


Clinical Strains of Mycobacterium tuberculosis Representing Different Genotype Families Exhibit Distinct Propensities to Adopt the Differentially Culturable State
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
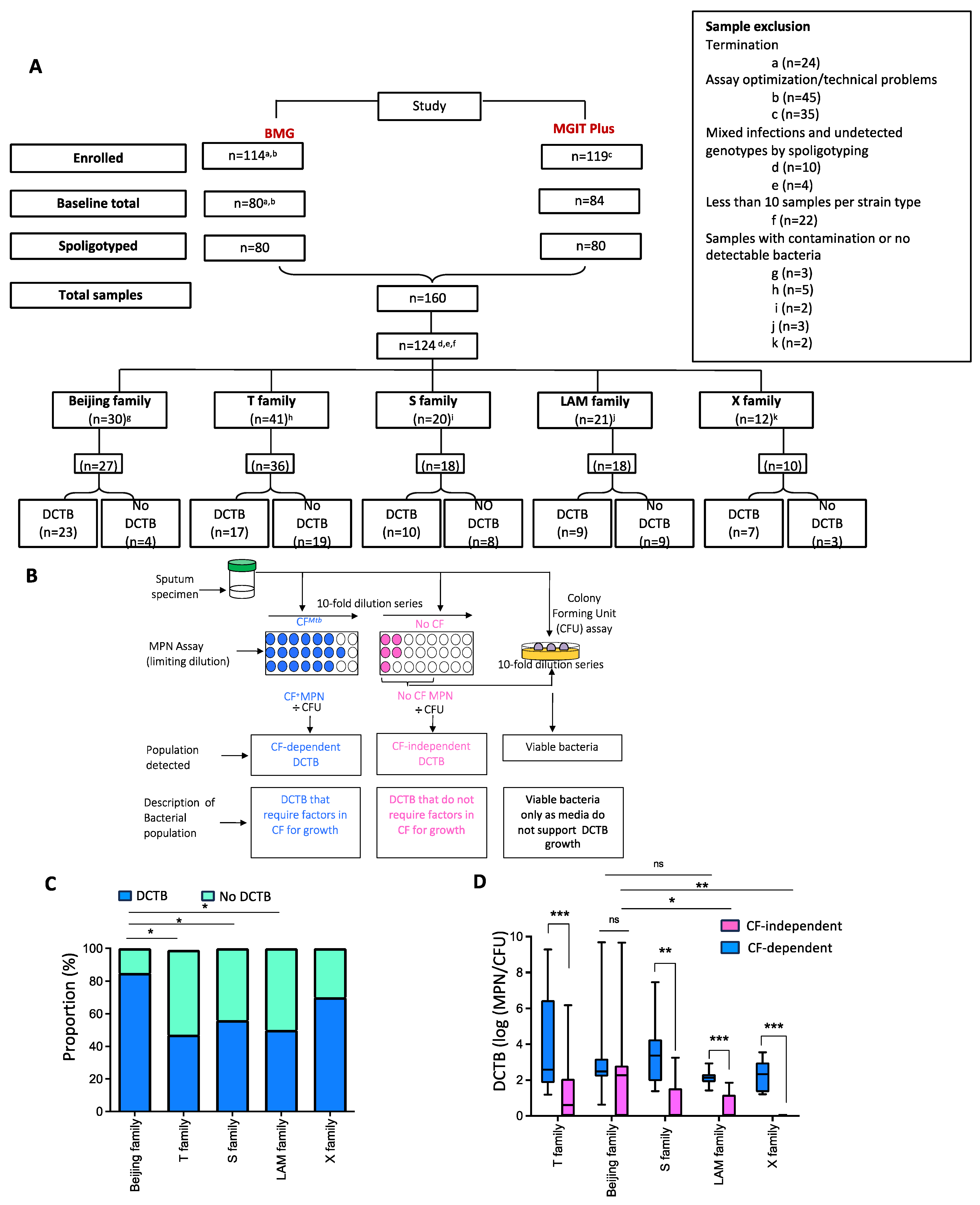
2.1. Assessment of DCTB in Clinical Cohorts
2.2. Spoligotyping of Mycobacterium tuberculosis Strains
2.3. Mycobacterial Culture Conditions
2.4. Growth Kinetics of Mycobacterial Strains
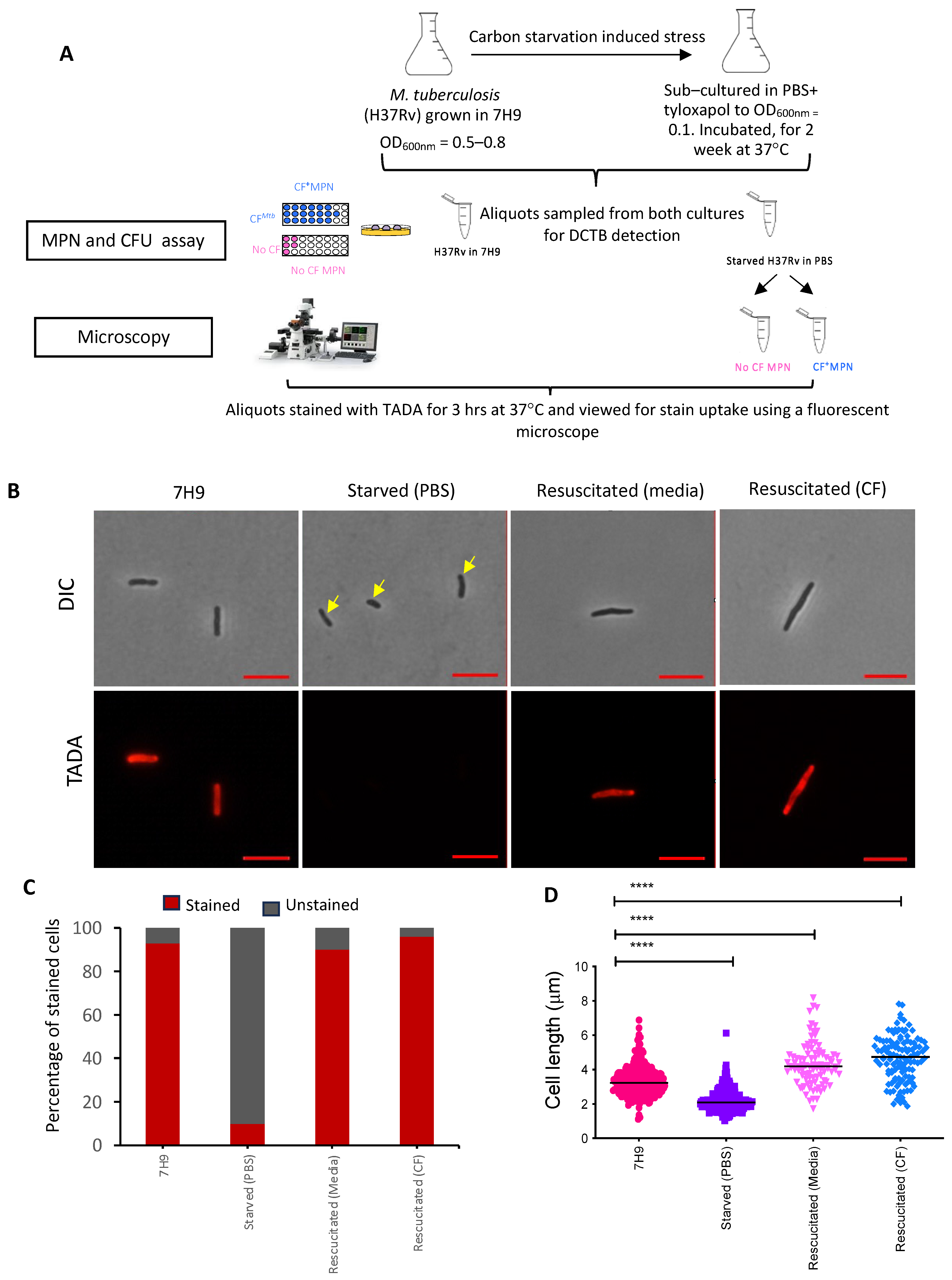
2.5. In Vitro Carbon Starvation Model for the Generation of Viable but Non-Culturable M. tuberculosis
2.6. Assessment of Starved Cells for Differentially Culturable Tubercle Bacteria (DCTB)
2.6.1. Preparation of Culture Filtrate
2.6.2. The MPN Assay
2.6.3. Determination of Colony Forming Units (CFUs)
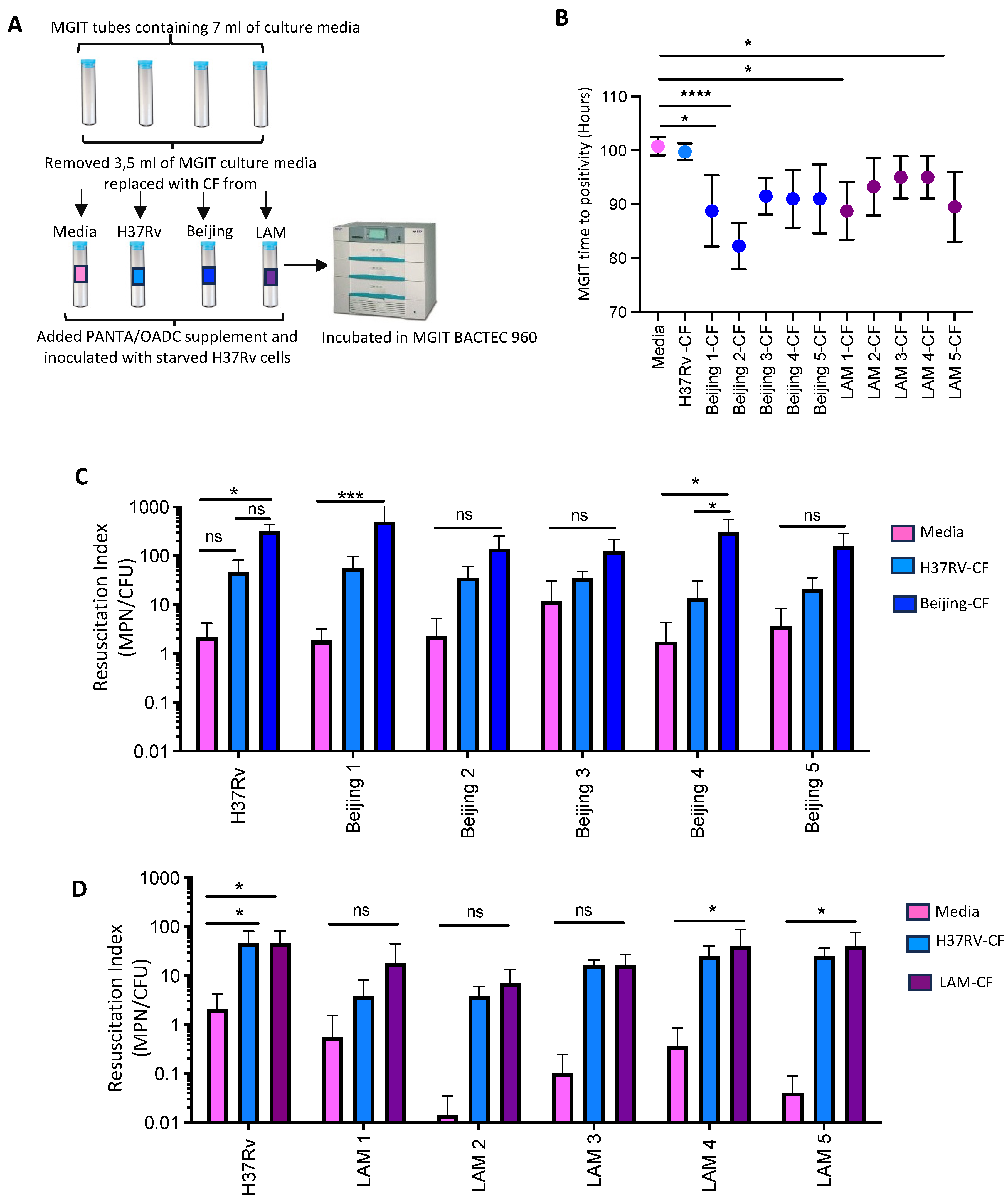
2.7. Resuscitation of Starved H37Rv, Beijing and LAM Cells in MGIT Supplemented with CF
2.8. Metabolic Labelling of Axenic, Starved and Resuscitated Cells
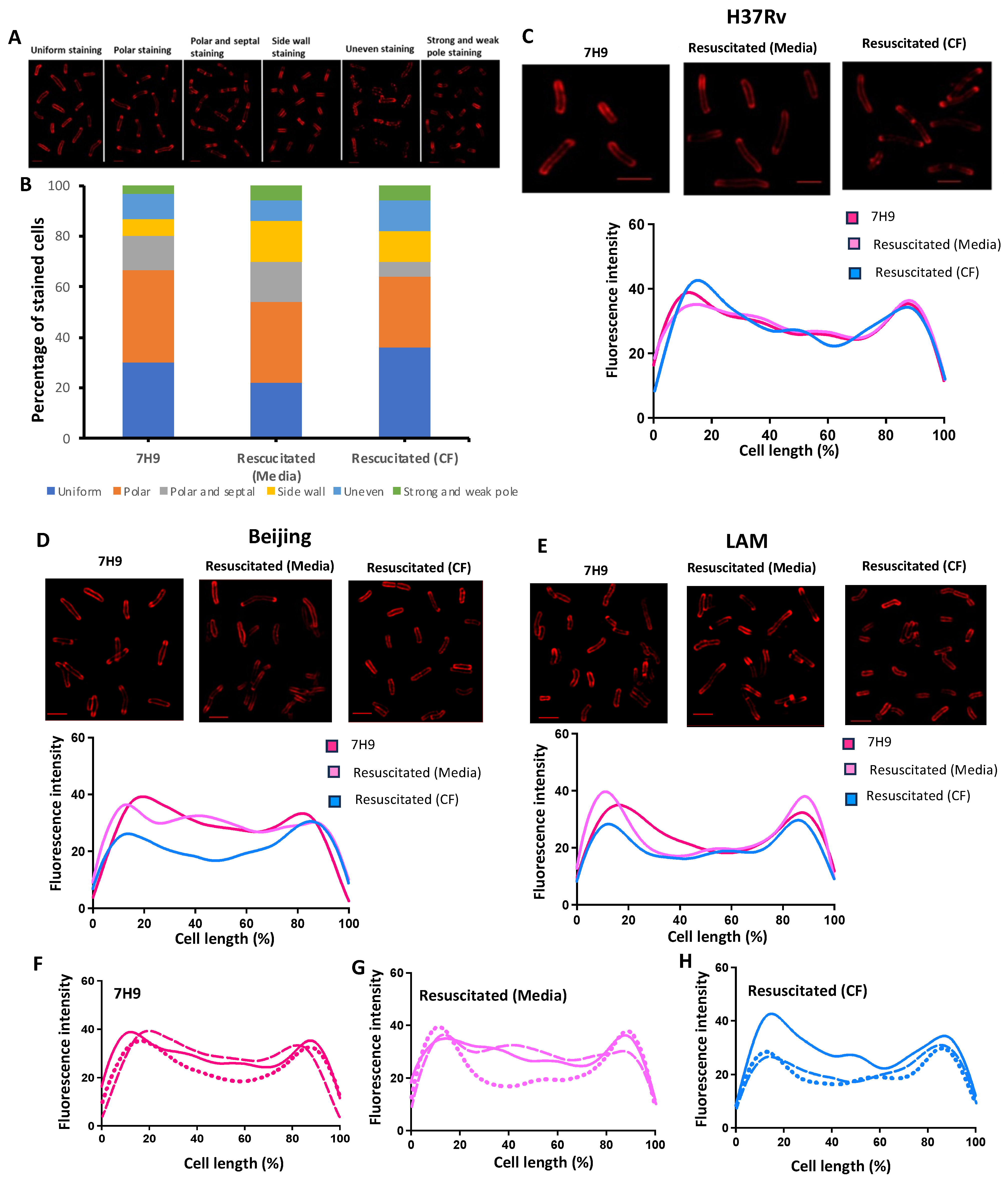
2.9. Cytological Assessment of Strained Bacteria by Microscopy
2.10. Fluorescent Staining Patterns
2.11. Cell Length Measurements
2.12. Data Analysis
3. Results
3.1. Quantification of DCTB in Sputum of Participants with Drug-Sensitive TB
3.2. Generation of DCTB Using an In Vitro Carbon Starvation Model
3.3. The Effects of Cognate CFs Derived from Beijing and LAM Strains in the Resuscitation of DCTB
3.4. Cytological Profiling of Axenic, Starved and Resuscitated Cells by Fluorescent Microscopy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chengalroyen, M.D.; Beukes, G.M.; Gordhan, B.G.; Streicher, E.M.; Churchyard, G.; Hafner, R.; Warren, R.; Otwombe, K.; Martinson, N.; Kana, B.D. Detection and quantification of differentially culturable tubercle bacteria in sputum from patients with tuberculosis. Am. J. Respir. Crit. Care Med. 2016, 194, 1532–1540. [Google Scholar] [CrossRef]
- Mukamolova, G.V.; Turapov, O.; Malkin, J.; Woltmann, G.; Barer, M.R. Resuscitation-promoting factors reveal an occult population of tubercle bacilli in sputum. Am. J. Respir. Crit. Care Med. 2010, 181, 174–180. [Google Scholar] [CrossRef]
- Gordhan, B.G.; Peters, J.S.; McIvor, A.; Machowski, E.E.; Ealand, C.; Waja, Z.; Martinson, N.; Kana, B.D. Detection of differentially culturable tubercle bacteria in sputum using mycobacterial culture filtrates. Sci. Rep. 2021, 11, 6493. [Google Scholar] [CrossRef]
- Gordhan, B.G.; Sewcharran, A.; Letsoalo, M.; Chinappa, T.; Yende-Zuma, N.; Padayatchi, N.; Naidoo, K.; Kana, B.D. Detection of differentially culturable tubercle bacteria in sputum from drug-resistant tuberculosis patients. Front. Cell. Infect. Microbiol. 2022, 12, 949370. [Google Scholar] [CrossRef]
- Peters, J.S.; McIvor, A.; Papadopoulos, A.O.; Masangana, T.; Gordhan, B.G.; Waja, Z.; Otwombe, K.; Letutu, M.; Kamariza, M.; Sterling, T.R.; et al. Differentially culturable tubercle bacteria as a measure of tuberculosis treatment response. Front. Cell. Infect. Microbiol. 2022, 12, 1064148. [Google Scholar] [CrossRef]
- Kana, B.D.; Mizrahi, V. Resuscitation-promoting factors as lytic enzymes for bacterial growth and signaling. FEMS Immunol. Med. Microbiol. 2010, 58, 39–50. [Google Scholar] [CrossRef]
- Gordhan, B.G.; Peters, J.; Kana, B.D. Application of model systems to study adaptive responses of Mycobacterium tuberculosis during infection and disease. Adv. Appl. Microbiol. 2019, 108, 115–161. [Google Scholar]
- Mukamolova, G.V.; Turapov, O.A.; Young, D.I.; Kaprelyants, A.S.; Kell, D.B.; Young, M. A family of autocrine growth factors in Mycobacterium tuberculosis. Mol. Microbiol. 2002, 46, 623–635. [Google Scholar] [CrossRef]
- Bamford, R.A.; Smith, A.; Metz, J.; Glover, G.; Titball, R.W.; Pagliara, S. Investigating the physiology of viable but non-culturable bacteria by microfluidics and time-lapse microscopy. BMC Biol. 2017, 15, 121. [Google Scholar] [CrossRef]
- Dartois, V. The path of anti-tuberculosis drugs: From blood to lesions to mycobacterial cells. Nat. Rev. Microbiol. 2014, 12, 159–167. [Google Scholar] [CrossRef]
- Dusthackeer, A.; Balasubramanian, M.; Shanmugam, G.; Priya, S.; Nirmal, C.R.; Sam Ebenezer, R.; Balasubramanian, A.; Mondal, R.K.; Thiruvenkadam, K.; Hemanth Kumar, A.K.; et al. Differential culturability of Mycobacterium tuberculosis in culture-negative sputum of patients with pulmonary tuberculosis and in a simulated model of dormancy. Front. Microbiol. 2019, 10, 2381. [Google Scholar] [CrossRef]
- Oliver, J.D. Recent findings on the viable but nonculturable state in pathogenic bacteria. FEMS Microbiol. Rev. 2010, 34, 415–425. [Google Scholar] [CrossRef]
- Gold, B.; Nathan, C. Targeting Phenotypically Tolerant Mycobacterium tuberculosis. Mycrobiology Spectr. 2017, 5, 1–69. [Google Scholar]
- Turapov, O.; O’Connor, B.D.; Sarybaeva, A.A.; Williams, C.; Patel, H.; Kadyrov, A.S.; Sarybaev, A.S.; Woltmann, G.; Barer, M.R.; Mukamolova, G.V. Phenotypically Adapted Mycobacterium tuberculosis Populations from Sputum Are Tolerant to First-Line Drugs. Antimicrob. Agents Chemother. 2016, 60, 2476–2483. [Google Scholar] [CrossRef]
- Betts, J.C.; Lukey, P.T.; Robb, L.C.; McAdam, R.A.; Duncan, K. Evaluation of a nutrient starvation model of Mycobacterium tuberculosis persistence by gene and protein expression profiling. Mol. Microbiol. 2002, 43, 717–731. [Google Scholar] [CrossRef]
- Kulka, K.; Hatfull, G.; Ojha, A.K. Growth of Mycobacterium tuberculosis biofilms. J. Vis. Exp. JoVE 2012, 60, e3820. [Google Scholar]
- Loebel, R.O.; Shorr, E.; Richardson, H.B. The influence of adverse conditions upon the respiratory metabolism and growth of human tubercle bacilli. J. Bacteriol. 1933, 26, 167–200. [Google Scholar] [CrossRef]
- Ojha, A.K.; Baughn, A.D.; Sambandan, D.; Hsu, T.; Trivelli, X.; Guerardel, Y.; Alahari, A.; Kremer, L.; Jacobs, W.R., Jr.; Hatfull, G.F. Growth of Mycobacterium tuberculosis biofilms containing free mycolic acids and harbouring drug-tolerant bacteria. Mol. Microbiol. 2008, 69, 164–174. [Google Scholar] [CrossRef]
- Salina, E.G.; Waddell, S.J.; Hoffmann, N.; Rosenkrands, I.; Butcher, P.D.; Kaprelyants, A.S. Potassium availability triggers Mycobacterium tuberculosis transition to, and resuscitation from, non-culturable (dormant) states. Open Biol. 2014, 4, 140106. [Google Scholar] [CrossRef]
- Wayne, L.G.; Hayes, L.G. An in vitro model for sequential study of shiftdown of Mycobacterium tuberculosis through two stages of nonreplicating persistence. Infect. Immun. 1996, 64, 2062–2069. [Google Scholar] [CrossRef]
- Shleeva, M.O.; Bagramyan, K.; Telkov, M.V.; Mukamolova, G.V.; Young, M.; Kell, D.B.; Kaprelyants, A.S. Formation and resuscitation of “non-culturable” cells of Rhodococcus rhodochrous and Mycobacterium tuberculosis in prolonged stationary phase. Microbiology 2002, 148, 1581–1591. [Google Scholar] [CrossRef] [PubMed]
- Voskuil, M.I.; Schnappinger, D.; Visconti, K.C.; Harrell, M.I.; Dolganov, G.M.; Sherman, D.R.; Schoolnik, G.K. Inhibition of respiration by nitric oxide induces a Mycobacterium tuberculosis dormancy program. J. Exp. Med. 2003, 198, 705–713. [Google Scholar] [CrossRef]
- Saito, K.; Warrier, T.; Somersan-Karakaya, S.; Kaminski, L.; Mi, J.; Jiang, X.; Park, S.; Shigyo, K.; Gold, B.; Roberts, J.; et al. Rifamycin action on RNA polymerase in antibiotic-tolerant Mycobacterium tuberculosis results in differentially detectable populations. Proc. Natl. Acad. Sci. USA 2017, 114, E4832–E4840. [Google Scholar] [CrossRef]
- Kamudumuli, P.S.; Beylis, N.; Blann, L.; Duse, A. Molecular typing of drug-susceptible and -resistant Mycobacterium tuberculosis in Johannesburg, South Africa. Int. J. Tuberc. Lung Dis. 2015, 19, 834–840. [Google Scholar] [CrossRef]
- Gandhi, N.R.; Moll, A.; Sturm, A.W.; Pawinski, R.; Govender, T.; Lalloo, U.; Zeller, K.; Andrews, J.; Friedland, G. Extensively drug-resistant tuberculosis as a cause of death in patients co-infected with tuberculosis and HIV in a rural area of South Africa. Lancet 2006, 368, 1575–1580. [Google Scholar] [CrossRef]
- Aguilar, D.; Hanekom, M.; Mata, D.; Gey van Pittius, N.C.; van Helden, P.D.; Warren, R.M.; Hernandez-Pando, R. Mycobacterium tuberculosis strains with the Beijing genotype demonstrate variability in virulence associated with transmission. Tuberculosis 2010, 90, 319–325. [Google Scholar] [CrossRef]
- Niemann, S.; Diel, R.; Khechinashvili, G.; Gegia, M.; Mdivani, N.; Tang, Y.W. Mycobacterium tuberculosis Beijing lineage favors the spread of multidrug-resistant tuberculosis in the Republic of Georgia. J. Clin. Microbiol. 2010, 48, 3544–3550. [Google Scholar] [CrossRef] [PubMed]
- Reed, M.B.; Gagneux, S.; Deriemer, K.; Small, P.M.; Barry, C.E., 3rd. The W-Beijing lineage of Mycobacterium tuberculosis overproduces triglycerides and has the DosR dormancy regulon constitutively upregulated. J. Bacteriol. 2007, 189, 2583–2589. [Google Scholar] [CrossRef] [PubMed]
- Kuru, E.; Hughes, H.V.; Brown, P.J.; Hall, E.; Tekkam, S.; Cava, F.; de Pedro, M.A.; Brun, Y.V.; VanNieuwenhze, M.S. In Situ probing of newly synthesized peptidoglycan in live bacteria with fluorescent D-amino acids. Angew. Chem. Int. Ed. Engl. 2012, 51, 12519–12523. [Google Scholar] [CrossRef]
- McIvor, A.; Gordhan, B.G.; Waja, Z.; Otwombe, K.; Martinson, N.A.; Kana, B.D. Supplementation of sputum cultures with culture filtrate to detect tuberculosis in a cross-sectional study of HIV-infected individuals. Tuberculosis 2021, 129, 102103. [Google Scholar] [CrossRef]
- Vitol, I.; Driscoll, J.; Kreiswirth, B.; Kurepina, N.; Bennett, K.P. Identifying Mycobacterium tuberculosis complex strain families using spoligotypes. Infect. Genet. Evol. 2006, 6, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Hanekom, M.; Van Pittius, N.G.; McEvoy, C.; Victor, T.; Van Helden, P.; Warren, R. Mycobacterium tuberculosis Beijing genotype: A template for success. Tuberculosis 2011, 91, 510–523. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Cave, M.D.; Zhang, L.; Foxman, B.; Marrs, C.F.; Bates, J.H.; Yang, Z.H. Association between Mycobacterium tuberculosis Beijing/W lineage strain infection and extrathoracic tuberculosis: Insights from epidemiologic and clinical characterization of the three principal genetic groups of M. tuberculosis clinical isolates. J. Clin. Microbiol. 2007, 45, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Parwati, I.; van Crevel, R.; van Soolingen, D. Possible underlying mechanisms for successful emergence of the Mycobacterium tuberculosis Beijing genotype strains. Lancet Infect. Dis. 2010, 10, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Gerasimova, A.; Kazakov, A.E.; Arkin, A.P.; Dubchak, I.; Gelfand, M.S. Comparative genomics of the dormancy regulons in mycobacteria. J. Bacteriol. 2011, 193, 3446–3452. [Google Scholar] [CrossRef] [PubMed]
- Cowley, D.; Govender, D.; February, B.; Wolfe, M.; Steyn, L.; Evans, J.; Wilkinson, R.J.; Nicol, M.P. Recent and rapid emergence of W-Beijing strains of Mycobacterium tuberculosis in Cape Town, South Africa. Clin. Infect. Dis. 2008, 47, 1252–1259. [Google Scholar] [CrossRef] [PubMed]
- de Jong, B.C.; Hill, P.C.; Aiken, A.; Awine, T.; Antonio, M.; Adetifa, I.M.; Jackson-Sillah, D.J.; Fox, A.; Deriemer, K.; Gagneux, S.; et al. Progression to active tuberculosis, but not transmission, varies by Mycobacterium tuberculosis lineage in The Gambia. J. Infect. Dis. 2008, 198, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Kremer, K.; van-der-Werf, M.J.; Au, B.K.; Anh, D.D.; Kam, K.M.; van-Doorn, H.R.; Borgdorff, M.W.; van-Soolingen, D. Vaccine-induced immunity circumvented by typical Mycobacterium tuberculosis Beijing strains. Emerg. Infect. Dis. 2009, 15, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Thwaites, G.; Caws, M.; Chau, T.T.; D’Sa, A.; Lan, N.T.; Huyen, M.N.; Gagneux, S.; Anh, P.T.; Tho, D.Q.; Torok, E.; et al. Relationship between Mycobacterium tuberculosis genotype and the clinical phenotype of pulmonary and meningeal tuberculosis. J. Clin. Microbiol. 2008, 46, 1363–1368. [Google Scholar] [CrossRef]
- Verrall, A.J.; Chaidir, L.; Ruesen, C.; Apriani, L.; Koesoemadinata, R.C.; van Ingen, J.; Sharples, K.; van Crevel, R.; Alisjahbana, B.; Hill, P.C.; et al. Lower Bacillus Calmette-Guerin protection against Mycobacterium tuberculosis infection after exposure to Beijing Strains. Am. J. Respir. Crit. Care Med. 2020, 201, 1152–1155. [Google Scholar] [CrossRef]
- Carey, A.F.; Wang, X.; Cicchetti, N.; Spaulding, C.N.; Liu, Q.; Hopkins, F.; Brown, J.; Sixsmith, J.; Sutiwisesak, R.; Behar, S.M.; et al. Multiplexed strain phenotyping defines consequences of genetic diversity in Mycobacterium tuberculosis for infection and vaccination outcomes. mSystems 2022, 7, e0011022. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Chowdhury, N.; Yamasaki, R.; Wood, T.K. Viable but non-culturable and persistence describe the same bacterial stress state. Environ. Microbiol. 2018, 20, 2038–2048. [Google Scholar] [CrossRef] [PubMed]
- Zainabadi, K.; Walsh, K.F.; Vilbrun, S.C.; Mathurin, L.D.; Lee, M.H.; Saito, K.; Mishra, S.; Ocheretina, O.; Pape, J.W.; Nathan, C.; et al. Characterization of differentially detectable Mycobacterium tuberculosis in the sputum of subjects with drug-sensitive or drug-resistant tuberculosis before and after two months of therapy. Antimicrob. Agents Chemother. 2021, 65, e0060821. [Google Scholar] [CrossRef] [PubMed]
- McAulay, K.; Saito, K.; Warrier, T.; Walsh, K.F.; Mathurin, L.D.; Royal-Mardi, G.; Lee, M.H.; Ocheretina, O.; Pape, J.W.; Fitzgerald, D.W.; et al. Differentially detectable Mycobacterium tuberculosis cells in sputum from treatment-naive subjects in Haiti and their proportionate increase after initiation of treatment. mBio 2018, 9, 10-1128. [Google Scholar] [CrossRef]
- Padarath, K. The Emergence of Differentially Culturable Tubercle Bacteria in Clinical Drug Sensitive Mycobacterium tuberculosis Strains. Master’s Thesis, University of the Witwatersrand, Johannesburg, South Africa, 2020. Available online: https://wiredspace.wits.ac.za/items/d39e7d9c-9e49-42ab-b5e8-9466117008a2 (accessed on 1 August 2023).
- Shleeva, M.O.; Kudykina, Y.K.; Vostroknutova, G.N.; Suzina, N.E.; Mulyukin, A.L.; Kaprelyants, A.S. Dormant ovoid cells of Mycobacterium tuberculosis are formed in response to gradual external acidification. Tuberculosis 2011, 91, 146–154. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Combined Studies | ||
|---|---|---|---|
| BMG Study (n = 61) | MGIT PLUS Study (n = 48) | BMG Study and MGIT PLUS Study (n = 109) | |
| Demographics | |||
| Sex: Female (%) Male (%) | 34 66 | 42 58 | 38 62 |
| Age, yrs, Median (IQR) | 35 (26.5–45.0) | 37 (30–44) | 36 (28.5–44.0) |
| BMI, Median (IQR) | 19.07 (17.8–22.1) | 21.5 (18.4–23.5) | 20.17 (18.0–22.8) |
| HIV Status, n (%) Negative Positive | 34 66 | 0 100 | 19 81 |
| GeneXpert cycle threshold (IQR) | 18.7 (14.3–24.4) * | 22.7 (18.7–26.6) # | 21.2 (16.3–25.2) § |
| Conventional TB Diagnosis, n (%) Smear grade negative Smear grade positive | 23 77 | 21 79 | 22 78 |
| MGIT Time to Positivity, d (IQR) | 6 (4.0–11.5) | 11.0 (7.3–15.0) | 8.0 (5.0–14.5) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gordhan, B.G.; Padarath, K.; Sewcharran, A.; McIvor, A.; VanNieuwenhze, M.S.; Waja, Z.; Martinson, N.; Kana, B.D. Clinical Strains of Mycobacterium tuberculosis Representing Different Genotype Families Exhibit Distinct Propensities to Adopt the Differentially Culturable State. Pathogens 2024, 13, 318. https://doi.org/10.3390/pathogens13040318
Gordhan BG, Padarath K, Sewcharran A, McIvor A, VanNieuwenhze MS, Waja Z, Martinson N, Kana BD. Clinical Strains of Mycobacterium tuberculosis Representing Different Genotype Families Exhibit Distinct Propensities to Adopt the Differentially Culturable State. Pathogens. 2024; 13(4):318. https://doi.org/10.3390/pathogens13040318
Chicago/Turabian StyleGordhan, Bhavna Gowan, Kiyasha Padarath, Astika Sewcharran, Amanda McIvor, Michael S. VanNieuwenhze, Ziyaad Waja, Neil Martinson, and Bavesh Davandra Kana. 2024. "Clinical Strains of Mycobacterium tuberculosis Representing Different Genotype Families Exhibit Distinct Propensities to Adopt the Differentially Culturable State" Pathogens 13, no. 4: 318. https://doi.org/10.3390/pathogens13040318
APA StyleGordhan, B. G., Padarath, K., Sewcharran, A., McIvor, A., VanNieuwenhze, M. S., Waja, Z., Martinson, N., & Kana, B. D. (2024). Clinical Strains of Mycobacterium tuberculosis Representing Different Genotype Families Exhibit Distinct Propensities to Adopt the Differentially Culturable State. Pathogens, 13(4), 318. https://doi.org/10.3390/pathogens13040318