
Gut Microbiota of Apis mellifera at Selected Ontogenetic Stages and Their Immunogenic Potential during Summer

 ,
,  ,
,
Abstract

1. Introduction
2. Material and Methods
2.1. Honeybee Sampling
2.2. Gut Dissection from the Whole Intestinal Tract of Honeybees
2.3. DNA Extraction
2.4. Sample Preparation, 16s Sequencing, and Taxonomic Analysis
2.5. Analysis in QIIME2
2.6. Bacterial Profiles
2.7. Venn Diagrams
2.8. Microbiome Profiling Based on Gram staining
2.9. Statistical Analysis
3. Results
3.1. Reads Profiling for Microbial Community
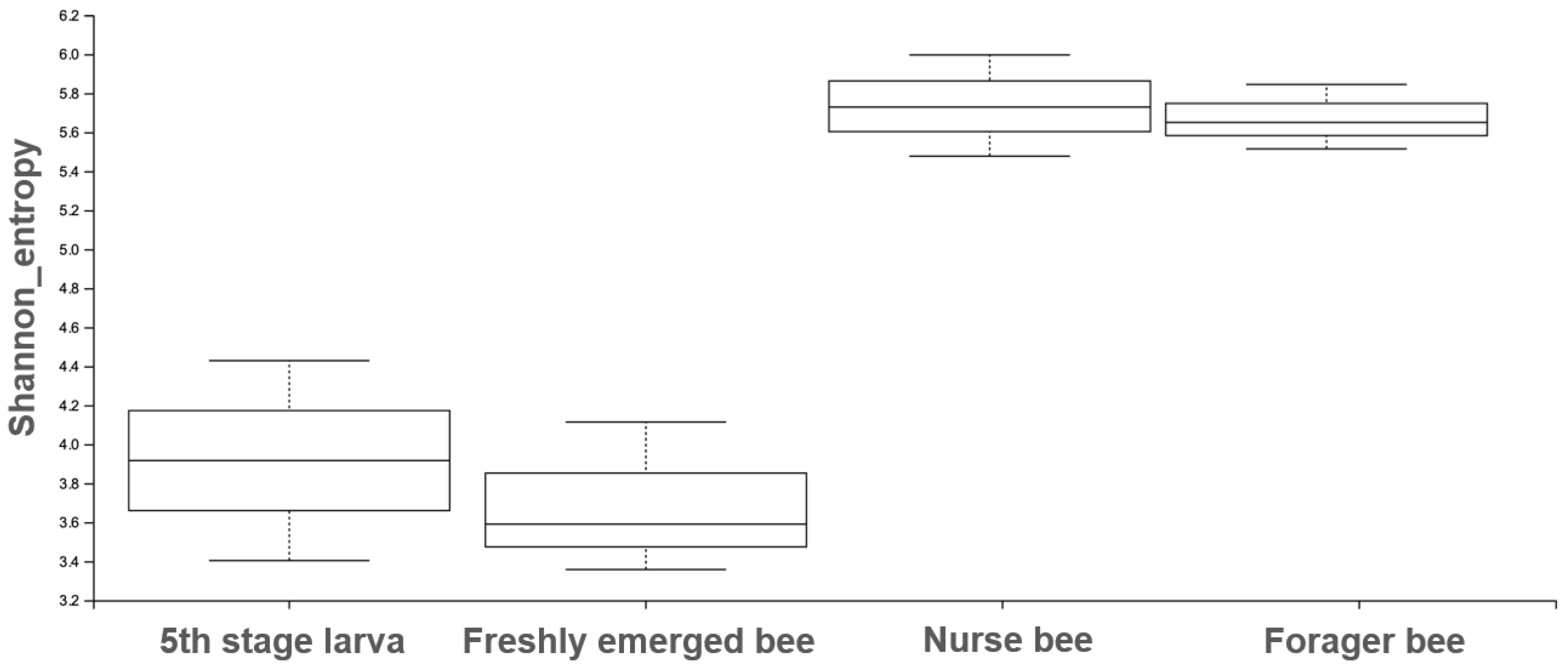
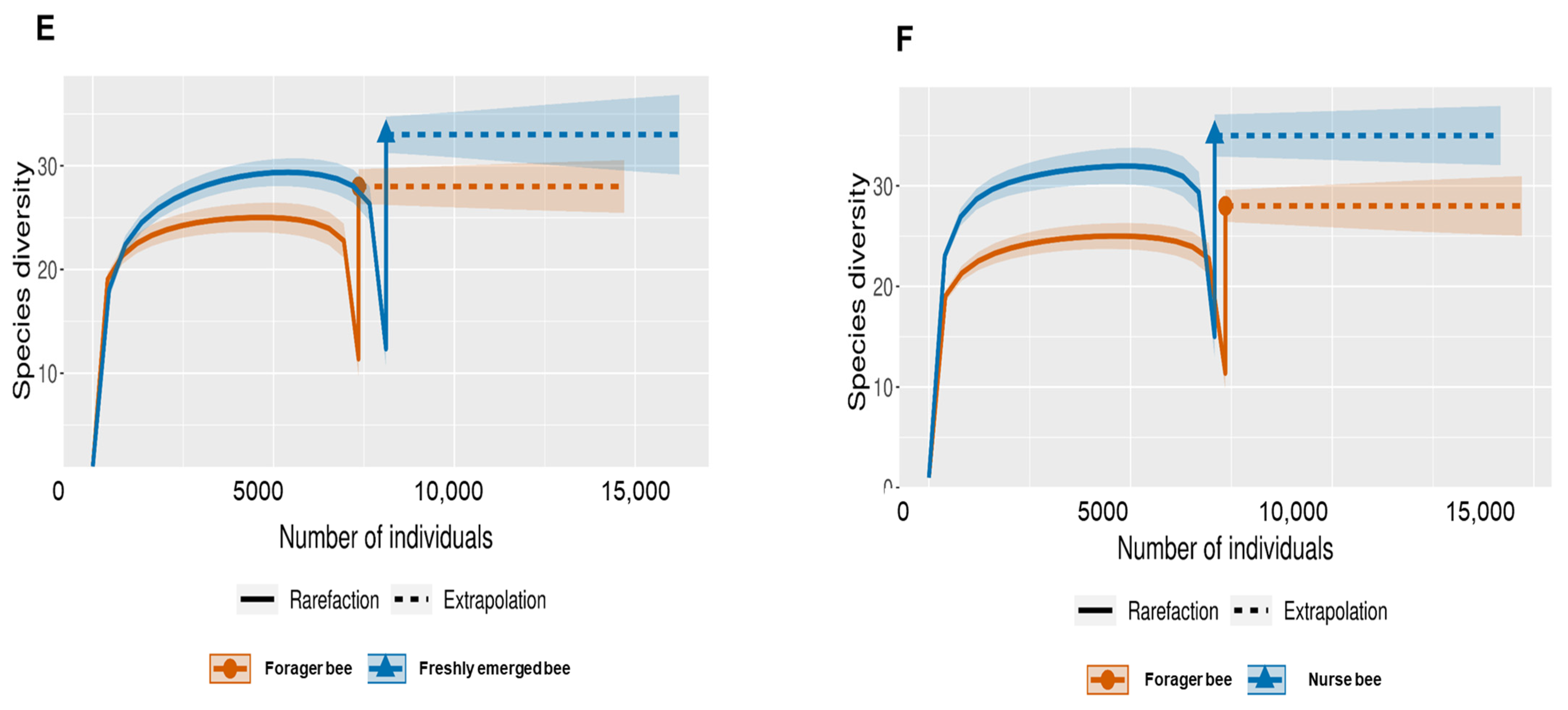
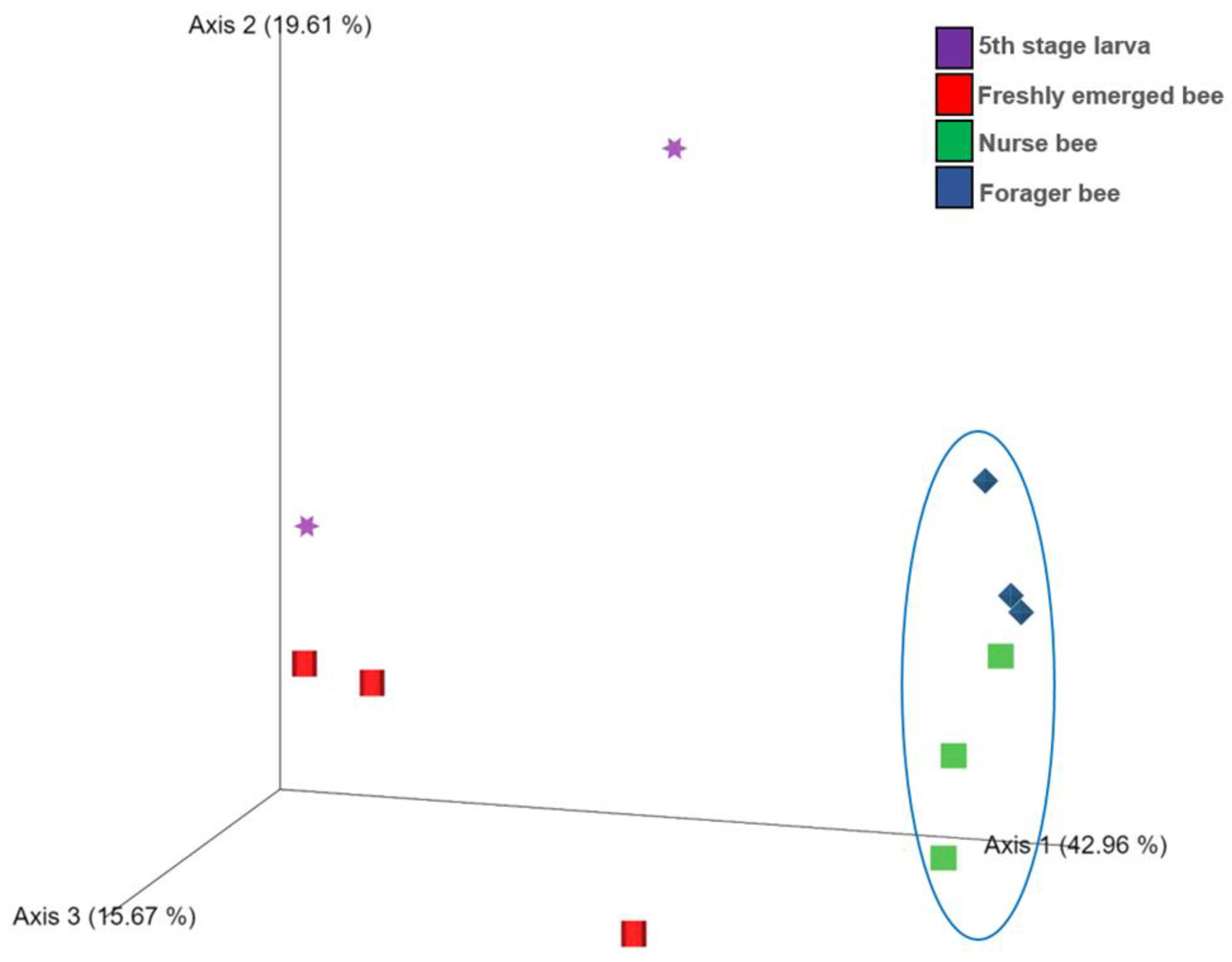
3.2. Gut Microbiota Diversity and Richness between the Age Groups
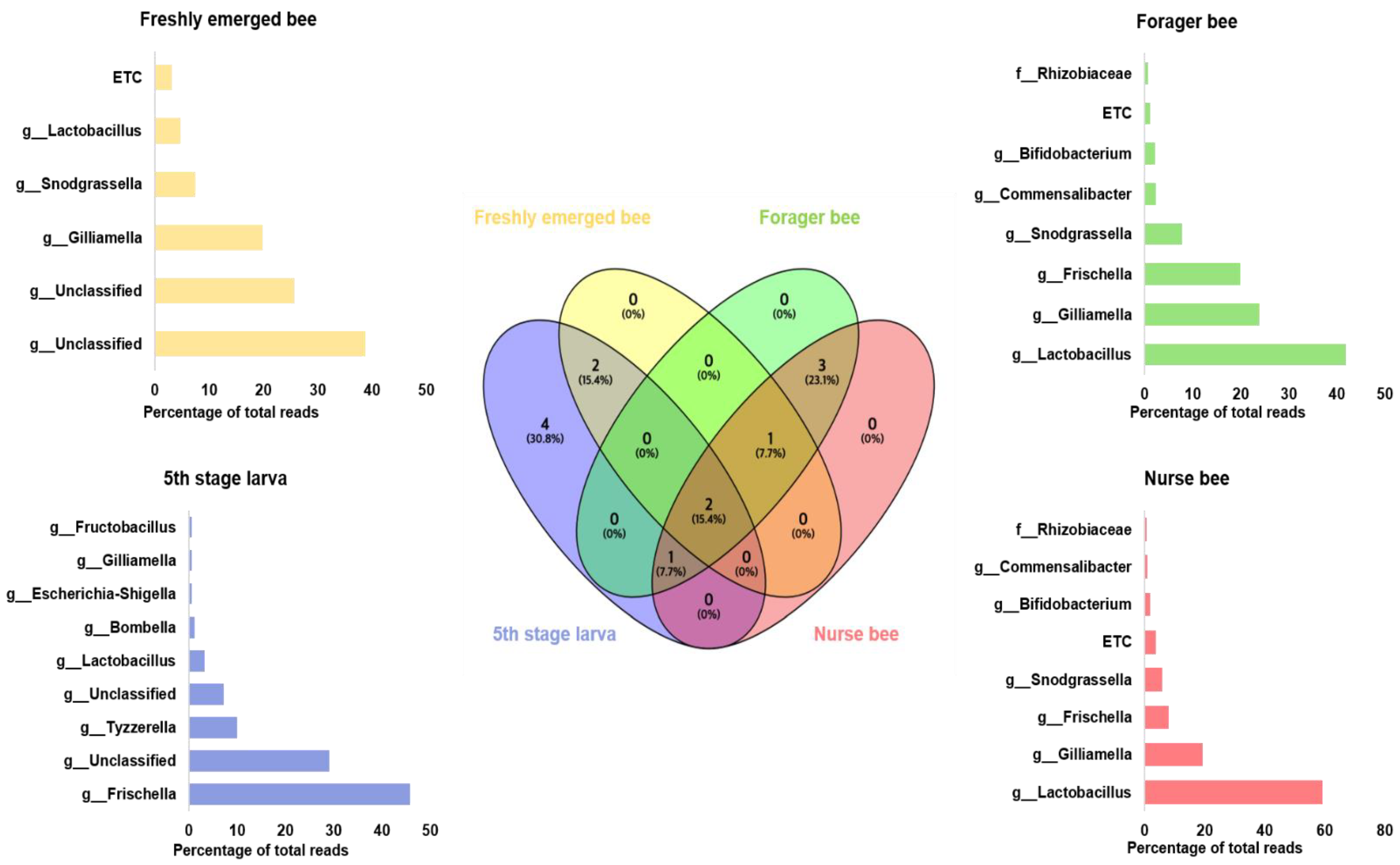
3.3. A. mellifera Gut Microbiota Abundance Distribution Based on Age
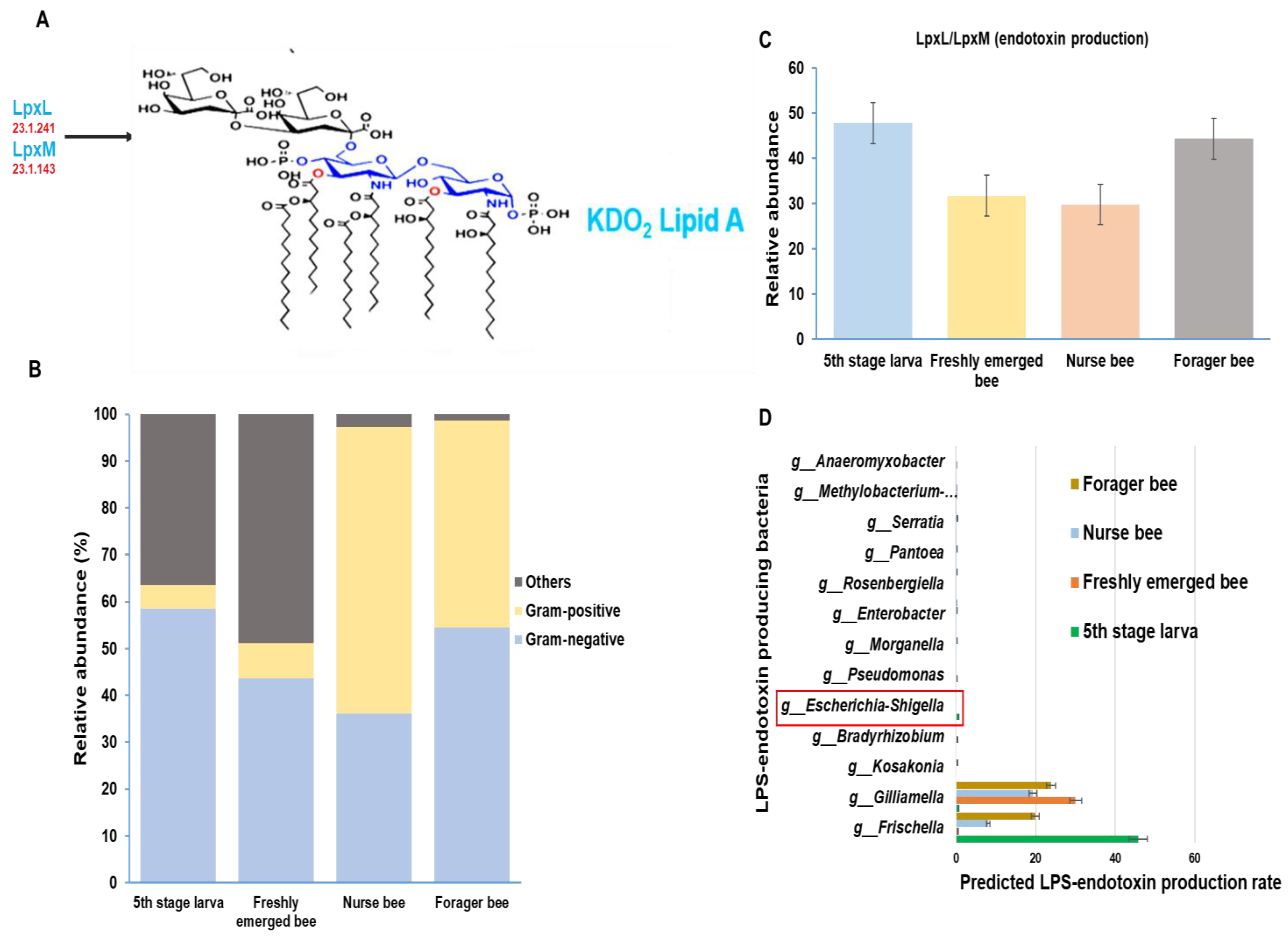
3.4. Gut Microbiota of A. mellifera Based on Gram Positivity and Gram Negativity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoover, S.E.; Ovinge, L.P. Pollen collection, honey production, and pollination services: Managing honey bees in an agricultural setting. J. Econ. Entomol. 2018, 111, 1509–1516. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T.J. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B 2007, 274, 303–313. [Google Scholar] [CrossRef]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Chandra, V.; Singh, A.K.; Singh, S.; Kumar, A.; Tiwari, D.K.; Sahay, R.; Maurya, R.C.; Singh, A. Management of Colony Collapse Disorder in Honeybee (Apis mellifera): A Farmer’s Friendly Approach. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 2557–2568. [Google Scholar] [CrossRef]
- Stokstad, E. The case of the empty hives. Science 2007, 316, 970–972. [Google Scholar] [CrossRef]
- Colin, T.; Meikle, W.G.; Paten, A.M.; Barron, A.B. Long-term dynamics of honey bee colonies following exposure to chemical stress. Sci. Total Environ. 2019, 677, 660–670. [Google Scholar] [CrossRef]
- Owen, R. Role of Human Action in the Spread of Honey Bee (Hymenoptera: Apidae) Pathogens. J. Econ. Entomol. 2017, 110, 797–801. [Google Scholar] [CrossRef]
- Le Conte, Y.; Navajas, M. Climate change: Impact on honey bee populations and diseases. Rev. Sci. Tech.-Off. Int. Des Epizoot. 2008, 27, 499–510. [Google Scholar]
- Raymann, K.; Moran, N.A. The role of the gut microbiome in health and disease of adult honey bee workers. Curr. Opin. Insect Sci. 2018, 26, 97–104. [Google Scholar] [CrossRef]
- Moran, N.A.; Plague, G.R.; Sandström, J.P.; Wilcox, J.L. A genomic perspective on nutrient provisioning by bacterial symbionts of insects. Proc. Natl. Acad. Sci. USA 2003, 100, 14543–14548. [Google Scholar] [CrossRef]
- Tola, Y.H.; Waweru, J.W.; Hurst, G.D.; Slippers, B.; Paredes, J.C. Characterization of the Kenyan honey bee (Apis mellifera) gut microbiota: A first look at tropical and Sub-Saharan African bee associated microbiomes. Microorganisms 2020, 8, 1721. [Google Scholar] [CrossRef]
- Hamdi, C.; Balloi, A.; Essanaa, J.; Crotti, E.; Gonella, E.; Raddadi, N.; Ricci, I.; Boudabous, A.; Borin, S.; Manino, A.; et al. Gut microbiome dysbiosis and honeybee health. J. Appl. Èntomol. 2011, 135, 524–533. [Google Scholar] [CrossRef]
- Jeyaprakash, A.; Hoy, M.A.; Allsopp, M.H. Bacterial diversity in worker adults of Apis mellifera capensis and Apis mellifera scutellata (Insecta: Hymenoptera) assessed using 16S rRNA sequences. J. Invertebr. Pathol. 2003, 84, 96–103. [Google Scholar] [CrossRef]
- Babendreier, D.; Joller, D.; Romeis, J.; Bigler, F.; Widmer, F. Bacterial community structures in honeybee intestines and their response to two insecticidal proteins. FEMS Microbiol. Ecol. 2007, 59, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Engel, P.; Moran, N.A. Functional and evolutionary insights into the simple yet specific gut microbiota of the honey bee from metagenomic analysis. Gut Microbes 2013, 4, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Mattila, H.R.; Rios, D.; Walker-Sperling, V.E.; Roeselers, G.; Newton, I.L. Characterization of the active microbiotas associated with honey bees reveals healthier and broader communities when colonies are genetically diverse. PLoS ONE 2012, 7, e32962. [Google Scholar] [CrossRef] [PubMed]
- Crailsheim, K.J. The flow of jelly within a honeybee colony. J. Comp. Physiol. B 1992, 162, 681–689. [Google Scholar] [CrossRef]
- Robinson, G.E. Regulation of division of labor in insect societies. Annu. Rev. Entomol. 1992, 37, 637–665. [Google Scholar] [CrossRef]
- Gätschenberger, H.; Azzami, K.; Tautz, J.; Beier, H. Antibacterial immune competence of honey bees (Apis mellifera) is adapted to different life stages and environmental risks. PLoS ONE 2013, 8, e66415. [Google Scholar] [CrossRef]
- Brune, A.; Friedrich, M. Microecology of the termite gut: Structure and function on a microscale. Curr. Opin. Microbiol. 2000, 3, 263–269. [Google Scholar] [CrossRef]
- Robinson, C.J.; Schloss, P.; Ramos, Y.; Raffa, K.; Handelsman, J. Robustness of the bacterial community in the cabbage white butterfly larval midgut. Microb. Ecol. 2010, 59, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Maigoro, A.Y.; Muhammad, M.; Bello, B.; Useh, U.; Lee, S.J. Exploration of Gut Microbiome Research in Africa: A Scoping Review. J. Med. Food 2023, 26, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Jayakodi, M.; Jung, J.W.; Park, D.; Ahn, Y.-J.; Lee, S.-C.; Shin, S.-Y.; Shin, C.; Yang, T.-J.; Kwon, H.W. Genome-wide characterization of long intergenic non-coding RNAs (lincRNAs) provides new insight into viral diseases in honey bees Apis cerana and Apis mellifera. BMC Genom. 2015, 16, 680. [Google Scholar] [CrossRef] [PubMed]
- Dillon, R.J.; Dillon, V. The gut bacteria of insects: Nonpathogenic interactions. Annu. Rev. Entomol. 2004, 49, 71–92. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Jung, J.W.; Choi, B.S.; Jayakodi, M.; Lee, J.; Lim, J.; Yu, Y.; Choi, Y.S.; Lee, M.L.; Park, Y.; et al. Uncovering the novel characteristics of Asian honey bee, Apis cerana, by whole genome sequencing. BMC Genom. 2015, 16, 1. [Google Scholar] [CrossRef] [PubMed]
- Tanada, Y.; Kaya, H. Associations between insects and nonpathogenics microorganisms. Insect Pathol. 1993, 12–51. [Google Scholar]
- Dong, Z.-X.; Li, H.-Y.; Chen, Y.-F.; Wang, F.; Deng, X.-Y.; Lin, L.-B.; Zhang, Q.-L.; Li, J.-L.; Guo, J. Colonization of the gut microbiota of honey bee (Apis mellifera) workers at different developmental stages. Microbiol. Res. 2020, 231, 126370. [Google Scholar] [CrossRef]
- Yun, B.-R.; Truong, A.-T.; Choi, Y.S.; Lee, M.Y.; Kim, B.Y.; Seo, M.; Yoon, S.-S.; Yoo, M.-S.; Van Quyen, D.; Cho, Y.S. Comparison of the gut microbiome of sacbrood virus-resistant and-susceptible Apis cerana from South Korea. Sci. Rep. 2022, 12, 10010. [Google Scholar] [CrossRef]
- Strand, M.R. The insect cellular immune response. Insect Sci. 2008, 15, 1–14. [Google Scholar] [CrossRef]
- Censoplano, N.; Epting, C.L.; Coates, B.M. The role of the innate immune system in sepsis. Clin. Pediatr. Emerg. Med. 2014, 15, 169–176. [Google Scholar] [CrossRef]
- Casteels, P.; Ampe, C.; Jacobs, F.; Tempst, P. Functional and chemical characterization of Hymenoptaecin, an antibacterial polypeptide that is infection-inducible in the honeybee (Apis mellifera). J. Biol. Chem. 1993, 268, 7044–7054. [Google Scholar] [CrossRef]
- Casteels, P.; Ampe, C.; Jacobs, F.; Vaeck, M.; Tempst, P. Apidaecins: Antibacterial peptides from honeybees. EMBO J. 1989, 8, 2387–2391. [Google Scholar] [CrossRef]
- Kwong, W.K.; Mancenido, A.L.; Moran, N.A. Immune system stimulation by the native gut microbiota of honey bees. R Soc. Open Sci. 2017, 4, 170003. [Google Scholar] [CrossRef]
- Marmaras, V.J.; Lampropoulou, M. Regulators and signalling in insect haemocyte immunity. Cell Signal. 2009, 21, 186–195. [Google Scholar] [CrossRef]
- Walderdorff, L.; Laval-Gilly, P.; Bonnefoy, A.; Falla-Angel, J. Imidacloprid intensifies its impact on honeybee and bumblebee cellular immune response when challenged with LPS (lippopolysacharide) of Escherichia coli. J. Insect Physiol. 2018, 108, 17–24. [Google Scholar] [CrossRef]
- Lavine, M.; Strand, M.R. Insect hemocytes and their role in immunity. Insect Biochem. Mol. Biol. 2002, 32, 1295–1309. [Google Scholar] [CrossRef]
- Feldhaar, H.; Gross, R. Immune reactions of insects on bacterial pathogens and mutualists. Microbes Infect. 2008, 10, 1082–1088. [Google Scholar] [CrossRef] [PubMed]
- Munford, R.S.; Hall, C.L. Detoxification of bacterial lipopolysaccharides (endotoxins) by a human neutrophil enzyme. Science 1986, 234, 203–205. [Google Scholar] [CrossRef] [PubMed]
- Vatanen, T.; Kostic, A.D.; d’Hennezel, E.; Siljander, H.; Franzosa, E.A.; Yassour, M.; Kolde, R.; Vlamakis, H.; Arthur, T.D.; Hämäläinen, A.-M.; et al. Variation in microbiome LPS immunogenicity contributes to autoimmunity in humans. Cell 2016, 165, 842–853. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Goulson, D.; Pennacchio, F.; Nazzi, F.; Goka, K.; Desneux, N. Are bee diseases linked to pesticides?—A brief review. Environ. Int. 2016, 89, 7–11. [Google Scholar] [CrossRef]
- Hroncova, Z.; Havlik, J.; Killer, J.; Doskocil, I.; Tyl, J.; Kamler, M.; Titera, D.; Hakl, J.; Mrazek, J.; Bunesova, V. Variation in honey bee gut microbial diversity affected by ontogenetic stage, age and geographic location. PLoS ONE 2015, 10, e0118707. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, H.-W.; Mustafa, B.; Lee, H.I.; Kwon, H.W. The relationships between microbiome diversity and epidemiology in domestic species of malaria-mediated mosquitoes of Korea. Sci. Rep. 2023, 13, 9081. [Google Scholar] [CrossRef]
- UniProt. UniProt: The universal protein knowledgebase in 2021. J. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Maigoro, A.Y.; Lee, S. Gut Microbiome-Based Analysis of Lipid A Biosynthesis in Individuals with Autism Spectrum Disorder: An In Silico Evaluation. Nutrients 2021, 13, 688. [Google Scholar] [CrossRef]
- RDC Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Martinson, V.G.; Moy, J.; Moran, N.A. Establishment of characteristic gut bacteria during development of the honeybee worker. Appl. Environ. Microbiol. 2012, 78, 2830–2840. [Google Scholar] [CrossRef]
- Mohr, K.I.; Tebbe, C.C. Diversity and phylotype consistency of bacteria in the guts of three bee species (Apoidea) at an oilseed rape field. Environ. Microbiol. 2006, 8, 258–272. [Google Scholar] [CrossRef]
- Janashia, I.; Alaux, C. Specific immune stimulation by endogenous bacteria in honey bees (Hymenoptera: Apidae). J. Econ. Entomol. 2016, 109, 1474–1477. [Google Scholar] [CrossRef]
- Härer, L.; Hilgarth, M.; Ehrmann, M.A. Comparative genomics of acetic acid bacteria within the genus Bombella in light of beehive habitat adaptation. Microorganisms 2022, 10, 1058. [Google Scholar] [CrossRef]
- Bulatov, T.; Gensel, S.; Mainz, A.; Dang, T.; Koller, T.O.; Voigt, K.; Ebeling, J.; Wilson, D.N.; Genersch, E.; Sussmuth, R.D. Total Synthesis and Biological Evaluation of Paenilamicins from the Honey Bee Pathogen Paenibacillus larvae. J. Am. Chem. Soc. 2022, 144, 288–296. [Google Scholar] [CrossRef]
- Turroni, F.; Van Sinderen, D.; Ventura, M. Genomics and ecological overview of the genus Bifidobacterium. Int. J. Food Microbiol. 2011, 149, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Reuter, G. The Lactobacillus and Bifidobacterium microflora of the human intestine: Composition and succession. Curr. Issues Intest. Microbiol. 2001, 2, 43–53. [Google Scholar] [PubMed]
- Olofsson, T.C.; Vásquez, A. Detection and identification of a novel lactic acid bacterial flora within the honey stomach of the honeybee Apis mellifera. Curr. Microbiol. 2008, 57, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Forsgren, E.; Olofsson, T.C.; Vásquez, A.; Fries, I. Novel lactic acid bacteria inhibiting Paenibacillus larvae in honey bee larvae. Apidologie (Celle) 2010, 41, 99–108. [Google Scholar] [CrossRef]
- Wu, M.; Sugimura, Y.; Takaya, N.; Takamatsu, D.; Kobayashi, M.; Taylor, D.; Yoshiyama, M. Characterization of bifidobacteria in the digestive tract of the Japanese honeybee, Apis cerana japonica. J. Invertebr. Pathol. 2013, 112, 88–93. [Google Scholar] [CrossRef]
- Copeland, D.C.; Maes, P.W.; Mott, B.M.; Anderson, K.E. Changes in gut microbiota and metabolism associated with phenotypic plasticity in the honey bee Apis mellifera. Front. Microbiol. 2022, 13, 1059001. [Google Scholar] [CrossRef]
- Trumbo, S.T.; Huang, Z.-Y.; Robinson, G.E. Division of labor between undertaker specialists and other middle-aged workers in honey bee colonies. Behav. Ecol. Sociobiol. 1997, 41, 151–163. [Google Scholar] [CrossRef]
- Engel, P.; Martinson, V.G.; Moran, N.A. Functional diversity within the simple gut microbiota of the honey bee. Proc. Natl. Acad. Sci. USA 2012, 109, 11002–11007. [Google Scholar] [CrossRef]
- Butler, È.; Alsterfjord, M.; Olofsson, T.C.; Karlsson, C.; Malmström, J.; Vásquez, A. Proteins of novel lactic acid bacteria from Apis mellifera mellifera: An insight into the production of known extra-cellular proteins during microbial stress. BMC Microbiol. 2013, 13, 235. [Google Scholar] [CrossRef]
- Guo, J.; Wu, J.; Chen, Y.; Evans, J.D.; Dai, R.; Luo, W.; Li, J. Characterization of gut bacteria at different developmental stages of Asian honey bees, Apis cerana. J. Invertebr. Pathol. 2015, 127, 110–114. [Google Scholar] [CrossRef]
- Gkantiragas, A.G.; Gabrielli, J.J. A Meta-Analysis of the 16S-rRNA Gut Microbiome Data in Honeybees (Apis Mellifera). bioRxiv 2021. [Google Scholar] [CrossRef]
- Bleau, N.; Bouslama, S.; Giovenazzo, P.; Derome, N. Dynamics of the honeybee (Apis mellifera) gut microbiota throughout the overwintering period in Canada. Microorganisms 2020, 8, 1146. [Google Scholar] [CrossRef]
- Cornman, R.S.; Tarpy, D.R.; Chen, Y.; Jeffreys, L.; Lopez, D.; Pettis, J.S.; vanEngelsdorp, D.; Evans, J.D. Pathogen webs in collapsing honey bee colonies. PLoS ONE 2012, 7, e43562. [Google Scholar] [CrossRef]
- Servin, A.L. Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens. FEMS Microbiol. 2004, 28, 405–440. [Google Scholar] [CrossRef]
- Steele, M.I.; Motta, E.V.; Gattu, T.; Martinez, D.; Moran, N.A. The gut microbiota protects bees from invasion by a bacterial pathogen. Microbiol. Spectr. 2021, 9, e0039421. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.E.; Martinson, V.G.; Urban-Mead, K.; Moran, N.A. Routes of acquisition of the gut microbiota of the honey bee Apis mellifera. Appl. Environ. Microbiol. 2014, 80, 7378–7387. [Google Scholar] [CrossRef] [PubMed]
- Farhana, A.; Khan, Y.S. Biochemistry, lipopolysaccharide. In StatPearls [Internet]; StatPearls Publishing: St. Petersburg, FL, USA, 2022. [Google Scholar]
- Richard, F.J.; Aubert, A.; Grozinger, C.M. Modulation of social interactions by immune stimulation in honey bee, Apis mellifera, workers. BMC Biol. 2008, 6, 50. [Google Scholar] [CrossRef] [PubMed]
- Laughton, A.M.; Boots, M.; Siva-Jothy, M.T. The ontogeny of immunity in the honey bee, Apis mellifera L. following an immune challenge. J. Insect Physiol. 2011, 57, 1023–1032. [Google Scholar] [CrossRef]
- Byhrø, E.M.H.; Salmela, H.; Vitlic, A.; Wang, Y.; Münch, D.; Amdam, G.V. Different activation of immune-related genes in honey bee nurses and foragers (Apis mellifera). Apidologie (Celle) 2019, 50, 463–471. [Google Scholar] [CrossRef]
- Bleau, C.; Karelis, A.D.; St-Pierre, D.H.; Lamontagne, L. Crosstalk between intestinal microbiota, adipose tissue and skeletal muscle as an early event in systemic low-grade inflammation and the development of obesity and diabetes. Diabetes Metab. Res. Rev. 2015, 31, 545–561. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1 | Group 2 | H | p-Value | q-Value |
|---|---|---|---|---|
| Fifth stage larva (n = 2) | Freshly emerged bee (n = 3) | 0.33 | 0.56 | 0.67 |
| Fifth stage larva (n = 2) | Forager bee (n = 3) | 3.0 | 0.08 | 0.12 |
| Fifth stage larva (n = 2) | Nurse bee (n = 3) | 3.0 | 0.08 | 0.12 |
| Freshly emerged bee (n = 3) | Forager bee (n = 3) | 3.85 | 0.04 | 0.12 |
| Freshly emerged bee (n = 3) | Nurse bee (n = 3) | 3.85 | 0.04 | 0.12 |
| Forager bee (n = 3) | Nurse bee (n = 3) | 0.04 | 0.82 | 0.82 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maigoro, A.Y.; Lee, J.-H.; Kim, H.; Frunze, O.; Kwon, H.-W. Gut Microbiota of Apis mellifera at Selected Ontogenetic Stages and Their Immunogenic Potential during Summer. Pathogens 2024, 13, 122. https://doi.org/10.3390/pathogens13020122
Maigoro AY, Lee J-H, Kim H, Frunze O, Kwon H-W. Gut Microbiota of Apis mellifera at Selected Ontogenetic Stages and Their Immunogenic Potential during Summer. Pathogens. 2024; 13(2):122. https://doi.org/10.3390/pathogens13020122
Chicago/Turabian StyleMaigoro, Abdulkadir Yusif, Jeong-Hyeon Lee, Hyunjee Kim, Olga Frunze, and Hyung-Wook Kwon. 2024. "Gut Microbiota of Apis mellifera at Selected Ontogenetic Stages and Their Immunogenic Potential during Summer" Pathogens 13, no. 2: 122. https://doi.org/10.3390/pathogens13020122
APA StyleMaigoro, A. Y., Lee, J.-H., Kim, H., Frunze, O., & Kwon, H.-W. (2024). Gut Microbiota of Apis mellifera at Selected Ontogenetic Stages and Their Immunogenic Potential during Summer. Pathogens, 13(2), 122. https://doi.org/10.3390/pathogens13020122