
Prodigiosin as an Antibiofilm Agent against the Bacterial Biofilm-Associated Infection of Pseudomonas aeruginosa
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Mice Ethics Statement
2.3. Isolation, Purification, Characterization, and Quantification of Prodigiosin (PG)
2.4. In Vitro Antimicrobial Activity Assay
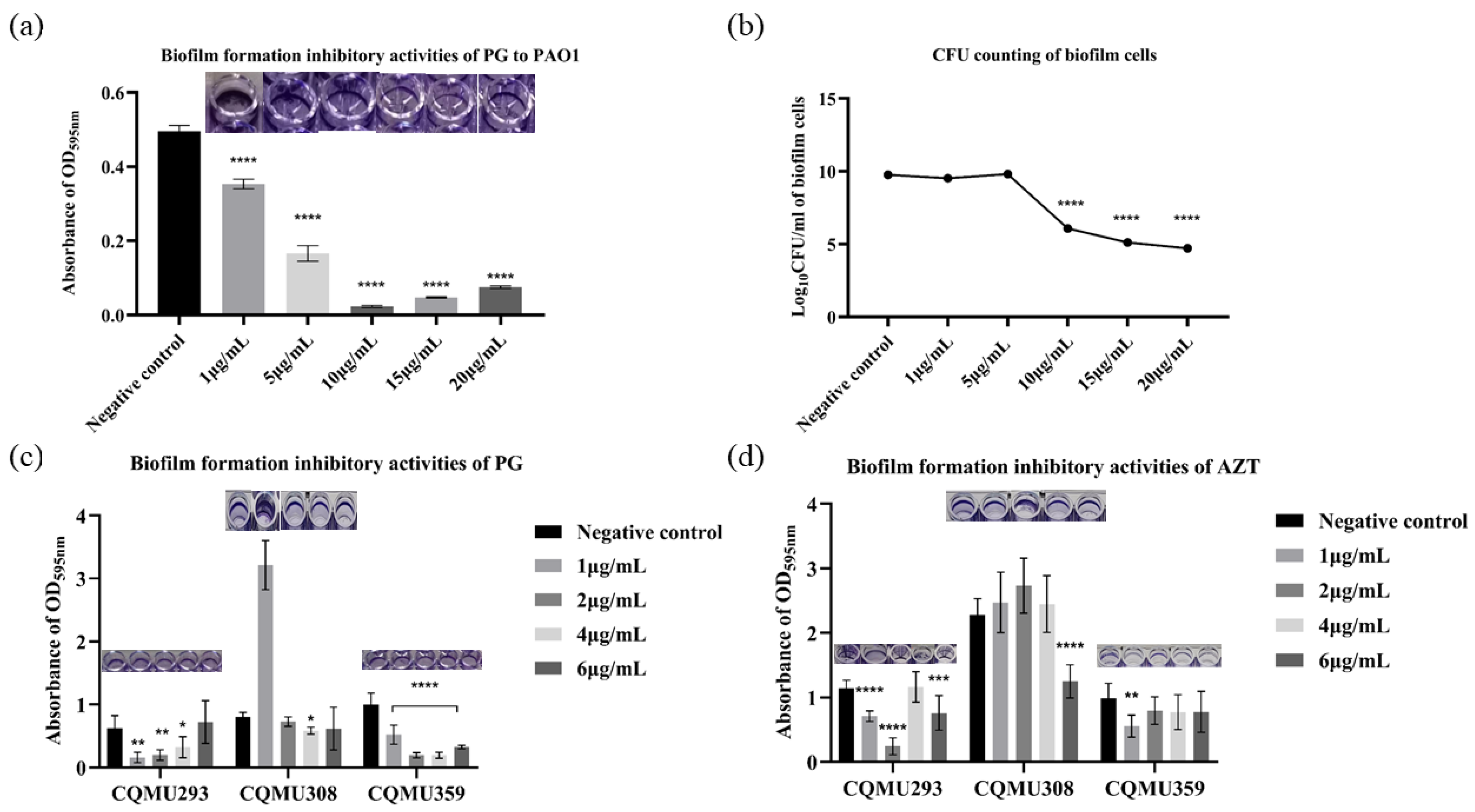
2.5. In Vitro Biofilm Formation Inhibitory Activity Assay
2.6. Bacteria Counts of Biofilms
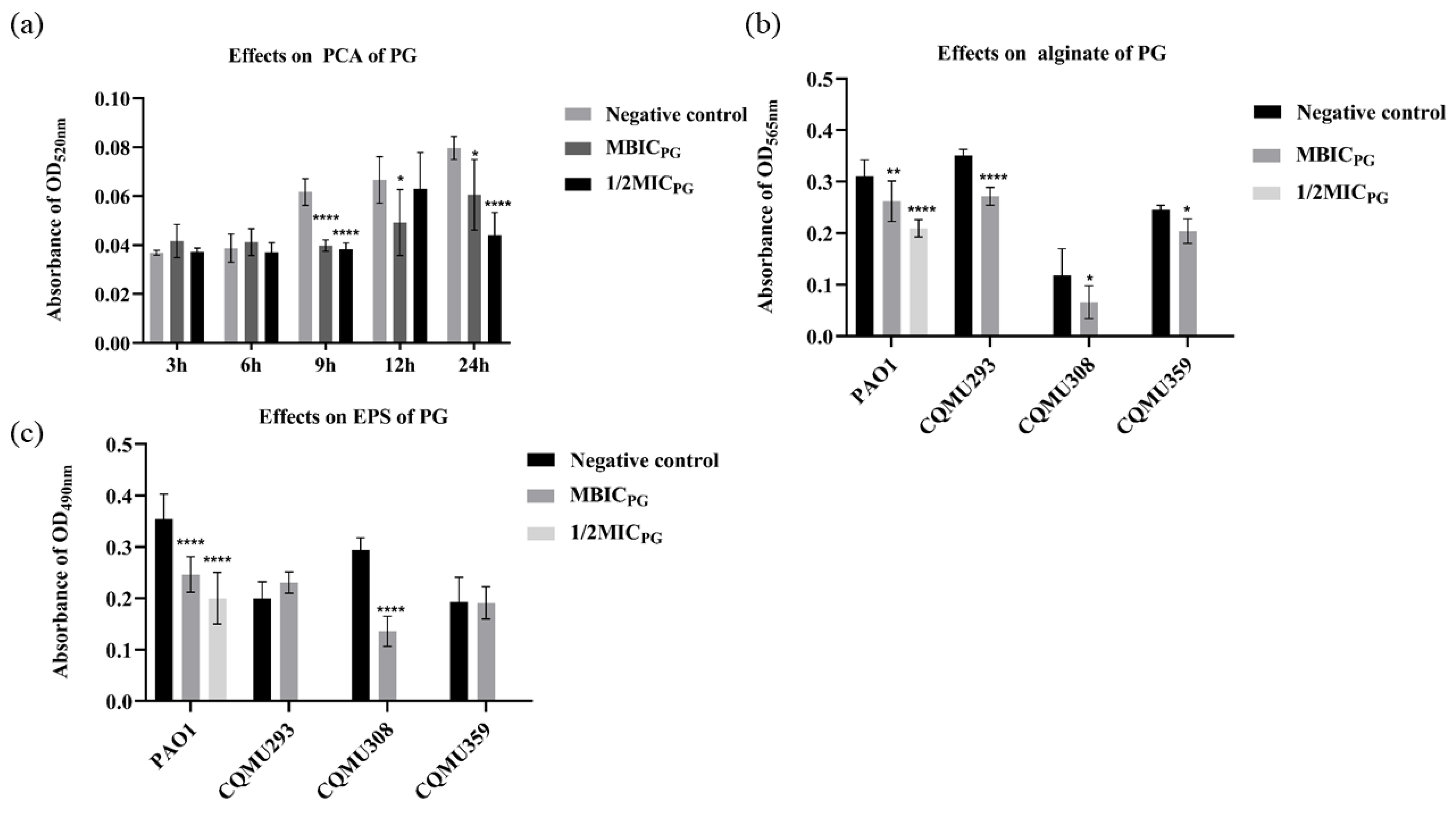
2.7. Extraction of Pyocyanin, Extracellular Polysaccharide, and Alginate
2.8. Real-Time PCR Analysis
2.9. Chronic Lung Infection of Mice Model
2.10. Statistical Analysis
3. Results
3.1. Qualitative and Quantitative Investigation of Prodigiosin (PG)
3.2. PG Showed Antimicrobial Activity against β-Lactam-Resistant Pseudomonas aeruginosa
3.3. PG Exhibited Biofilm Inhibitory Activities against P. aeruginosa In Vitro
3.4. PG Decreased the Content of Pyocyanin (PCA), Alginate, and Extracellular Polysaccharide (EPS)
3.5. Effects of PG on the Gene Expression of EPS
3.6. The Effects of PG on Chronic Lung Infection in Mice
3.6.1. In Vivo Anti-Biofilm Activity of PG
3.6.2. Effect of PG on Mice Lungs of Inflammatory Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cangui-Panchi, S.P.; Ñacato-Toapanta, A.L.; Enríquez-Martínez, L.J.; Reyes, J.; Garzon-Chavez, D.; Machado, A. Biofilm-forming microorganisms causing hospital-acquired infections from intravenous catheter: A systematic review. Curr. Res. Microb. Sci. 2022, 3, 100175. [Google Scholar] [CrossRef] [PubMed]
- Botelho, J.; Grosso, F.; Peixe, L. Antibiotic resistance in Pseudomonas aeruginosa—Mechanisms, epidemiology and evolution. Drug Resist. Updates 2019, 44, 100640. [Google Scholar] [CrossRef] [PubMed]
- Saleem, S.; Bokhari, H. Resistance profile of genetically distinct clinical Pseudomonas aeruginosa isolates from public hospitals in central Pakistan. J. Infect. Public Health 2020, 13, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Appaneal, H.J.; Caffrey, A.R.; Jiang, L.; Dosa, D.; Mermel, L.A.; LaPlante, K.L. Antibiotic resistance rates for Pseudomonas aeruginosa clinical respiratory and bloodstream isolates among the Veterans Affairs Healthcare System from 2009 to 2013. Diagn. Microbiol. Infect. Dis. 2018, 90, 311–315. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas aeruginosa Biofilms. Int. J. Mol. Sci. 2020, 21, 8671. [Google Scholar] [CrossRef]
- Atiencia-Carrera, M.B.; Cabezas-Mera, F.S.; Tejera, E.; Machado, A. Prevalence of biofilms in Candida spp. bloodstream infections: A meta-analysis. PLoS ONE 2022, 17, e0263522. [Google Scholar] [CrossRef]
- Staniszewska, M. Virulence Factors in Candida species. Current Protein Pept. Sci. 2020, 21, 313–323. [Google Scholar] [CrossRef]
- Yin, W.; Wang, Y.; Liu, L.; He, J. Biofilms: The Microbial “Protective Clothing” in Extreme Environments. Int. J. Mol. Sci. 2019, 20, 3423. [Google Scholar] [CrossRef]
- Spoering, A.L.; Lewis, K. Biofilms and planktonic cells of Pseudomonas aeruginosa have similar resistance to killing by antimicrobials. J. Bacteriol. 2001, 183, 6746–6751. [Google Scholar] [CrossRef]
- Ma, L.; Conover, M.; Lu, H.; Parsek, M.R.; Bayles, K.; Wozniak, D.J. Assembly and development of the Pseudomonas aeruginosa biofilm matrix. PLoS Pathog. 2009, 5, e1000354. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Gu, H.; Kilberg, J.B.; Ren, D. Sensitizing bacterial cells to antibiotics by shape recovery triggered biofilm dispersion. Acta Biomater. 2018, 81, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, J.; Yu, X.; Cao, R.; Hong, M.; Xu, Z.; Ren, L.J.; Wang, Y.; Zhu, H. Antimicrobial peptides fight against Pseudomonas aeruginosa at a sub-inhibitory concentration via anti-QS pathway. Bioorg. Chem. 2023, 141, 106922. [Google Scholar] [CrossRef] [PubMed]
- Urooj, S.; Mirani, Z.A.; Pathan, F.A.; Mustafa, G.; Aziz, M.; Jabeen, B.; Shah, S.H.; Ullah, A.; Hassan, N.; Khan, M.N.; et al. Silicon nanoparticles: A promising approach for control of Pseudomonas aeruginosa biofilms. Iran. J. Basic Med. Sci. 2023, 26, 1475–1483. [Google Scholar] [PubMed]
- Alghofaili, F. Use of bacterial culture supernatants as anti-biofilm agents against Pseudomonas aeruginosa and Klebsiella pneumoniae. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 1388–1397. [Google Scholar] [PubMed]
- Yan, J.; Yin, Q.; Nie, H.; Liang, J.; Liu, X.R.; Li, Y.; Xiao, H. Prodigiosin as an antibiofilm agent against multidrug-resistant Staphylococcus aureus. Biofouling 2023, 39, 444–458. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.; Li, Y.; Lu, X.L.; Yan, J.; Liu, X.R.; Yin, Q. Prodigiosin derived from chromium-resistant Serratia sp. prevents inflammation and modulates gut microbiota homeostasis in DSS-induced colitis mice. Int. Immunopharmacol. 2023, 116, 109800. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qiu, Y.; Yin, Q.; Li, X.; Bai, Q.; Li, Y.; Xiao, H. iTRAQ-based quantitative proteomic reveals proteomic changes in Serratia sp. CM01 and mechanism of Cr(VI) resistance. Ecotoxicol. Environ. Saf. 2021, 228, 112899. [Google Scholar] [CrossRef]
- Yin, Q.; Wu, S.; Wu, L.; Wang, Z.; Mu, Y.; Zhang, R.; Dong, C.; Zhou, B.; Zhao, B.; Zheng, J.; et al. A novel in silico antimicrobial peptide DP7 combats MDR Pseudomonas aeruginosa and related biofilm infections. J. Antimicrob. Chemother. 2020, 75, 3248–3259. [Google Scholar] [CrossRef]
- Song, M.J.; Bae, J.; Lee, D.S.; Kim, C.H.; Kim, J.S.; Kim, S.W.; Hong, S.I. Purification and characterization of prodigiosin produced by integrated bioreactor from Serratia sp. KH-95. J. Biosci. Bioeng. 2006, 101, 157–161. [Google Scholar] [CrossRef]
- CLSI. M100—Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Bujňáková, D.; Čuvalová, A.; Čížek, M.; Humenik, F.; Salzet, M.; Čížková, D. Canine Bone Marrow Mesenchymal Stem Cell Conditioned Media Affect Bacterial Growth, Biofilm-Associated Staphylococcus aureus and AHL-Dependent Quorum Sensing. Microorganisms 2020, 8, 1478. [Google Scholar] [CrossRef] [PubMed]
- She, P.; Wang, Y.; Liu, Y.; Tan, F.; Chen, L.; Luo, Z.; Wu, Y. Effects of exogenous glucose on Pseudomonas aeruginosa biofilm formation and antibiotic resistance. MicrobiologyOpen 2019, 8, e933. [Google Scholar] [CrossRef] [PubMed]
- Ismail, M.M.; Hassan, M.; Moawad, S.S.; Okba, M.M.; Ashour, R.M.; Fayek, N.M.; Saber, F.R. Exploring the Antivirulence Activity of Pulverulentone A, a Phloroglucinol-Derivative from Callistemon citrinus Leaf Extract, against Multi-Drug Resistant Pseudomonas aeruginosa. Antibiotics 2021, 10, 907. [Google Scholar]
- Yasuda, H.; Ajiki, Y.; Koga, T.; Kawada, H.; Yokota, T. Interaction between biofilms formed by Pseudomonas aeruginosa and clarithromycin. Antimicrob. Agents Chemother. 1993, 37, 1749–1755. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q. Studies on Luminescence Characteristics, Bacteriostatic Evaluation, and Exploration of Biofilm Formation Mechanism of lux.-tagged Recombinant Luminescent Pseudomonas aeruginosa; Northwest A&F University: Xianyang, China, 2020. [Google Scholar]
- Facchini, M.; De Fino, I.; Riva, C.; Bragonzi, A. Long term chronic Pseudomonas aeruginosa airway infection in mice. J. Vis. Exp. 2014, 85, 51019. [Google Scholar]
- Chang, C.C.; Wang, Y.H.; Chern, C.M.; Liou, K.T.; Hou, Y.C.; Peng, Y.T.; Shen, Y.C. Prodigiosin inhibits gp91(phox) and iNOS expression to protect mice against the oxidative/nitrosative brain injury induced by hypoxia-ischemia. Toxicol. Appl. Pharmacol. 2011, 257, 137–147. [Google Scholar] [CrossRef]
- Kukavica-Ibrulj, I.; Facchini, M.; Cigana, C.; Levesque, R.C.; Bragonzi, A. Assessing Pseudomonas aeruginosa virulence and the host response using murine models of acute and chronic lung infection. Methods Mol. Biol. 2014, 1149, 757–771. [Google Scholar]
- Klesney-Tait, J.; Keck, K.; Li, X.; Gilfillan, S.; Otero, K.; Baruah, S.; Meyerholz, D.K.; Varga, S.M.; Knudson, C.J.; Moninger, T.O.; et al. Transepithelial migration of neutrophils into the lung requires TREM-1. J. Clin. Investig. 2013, 123, 138–149. [Google Scholar] [CrossRef]
- Breidenstein, E.B.; de la Fuente-Núñez, C.; Hancock, R.E. Pseudomonas aeruginosa: All roads lead to resistance. Trends Microbiol. 2011, 19, 419–426. [Google Scholar] [CrossRef]
- De Sousa, T.; Hébraud, M.; Dapkevicius, M.; Maltez, L.; Pereira, J.E.; Capita, R.; Alonso-Calleja, C.; Igrejas, G.; Poeta, P. Genomic and Metabolic Characteristics of the Pathogenicity in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2021, 22, 12892. [Google Scholar]
- CDC. Prevention: Antibiotic Resistance Threats in the United States; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2013. [Google Scholar]
- Pires, D.P.; Vilas, B.D.; Sillankorva, S.; Azeredo, J. Phage Therapy: A Step Forward in the Treatment of Pseudomonas aeruginosa Infections. J. Virol. 2015, 89, 7449–7456. [Google Scholar] [CrossRef] [PubMed]
- Poole, K. Outer membranes and efflux: The path to multidrug resistance in Gram-negative bacteria. Curr. Pharm. Biotechnol. 2002, 3, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Rybtke, M.; Hultqvist, L.D.; Givskov, M.; Tolker-Nielsen, T. Pseudomonas aeruginosa Biofilm Infections: Community Structure, Antimicrobial Tolerance and Immune Response. J. Mol. Biol. 2015, 427, 3628–3645. [Google Scholar] [CrossRef] [PubMed]
- Bjarnsholt, T.; Kirketerp-Møller, K.; Jensen, P.; Madsen, K.G.; Phipps, R.; Krogfelt, K.; Høiby, N.; Givskov, M. Why chronic wounds will not heal: A novel hypothesis. Wound Repair Regen. 2008, 16, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Jardak, M.; Atoissi, A.; Msalbi, D.; Atoui, D.; Bouizgarne, B.; Rigane, G.; Ben, S.R.; Aifa, S.; Mnif, S. Antibacterial, antibiofilm and cytotoxic properties of prodigiosin produced by a newly isolated Serratia sp. C6LB from a milk collection center. Microb. Pathog. 2022, 164, 105449. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, J.; Wang, X.; Kong, D.; Du, W.; Li, H.; Hse, C.Y.; Shupe, T.; Zhou, D.; Zhao, K. Biological Potential and Mechanism of Prodigiosin from Serratia marcescens Subsp. lawsoniana in Human Choriocarcinoma and Prostate Cancer Cell Lines. Int. J. Mol. Sci. 2018, 19, 3465. [Google Scholar] [CrossRef] [PubMed]
- Kimyon, Ö.; Das, T.; Ibugo, A.I.; Kutty, S.K.; Ho, K.K.; Tebben, J.; Kumar, N.; Manefield, M. Serratia Secondary Metabolite Prodigiosin Inhibits Pseudomonas aeruginosa Biofilm Development by Producing Reactive Oxygen Species that Damage Biological Molecules. Front. Microbiol. 2016, 7, 972. [Google Scholar] [CrossRef]
- Yin, Q.; Liang, J.; Zhang, W.; Zhang, L.; Hu, Z.L.; Zhang, Y.; Xu, Y. Butenolide, a Marine-Derived Broad-Spectrum Antibiofilm Agent against Both Gram-Positive and Gram-Negative Pathogenic Bacteria. Mar. Biotechnol. 2019, 21, 88–98. [Google Scholar] [CrossRef]
- Sidrim, J.J.; Vasconcelos, D.C.; Riello, G.B.; Guedes, G.M.; Serpa, R.; Bandeira, T.J.; Monteiro, A.J.; Cordeiro, R.A.; Castelo-Branco, D.S.; Rocha, M.F.; et al. Promethazine improves antibiotic efficacy and disrupts biofilms of Burkholderia pseudomallei. Biofouling 2017, 33, 88–97. [Google Scholar] [CrossRef]
- Rama Devi, K.; Srinivasan, R.; Kannappan, A.; Santhakumari, S.; Bhuvaneswari, M.; Rajasekar, P.; Prabhu, N.M.; Veera Ravi, A. In vitro and in vivo efficacy of rosmarinic acid on quorum sensing mediated biofilm formation and virulence factor production in Aeromonas hydrophila. Biofouling 2016, 32, 1171–1183. [Google Scholar]
- Suryawanshi, R.K.; Patil, C.D.; Koli, S.H.; Hallsworth, J.E.; Patil, S.V. Antimicrobial activity of prodigiosin is attributable to plasma-membrane damage. Nat. Prod. Res. 2017, 31, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W. Introduction to biofilm. Int. J. Antimicrob. Agents 1999, 11, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Govan, J.R.; Deretic, V. Microbial pathogenesis in cystic fibrosis: Mucoid Pseudomonas aeruginosa and Burkholderia cepacia. Microbiol. Rev. 1996, 60, 539–574. [Google Scholar] [CrossRef] [PubMed]
- Friedman, L.; Kolter, R. Genes involved in matrix formation in Pseudomonas aeruginosa PA14 biofilms. Mol. Microbiol. 2004, 51, 675–690. [Google Scholar] [CrossRef] [PubMed]
- Friedman, L.; Kolter, R. Two genetic loci produce distinct carbohydrate-rich structural components of the Pseudomonas aeruginosa biofilm matrix. J. Bacteriol. 2004, 186, 4457–4465. [Google Scholar] [CrossRef] [PubMed]
- Doggett, R.G. Incidence of mucoid Pseudomonas aeruginosa from clinical sources. Appl. Microbiol. 1969, 18, 936–937. [Google Scholar] [CrossRef] [PubMed]
- Hentzer, M.; Teitzel, G.M.; Balzer, G.J.; Heydorn, A.; Molin, S.; Givskov, M.; Parsek, M.R. Alginate overproduction affects Pseudomonas aeruginosa biofilm structure and function. J. Bacteriol. 2001, 183, 5395–5401. [Google Scholar] [CrossRef] [PubMed]
- Colvin, K.M.; Gordon, V.D.; Murakami, K.; Borlee, B.R.; Wozniak, D.J.; Wong, G.C.; Parsek, M.R. The pel polysaccharide can serve a structural and protective role in the biofilm matrix of Pseudomonas aeruginosa. PLoS Pathog. 2011, 7, e1001264. [Google Scholar] [CrossRef]
- Schurr, M.J. Which bacterial biofilm exopolysaccharide is preferred, Psl or alginate? J. Bacteriol. 2013, 195, 1623–1626. [Google Scholar] [CrossRef]
- Orgad, O.; Oren, Y.; Walker, S.L.; Herzberg, M. The role of alginate in Pseudomonas aeruginosa EPS adherence, viscoelastic properties and cell attachment. Biofouling 2011, 27, 787–798. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Name | Type | Primer Sequences (5′ to 3′) |
|---|---|---|
| proC | Fw | CAGGCCGGGCAGTTGCTGTC |
| Rev | GGTCAGGCGCGAGGCTGTCT | |
| algA | Fw | AGAACTGAAGAAGCACGACG |
| Rev | TTCTCCATCACCGCGTAGT | |
| pelA | Fw | ATGGCTGAAGGTATGGCTG |
| Rev | AGGTGCTGGAGGACTTCATC | |
| pslM | Fw | CTATGACGCACGGCAACTGG |
| Rev | CGCCATTGACCAGGTGCAT |
| Target Name | Type | Primer Sequences (5′ to 3′) |
|---|---|---|
| GADPH | Fw | GGACTTACAGAGGTCCGCTT |
| Rev | CTATAGGGCCTGGGTCAGTG | |
| IL-1β | Fw | CAACCAACAAGTGATATTCTCCATG |
| Rev | GATCCACACTCTCCAGCTGCA | |
| IL-6 | Fw | GAGGATACCACTCCCAACAGACC |
| Rev | AAGTGCATCATCGTTGTTCATACA | |
| IL-10 | Fw | CGAGATGCCTTCAGCAGAG |
| Rev | CGCCTTGATGTCTGGGTCTT | |
| TNF-α | Fw | CATCTTCTCAAAATTCGAGTGACAA |
| Rev | TGGGAGTAGACAAGGTACAACCC |
| Strains | MICPG (μg/mL) | MBCPG (μg/mL) | MICAZT (μg/mL) | MBCAZT (μg/mL) |
|---|---|---|---|---|
| PAO1 | 32 | 128 | - | - |
| CQMU105 | 64 | 128 | 128 | 256 |
| CQMU184 | 16 | 32 | 256 | >256 |
| CQMU293 | 8 | 32 | 32 | 128 |
| CQMU308 | 8 | 32 | 64 | >128 |
| CQMU359 | 8 | 32 | 32 | 64 |
| CQMU392 | 16 | 32 | 128 | 256 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Z.; Xiao, H.; Li, H.; Lu, X.; Yan, J.; Nie, H.; Yin, Q. Prodigiosin as an Antibiofilm Agent against the Bacterial Biofilm-Associated Infection of Pseudomonas aeruginosa. Pathogens 2024, 13, 145. https://doi.org/10.3390/pathogens13020145
Ma Z, Xiao H, Li H, Lu X, Yan J, Nie H, Yin Q. Prodigiosin as an Antibiofilm Agent against the Bacterial Biofilm-Associated Infection of Pseudomonas aeruginosa. Pathogens. 2024; 13(2):145. https://doi.org/10.3390/pathogens13020145
Chicago/Turabian StyleMa, Zhiwen, Hong Xiao, Hailin Li, Xiaoling Lu, Jing Yan, Hao Nie, and Qi Yin. 2024. "Prodigiosin as an Antibiofilm Agent against the Bacterial Biofilm-Associated Infection of Pseudomonas aeruginosa" Pathogens 13, no. 2: 145. https://doi.org/10.3390/pathogens13020145
APA StyleMa, Z., Xiao, H., Li, H., Lu, X., Yan, J., Nie, H., & Yin, Q. (2024). Prodigiosin as an Antibiofilm Agent against the Bacterial Biofilm-Associated Infection of Pseudomonas aeruginosa. Pathogens, 13(2), 145. https://doi.org/10.3390/pathogens13020145