google Scholar] [CrossRef] [PubMed]
- García, M.A.; Gil, J.; Ventoso, I.; Guerra, S.; Domingo, E.; Rivas, C.; Esteban, M. Impact of protein kinase PKR in cell biology: From antiviral to antiproliferative action. Microbiol. Mol. Biol. Rev. 2006, 70, 1032–1060. [Google Scholar] [CrossRef]
- Clemens, M.J. PKR—A protein kinase regulated by double-stranded RNA. Int. J. Biochem. Cell Biol. 1997, 29, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.A.; Meurs, E.F.; Esteba, M. The dsRNA protein kinase PKR: Virus and cell control. Biochimie 2006, 89, 799–811. [Google Scholar] [CrossRef]
- Borden, E.C.; Williams, B.R. Interferon-stimulated genes and their protein products: What and how? J. Interferon Cytokine Res. 2011, 31, 1–4. [Google Scholar] [CrossRef]
- Balachandran, S.; Roberts, P.C.; Brown, L.E.; Truong, H.; Pattnaik, A.K.; Archer, D.R.; Barber, G.N. Essential role for the dsRNA-dependent protein kinase PKR in innate immunity to viral infection. Immunity 2000, 13, 129–141. [Google Scholar] [CrossRef]
- Dar, A.C.; Dever, T.E.; Sicheri, F. Higher-order substrate recognition of eIF2alpha by the RNA-dependent protein kinase PKR. Cell 2005, 122, 887–900. [Google Scholar] [CrossRef]
- Dey, M.; Cao, C.; Dar, A.C.; Tamura, T.; Ozato, K.; Sicheri, F.; Dever, T.E. Mechanistic link between PKR dimerization, autophosphorylation, and eIF2alpha substrate recognition. Cell 2005, 122, 901–913. [Google Scholar] [CrossRef]
- Levy, D.E.; Marié, I.J.; Durbin, J.E. Induction and function of type I and III interferon in response to viral infection. COVIRO 2011, 1, 476–486. [Google Scholar] [CrossRef]
- Zhang, P.; Samuel, C.E. Induction of protein kinase PKR-dependent activation of interferon regulatory factor 3 by vaccinia virus occurs through adapter IPS-1 signaling. J. Biol. Chem. 2008, 283, 34580–34587. [Google Scholar] [CrossRef] [PubMed]
- Barber, G.N. The dsRNA-dependent protein kinase, PKR and cell death. Cell Death Differ. 2005, 12, 563–570. [Google Scholar] [CrossRef] [PubMed]
- Samuel, C.E.; Kuhen, K.L.; George, C.X.; Ortega, L.G.; Rende-Fournier, R.; Tanaka, H. The PKR protein kinase—An interferon-inducible regulator of cell growth and differentiation. Int. J. Hematol. 1997, 65, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.R. PKR; a sentinel kinase for cellular stress. Oncogene 1999, 18, 6112–6120. [Google Scholar] [CrossRef]
- Tallóczy, Z.; Virgin, H., IV.; Levine, B. PKR-dependent autophagic degradation of herpes simplex virus type 1. Autophagy 2006, 2, 24–29. [Google Scholar] [CrossRef]
- Jagus, R.; Joshi, B.; Barber, G.N. PKR, apoptosis and cancer. Int. J. Biochem. Cell Biol. 1999, 31, 123–138. [Google Scholar] [CrossRef]
- Pan, S.; Liu, X.; Ma, Y.; Cao, Y.; He, B. Herpes Simplex Virus 1 γ134.5 Protein Inhibits STING Activation That Restricts Viral Replication. J. Virol. J. 2018, 92, e01015-18. [Google Scholar]
- Sharma, N.; Wang, C.; Kessler, P.; Sen, G.C. Herpes simplex virus 1 evades cellular antiviral response by inducing microRNA-24, which attenuates STING synthesis. PLoS Pathog. 2021, 17, e1009950. [Google Scholar] [CrossRef]
- Watanabe, T.; Hiasa, Y.; Tokumoto, Y.; Hirooka, M.; Abe, M.; Ikeda, Y.; Matsuura, B.; Chung, R.T.; Onji, M. Protein Kinase R Modulates c-Fos and c-Jun Signaling to Promote Proliferation of Hepatocellular Carcinoma with Hepatitis C Virus Infection. PLoS ONE 2013, 8, e67750. [Google Scholar] [CrossRef]
- Pennisi, R.; Musarra-Pizzo, M.; Lei, Z.; Zhou, G.G.; Sciortino, M.T. VHS, US3 and UL13 viral tegument proteins are required for Herpes Simplex Virus-Induced modification of protein kinase R. Sci. Rep. 2020, 10, 5580. [Google Scholar] [CrossRef]
- EL-Aguel, A.; Pennisi, R.; Smeriglio, A.; Kallel, I.; Tamburello, M.P.; D’Arrigo, M.; Barreca, D.; Gargouri, A.; Trombetta, D.; Mandalari, G.; et al. Punica granatum Peel and Leaf Extracts as Promising Strategies for HSV-1 Treatment. Viruses 2022, 14, 2639. [Google Scholar] [CrossRef] [PubMed]
- Zuo, W.; Wakimoto, M.; Kozaiwa, N.; Shirasaka, Y.; Oh, S.-W.; Fujiwara, S.; Miyachi, H.; Kogure, M.; Kato, H.; Fujita, T. PKR and TLR3 trigger distinct signals that coordinate the induction of antiviral apoptosis. Cell Death Dis. 2022, 13, 707. [Google Scholar] [CrossRef] [PubMed]
- Rojas, J.M.; Alejo, A.; Martín, V.; Sevilla, N. Viral pathogen-induced mechanisms to antagonize mammalian interferon (IFN) signaling pathway. Cell. Mol. Life Sci. 2021, 78, 1423–1444. [Google Scholar] [CrossRef]
- Huang, B.; Qi, Z.T.; Xu, Z.; Nie, P. Global characterization of interferon regulatory factor (IRF) genes in vertebrates: Glimpse of the diversification in evolution. BMC Immunol. 2010, 11, 22. [Google Scholar] [CrossRef] [PubMed]
- Meurs, E.; Chong, K.; Galabru, J.; Thomas, N.S.; Kerr, I.M.; Williams, B.R.; Hovanessian, A.G. Molecular cloning and characterization of the human double stranded RNA-activated protein kinase induced by interferon. Cell 1990, 62, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Lagunoff, M.; Roizman, B. Expression of a herpes simplex virus 1 open reading frame antisense to the gamma(1)34.5 gene and transcribed by an RNA 3′ coterminal with the unspliced latency-associated transcript. J. Virol. 1994, 68, 6021–6028. [Google Scholar] [CrossRef]
- Cole, J.L. Activation of PKR: An open and shut case? Trends Biochem. Sci. 2007, 32, 57–62. [Google Scholar] [CrossRef]
- Kumar, A.; Yang, Y.L.; Flati, V.; Der, S.; Kadereit, S.; Deb, A.; Haque, J.; Reis, L.; Weissmann, C.; Williams, B.R. Deficient cytokine signaling in mouse embryo fibroblasts with a targeted deletion in the PKR gene: Role of IRF-1 and NF-κB. EMBO J. 1997, 16, 406–416. [Google Scholar] [CrossRef]
- Wong, A.H.; Tam, N.W.; Yang, Y.L.; Cuddihy, A.R.; Li, S.; Kirchhoff, S.; Hauser, H.; Decker, T.; Koromilas, A.E. Physical association between STAT1 and the interferon-inducible protein kinase PKR and implications for interferon and double-stranded RNA signaling pathways. EMBO J. 1997, 16, 1291–1304. [Google Scholar] [CrossRef]
- Kang, R.; Tang, D. PKR-dependent inflammatory signals. Sci. Signal 2012, 5, 247. [Google Scholar] [CrossRef]
- Gil, J.; Esteban, M. The interferon-induced protein kinase (PKR), triggers apoptosis through FADD-mediated activation of caspase 8 in a manner independent of Fas and TNF-alpha receptors. Oncogene 2000, 19, 3665–3674. [Google Scholar] [CrossRef]
- Carty, M.; Guy, C.; Bowie, A.G. Detection of Viral Infections by Innate Immunity. Biochem. Pharmacol. 2021, 183, 114316. [Google Scholar] [CrossRef] [PubMed]
- Fiume, G.; Vecchio, E.; De Laurentiis, A.; Trimboli, F.; Palmieri, C.; Pisano, A.; Falcone, C.; Pontoriero, M.; Rossi, A.; Scialdone, A.; et al. Human immunodeficiency virus-1 Tat activates NF-kappaB via physical interaction with IkappaB-alpha and p65. Nucleic Acids Res. 2012, 40, 3548–3562. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; He, S.; Minassian, A.; Li, J.; Feng, P. Recent advances on viral manipulation of NF-κB signaling pathway. COVIRO 2015, 15, 103–111. [Google Scholar] [CrossRef]
- Lim, K.H.; Choi, H.S.; Park, Y.K.; Park, E.S.; Shin, G.C.; Kim, D.H.; Ahn, S.H.; Kim, K.H. HBx-induced NF-kappaB signaling in liver cells is potentially mediated by the ternary complex of HBx with p22-FLIP and NEMO. PLoS ONE 2013, 8, e57331. [Google Scholar]
- Xu, M.; Katzenellenbogen, R.A.; Grandori, C.; Galloway, D.A. NFX1 plays a role in human papillomavirus type 16 E6 activation of NFkappaB activity. J. Virol. 2010, 84, 11461–11469. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Zheng, S.; Huang, Z.; Lin, Y.; Shen, Q.; Zheng, C. Herpes Simplex Virus 1 Tegument Protein UL46 Inhibits TANK-Binding Kinase 1-Mediated Signaling. mBio 2019, 10, e00919-19. [Google Scholar] [CrossRef]
- Christensen, M.H.; Jensen, S.B.; Miettinen, J.J.; Luecke, S.; Prabakaran, T.; Reinert, L.S.; Mettenleiter, T.; Chen, Z.J.; Knipe, D.M.; Sandri-Goldin, R.M.; et al. HSV-1 ICP27 targets the TBK1-activated STING signalsome to inhibit virusinduced type I IFN expression. EMBO J. 2016, 35, 1385–1399. [Google Scholar] [CrossRef]
- Zhou, R.; Zhang, Q.; Xu, P. TBK1, a central kinase in innate immune sensing of nucleic acids and beyond. Acta Biochim. Biophys. Sin. (ABBS) 2020, 52, 757–767. [Google Scholar] [CrossRef]
- Smale, S.T. Selective transcription in response to an inflammatory stimulus. Cell 2010, 19, 833–844. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Direction | si-PKR#1 (nt. 584–604) * | si-PKR#2 (nt. 786–806) | si-PKR#3 (nt. 639–658) |
|---|---|---|---|
| TARGET | GAGAATTTCCAGAAGGTGA | TTCAGGACCTCCACATGATAG | TTCAGGACCTCCACATGATAG |
| SENSE | GAGAAUUUCCAGAAGGUGA | UUCAGGACCUCCACAUGAUAG | UUCAGGACCUCCACAUGAUAG |
| ANTISENSE | UCACCUUCUGGAAAUUCUC | CUAUCAUGUGGAGGUCCUGAA | CUAUCAUGUGGAGGUCCUGAA |
| siRNA-Scrambled | |||
| SENSE | UUCUCCGAACGUGUCACGUTT | ||
| ANTISENSE | ACGUGACACGUUCGGAGAATT | ||
| Gene | Forward | Reverse |
|---|---|---|
| ICP0 | 5′-TCTGCATCCCGTGCATGAAAAC-3′ | 5′-CTGATTGCCCGTCCAGATAAAG-3′ |
| UL42 | 5′-CTCCCTCCTGAGCGTGTTTC-3′ | 5′-CACAAAGCTCGTCAGTTCGC-3′ |
| US11 | 5′-GGCTTCAGATGGCTTCGAG-3′ | 5′-GGGCGACCCAGATGTTTAC-3′ |
| TBK1 | 5′-GCAGTTTGTTTCTCTGTATGGC-3′ | 5′-AATGTTACCCCAATGCTCCA-3′ |
| IRF3 | 5′-GCCGAGGCCACTGGTGCATAT-3′ | 5′-TGGGTCGTGAGGGTCCTTGCT-3′ |
| NF-ΚB | 5′-TAAAGCCCCCAATGCATCCAAC-3′ | 5′-CCAAATCCTTCCCAGACTCCAC-3′ |
| GAPDH | 5′-GAGAAGGCTGGGGCTCAT-3′ | 5′-TGCTGATGATCTTGAGGCTG-3′ |
| PKR | 5′-CAGGCAAACAAGGTCCCATC-3′ | 5′-GCGAGTGTGCTGGTCACTAA-3′ |
| HSV-1 | 5′-CATCACCGACCCGGAGAGGGAC-3′ | 5′-GGGCCAGGCGCTTGTTGGTGTA-3′ |
| HSV-1 TaqMan probe | 5′-6FAM-ccgccgaactgagcagacacccgcgc-TAMRA 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pennisi, R.; Sciortino, M.T. HSV-1 Triggers an Antiviral Transcriptional Response during Viral Replication That Is Completely Abrogated in PKR−/− Cells. Pathogens 2023, 12, 1126. https://doi.org/10.3390/pathogens12091126
Pennisi R, Sciortino MT. HSV-1 Triggers an Antiviral Transcriptional Response during Viral Replication That Is Completely Abrogated in PKR−/− Cells. Pathogens. 2023; 12(9):1126. https://doi.org/10.3390/pathogens12091126
Chicago/Turabian StylePennisi, Rosamaria, and Maria Teresa Sciortino. 2023. "HSV-1 Triggers an Antiviral Transcriptional Response during Viral Replication That Is Completely Abrogated in PKR−/− Cells" Pathogens 12, no. 9: 1126. https://doi.org/10.3390/pathogens12091126
APA StylePennisi, R., & Sciortino, M. T. (2023). HSV-1 Triggers an Antiviral Transcriptional Response during Viral Replication That Is Completely Abrogated in PKR−/− Cells. Pathogens, 12(9), 1126. https://doi.org/10.3390/pathogens12091126