
Therapeutic Applications of Aggregatibacter actinomycetemcomitans Leukotoxin


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Hematologic Malignancies
3. Psoriasis
4. Allergic Asthma
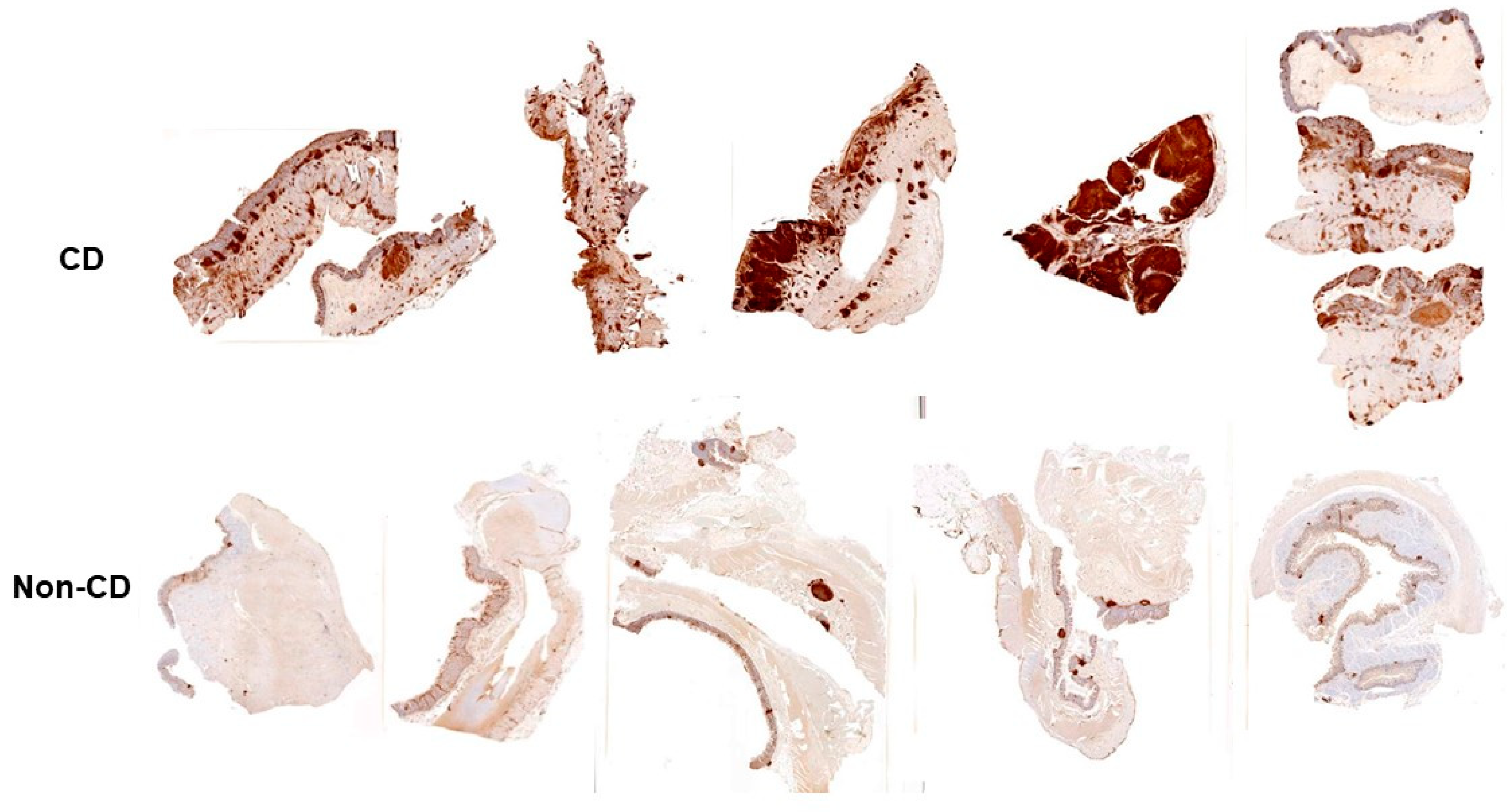
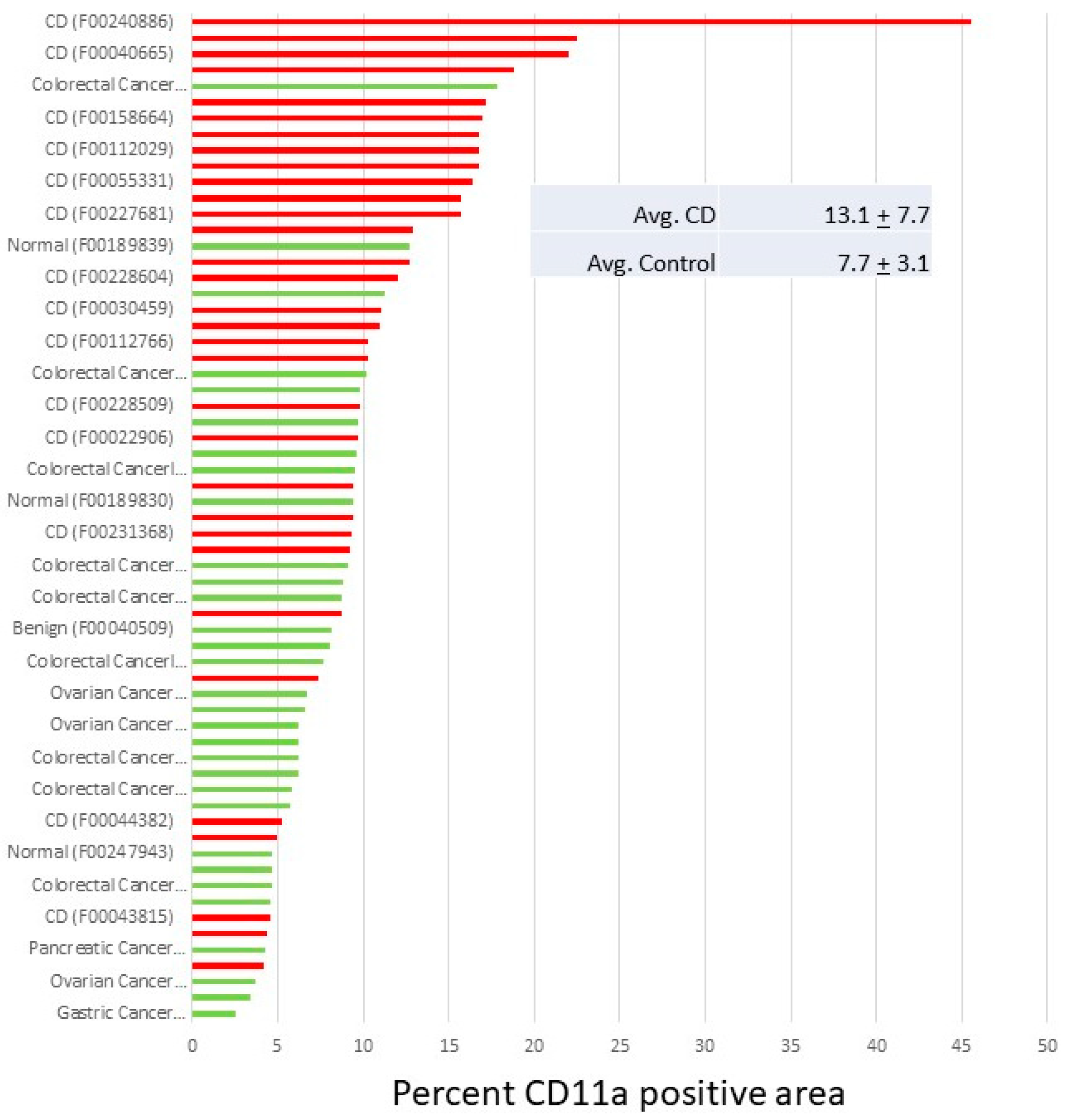
5. Inflammatory Bowel Disease
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Haubek, D.; Ennibi, O.K.; Poulsen, K.; Vaeth, M.; Poulsen, S.; Kilian, M. Risk of aggressive periodontitis in adolescent carriers of the JP2 clone of Aggregatibacter (Actinobacillus) actinomycetemcomitans in Morocco: A prospective longitudinal cohort study. Lancet 2008, 371, 237–242. [Google Scholar] [CrossRef]
- Lally, E.T.; Kieba, I.R.; Demuth, D.R.; Rosenbloom, J.; Golub, E.E.; Taichman, N.S.; Gibson, C.W. Identification and expression of the Actinobacillus actinomycetemcomitans leukotoxin gene. Biochem. Biophys. Res. Commun. 1989, 159, 256–262. [Google Scholar] [CrossRef]
- Lally, E.T.; Kieba, I.R.; Sato, A.; Green, C.L.; Rosenbloom, J.; Korostoff, J.; Wang, J.F.; Shenker, B.J.; Ortlepp, S.; Robinson, M.K.; et al. RTX toxins recognize a beta2 integrin on the surface of human target cells. J. Biol. Chem. 1997, 272, 30463–30469. [Google Scholar] [CrossRef]
- Evans, R.; Patzak, I.; Svensson, L.; De Filippo, K.; Jones, K.; McDowall, A.; Hogg, N. Integrins in immunity. J. Cell Sci. 2009, 122, 215–225. [Google Scholar] [CrossRef]
- Hogg, N.; Harvey, J.; Cabanas, C.; Landis, R.C. Control of leukocyte integrin activation. Am. Rev. Respir. Dis. 1993, 148, S55–S59. [Google Scholar] [CrossRef]
- Smith, A.; Stanley, P.; Jones, K.; Svensson, L.; McDowall, A.; Hogg, N. The role of the integrin LFA-1 in T-lymphocyte migration. Immunol. Rev. 2007, 218, 135–146. [Google Scholar] [CrossRef]
- Tsai, C.C.; McArthur, W.P.; Baehni, P.C.; Hammond, B.F.; Taichman, N.S. Extraction and partial characterization of a leukotoxin from a plaque-derived Gram-negative microorganism. Infect. Immun. 1979, 25, 427–439. [Google Scholar] [CrossRef]
- Tsai, C.C.; Shenker, B.J.; DiRienzo, J.M.; Malamud, D.; Taichman, N.S. Extraction and isolation of a leukotoxin from Actinobacillus actinomycetemcomitans with polymyxin B. Infect. Immun. 1984, 43, 700–705. [Google Scholar] [CrossRef]
- DiRienzo, J.M.; Tsai, C.C.; Shenker, B.J.; Taichman, N.S.; Lally, E.T. Monoclonal antibodies to leukotoxin of Actinobacillus actinomycetemcomitans. Infect. Immun. 1985, 47, 31–36. [Google Scholar] [CrossRef]
- Lally, E.T.; Golub, E.E.; Kieba, I.R.; Taichman, N.S.; Decker, S.; Berthold, P.; Gibson, C.W.; Demuth, D.R.; Rosenbloom, J. Structure and function of the B and D genes of the Actinobacillus actinomycetemcomitans leukotoxin complex. Microb. Pathog. 1991, 11, 111–121. [Google Scholar] [CrossRef]
- Kachlany, S.C.; Fine, D.H.; Figurski, D.H. Secretion of RTX leukotoxin by Actinobacillus actinomycetemcomitans. Infect. Immun. 2000, 68, 6094–6100. [Google Scholar] [CrossRef]
- Kachlany, S.C.; Fine, D.H.; Figurski, D.H. Purification of secreted leukotoxin (LtxA) from Actinobacillus actinomycetemcomitans. Protein Expr. Purif. 2002, 25, 465–471. [Google Scholar] [CrossRef]
- Berthold, P.; Forti, D.; Kieba, I.R.; Rosenbloom, J.; Taichman, N.S.; Lally, E.T. Electron immunocytochemical localization of Actinobacillus actinomycetemcomitans leukotoxin. Oral Microbiol. Immunol. 1992, 7, 24–27. [Google Scholar] [CrossRef]
- Kato, S.; Kowashi, Y.; Demuth, D.R. Outer membrane-like vesicles secreted by Actinobacillus actinomycetemcomitans are enriched in leukotoxin. Microb. Pathog. 2002, 32, 1–13. [Google Scholar] [CrossRef]
- Ohta, H.; Kato, K.; Kokeguchi, S.; Hara, H.; Fukui, K.; Murayama, Y. Nuclease-sensitive binding of an Actinobacillus actinomycetemcomitans leukotoxin to the bacterial cell surface. Infect. Immun. 1991, 59, 4599–4605. [Google Scholar] [CrossRef]
- Hogg, N.; Smith, A.; McDowall, A.; Giles, K.; Stanley, P.; Laschinger, M.; Henderson, R. How T cells use LFA-1 to attach and migrate. Immunol. Lett. 2004, 92, 51–54. [Google Scholar] [CrossRef]
- Giblin, P.A.; Lemieux, R.M. LFA-1 as a key regulator of immune function: Approaches toward the development of LFA-1-based therapeutics. Curr. Pharm. Des. 2006, 12, 2771–2795. [Google Scholar] [CrossRef]
- Ma, Q.; Shimaoka, M.; Lu, C.; Jing, H.; Carman, C.V.; Springer, T.A. Activation-induced conformational changes in the I domain region of lymphocyte function-associated antigen 1. J. Biol. Chem. 2002, 277, 10638–10641. [Google Scholar] [CrossRef]
- Dransfield, I.; Cabanas, C.; Barrett, J.; Hogg, N. Interaction of leukocyte integrins with ligand is necessary but not sufficient for function. J. Cell Biol. 1992, 116, 1527–1535. [Google Scholar] [CrossRef] [PubMed]
- Dransfield, I.; Cabanas, C.; Craig, A.; Hogg, N. Divalent cation regulation of the function of the leukocyte integrin LFA-1. J. Cell Biol. 1992, 116, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Porter, J.C.; Bracke, M.; Smith, A.; Davies, D.; Hogg, N. Signaling through integrin LFA-1 leads to filamentous actin polymerization and remodeling, resulting in enhanced T cell adhesion. J. Immunol. 2002, 168, 6330–6335. [Google Scholar] [CrossRef] [PubMed]
- Hioe, C.E.; Tuen, M.; Vasiliver-Shamis, G.; Alvarez, Y.; Prins, K.C.; Banerjee, S.; Nadas, A.; Cho, M.W.; Dustin, M.L.; Kachlany, S.C. HIV envelope gp120 activates LFA-1 on CD4 T-lymphocytes and increases cell susceptibility to LFA-1-targeting leukotoxin (LtxA). PLoS ONE 2011, 6, e23202. [Google Scholar] [CrossRef] [PubMed]
- Kachlany, S.C.; Schwartz, A.B.; Balashova, N.V.; Hioe, C.E.; Tuen, M.; Le, A.; Kaur, M.; Mei, Y.; Rao, J. Anti-leukemia activity of a bacterial toxin with natural specificity for LFA-1 on white blood cells. Leuk. Res. 2010, 34, 777–785. [Google Scholar] [CrossRef] [PubMed]
- DiFranco, K.M.; Gupta, A.; Galusha, L.E.; Perez, J.; Nguyen, T.V.; Fineza, C.D.; Kachlany, S.C. Leukotoxin (Leukothera(R)) targets active leukocyte function antigen-1 (LFA-1) protein and triggers a lysosomal mediated cell death pathway. J. Biol. Chem. 2012, 287, 17618–17627. [Google Scholar] [CrossRef] [PubMed]
- DiFranco, K.M.; Kaswala, R.H.; Patel, C.; Kasinathan, C.; Kachlany, S.C. Leukotoxin kills rodent WBC by targeting leukocyte function associated antigen 1. Comp. Med. 2013, 63, 331–337. [Google Scholar] [PubMed]
- Kaur, M.; Kachlany, S.C. Aggregatibacter actinomycetemcomitans leukotoxin (LtxA; Leukothera) induces cofilin dephosphorylation and actin depolymerization during killing of malignant monocytes. Microbiology 2014, 160, 2443–2452. [Google Scholar] [CrossRef] [PubMed]
- DiFranco, K.M.; Johnson-Farley, N.; Bertino, J.R.; Elson, D.; Vega, B.A.; Belinka, B.A., Jr.; Kachlany, S.C. LFA-1-targeting Leukotoxin (LtxA; Leukothera(R)) causes lymphoma tumor regression in a humanized mouse model and requires caspase-8 and Fas to kill malignant lymphocytes. Leuk. Res. 2015, 39, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Vega, B.A.; Schober, L.T.; Kim, T.; Belinka, B.A., Jr.; Kachlany, S.C. Aggregatibacter actinomycetemcomitans leukotoxin (LtxA; Leukothera(R)) requires death receptor Fas, in addition to LFA-1, to trigger cell death in T lymphocytes. Infect. Immun. 2019, 87, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Prince, D.J.; Patel, D.; Kachlany, S.C. Leukotoxin (LtxA/Leukothera) induces ATP expulsion via pannexin-1 channels and subsequent cell death in malignant lymphocytes. Sci. Rep. 2021, 11, 18086. [Google Scholar] [CrossRef]
- Hasegawa, Y.; Yokono, K.; Taki, T.; Amano, K.; Tominaga, Y.; Yoneda, R.; Yagi, N.; Maeda, S.; Yagita, H.; Okumura, K.; et al. Prevention of autoimmune insulin-dependent diabetes in non-obese diabetic mice by anti-LFA-1 and anti-ICAM-1 mAb. Int. Immunol. 1994, 6, 831–838. [Google Scholar] [CrossRef]
- McMurray, R.W. Adhesion molecules in autoimmune disease. Semin. Arthritis Rheum. 1996, 25, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Yokomori, H.; Oda, M.; Yoshimura, K.; Nomura, M.; Ogi, M.; Wakabayashi, G.; Kitajima, M.; Ishii, H. Expression of intercellular adhesion molecule-1 and lymphocyte function-associated antigen-1 protein and messenger RNA in primary biliary cirrhosis. Intern. Med. 2003, 42, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Connolly, M.K.; Kitchens, E.A.; Chan, B.; Jardieu, P.; Wofsy, D. Treatment of murine lupus with monoclonal antibodies to lymphocyte function-associated antigen-1: Dose-dependent inhibition of autoantibody production and blockade of the immune response to therapy. Clin. Immunol. Immunopathol. 1994, 72, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Elovaara, I.; Lalla, M.; Spare, E.; Lehtimaki, T.; Dastidar, P. Methylprednisolone reduces adhesion molecules in blood and cerebrospinal fluid in patients with MS. Neurology 1998, 51, 1703–1708. [Google Scholar] [CrossRef]
- Engelhardt, B. Molecular mechanisms involved in T cell migration across the blood-brain barrier. J. Neural Transm. 2006, 113, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Thomson, A.W.; Satoh, S.; Nussler, A.K.; Tamura, K.; Woo, J.; Gavaler, J.; van Thiel, D.H. Circulating intercellular adhesion molecule-1 (ICAM-1) in autoimmune liver disease and evidence for the production of ICAM-1 by cytokine-stimulated human hepatocytes. Clin. Exp. Immunol. 1994, 95, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Yusuf-Makagiansar, H.; Anderson, M.E.; Yakovleva, T.V.; Murray, J.S.; Siahaan, T.J. Inhibition of LFA-1/ICAM-1 and VLA-4/VCAM-1 as a therapeutic approach to inflammation and autoimmune diseases. Med. Res. Rev. 2002, 22, 146–167. [Google Scholar] [CrossRef]
- Aureli, A.; Marziani, B.; Venditti, A.; Sconocchia, T.; Sconocchia, G. Acute Lymphoblastic Leukemia Immunotherapy Treatment: Now, Next, and Beyond. Cancers 2023, 15, 3346. [Google Scholar] [CrossRef]
- Eisfeld, A.K. Disparities in acute myeloid leukemia treatments and outcomes. Curr. Opin. Hematol. 2024, 31, 58–63. [Google Scholar] [CrossRef]
- Mikhael, J.; Cichewicz, A.; Mearns, E.S.; Girvan, A.; Pierre, V.; Rawashdh, N.A.; Yellow-Duke, A.; Cornell, R.F.; Nixon, M. Overall Survival in Patients with Multiple Myeloma in the U.S.: A Systematic Literature Review of Racial Disparities. Clin. Lymphoma Myeloma Leuk. 2024, 24, e1–e12. [Google Scholar] [CrossRef]
- Schimmoeller, C.J.; Bastian, C.; Fleming, J.; Morales, J. A Review of Hodgkin Lymphoma in the Era of Checkpoint Inhibitors. Cureus 2023, 15, e41660. [Google Scholar] [CrossRef] [PubMed]
- Afvari, S.; Beck, T.C.; Kazlouskaya, M.; Afrahim, R.; Valdebran, M. Diet, sleep, and exercise in inflammatory skin diseases. Our Dermatol. Online 2023, 14, 430–435. [Google Scholar] [CrossRef]
- Bhagwat, A.P.; Madke, B. The Current Advancement in Psoriasis. Cureus 2023, 15, e47006. [Google Scholar] [CrossRef]
- Lie, E.; Choi, M.; Wang, S.P.; Eichenfield, L.F. Topical Management of Pediatric Psoriasis: A Review of New Developments and Existing Therapies. Paediatr. Drugs 2024, 26, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Reali, E.; Ferrari, D. From the Skin to Distant Sites: T Cells in Psoriatic Disease. Int. J. Mol. Sci. 2023, 24, 15707. [Google Scholar] [CrossRef]
- Stenderup, K.; Rosada, C.; Dam, T.N.; Salerno, E.; Belinka, B.A.; Kachlany, S.C. Resolution of psoriasis by a leukocyte-targeting bacterial protein in a humanized mouse model. J. Investig. Dermatol. 2011, 131, 2033–2039. [Google Scholar] [CrossRef] [PubMed]
- Dunn, J.L.M.; Rothenberg, M.E. 2021 year in review: Spotlight on eosinophils. J. Allergy Clin. Immunol. 2022, 149, 517–524. [Google Scholar] [CrossRef]
- Kim, Y.M.; Kim, Y.S.; Jeon, S.G.; Kim, Y.K. Immunopathogenesis of allergic asthma: More than the th2 hypothesis. Allergy Asthma Immunol. Res. 2013, 5, 189–196. [Google Scholar] [CrossRef]
- Matos-Semedo, F.; Cruz, C.; Inacio, F.; Gama, J.M.R.; Nwaru, B.I.; Taborda-Barata, L. House dust mite (HDM) and storage mite (SM) molecular sensitisation profiles and association with clinical outcomes in allergic asthma and rhinitis: Protocol for a systematic review. BMJ Open 2021, 11, e046519. [Google Scholar] [CrossRef]
- Medeleanu, M.V.; Qian, Y.C.; Moraes, T.J.; Subbarao, P. Early-immune development in asthma: A review of the literature. Cell. Immunol. 2023, 393, 104770. [Google Scholar] [CrossRef]
- Woloski, J.R.; Heston, S.; Escobedo Calderon, S.P. Respiratory Allergic Disorders. Prim. Care 2016, 43, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Espinosa, V.; Galusha, L.E.; Rahimian, V.; Miro, K.L.; Rivera-Medina, A.; Kasinathan, C.; Capitle, E.; Aguila, H.A.; Kachlany, S.C. Expression and targeting of lymphocyte function-associated antigen 1 (LFA-1) on white blood cells for treatment of allergic asthma. J. Leukoc. Biol. 2015, 97, 439–446. [Google Scholar] [CrossRef]
- Abdulla, M.; Mohammed, N. A Review on Inflammatory Bowel Diseases: Recent Molecular Pathophysiology Advances. Biol. Targets Ther. 2022, 16, 129–140. [Google Scholar] [CrossRef]
- Imbrizi, M.; Magro, F.; Coy, C.S.R. Pharmacological Therapy in Inflammatory Bowel Diseases: A Narrative Review of the Past 90 Years. Pharmaceuticals 2023, 16, 1272. [Google Scholar] [CrossRef]
- Khan, S.; Sebastian, S.A.; Parmar, M.P.; Ghadge, N.; Padda, I.; Keshta, A.S.; Minhaz, N.; Patel, A. Factors influencing the quality of life in inflammatory bowel disease: A comprehensive review. Dis. Mon. 2023, 70, 101672. [Google Scholar] [CrossRef]
- Mihai, I.R.; Burlui, A.M.; Rezus, I.I.; Mihai, C.; Macovei, L.A.; Cardoneanu, A.; Gavrilescu, O.; Dranga, M.; Rezus, E. Inflammatory Bowel Disease as a Paradoxical Reaction to Anti-TNF-alpha Treatment-A Review. Life 2023, 13, 1779. [Google Scholar] [CrossRef]
- Sousa, P.; Bertani, L.; Rodrigues, C. Management of inflammatory bowel disease in the elderly: A review. Dig. Liver Dis. 2023, 55, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Ostanin, D.V.; Furr, K.L.; Pavlick, K.P.; Gray, L.; Kevil, C.G.; Shukla, D.; D’Souza, D.; Hoffman, J.M.; Grisham, M.B. T cell-associated CD18 but not CD62L, ICAM-1, or PSGL-1 is required for the induction of chronic colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G1706–G1714. [Google Scholar] [CrossRef] [PubMed]
- Pavlick, K.P.; Ostanin, D.V.; Furr, K.L.; Laroux, F.S.; Brown, C.M.; Gray, L.; Kevil, C.G.; Grisham, M.B. Role of T-cell-associated lymphocyte function-associated antigen-1 in the pathogenesis of experimental colitis. Int. Immunol. 2006, 18, 389–398. [Google Scholar] [CrossRef]
- Abdelbaqi, M.; Chidlow, J.H.; Matthews, K.M.; Pavlick, K.P.; Barlow, S.C.; Linscott, A.J.; Grisham, M.B.; Fowler, M.R.; Kevil, C.G. Regulation of dextran sodium sulfate induced colitis by leukocyte beta 2 integrins. Lab. Investig. A J. Tech. Methods Pathol. 2006, 86, 380–390. [Google Scholar] [CrossRef]
- James, D.G.; Seo, D.H.; Chen, J.; Vemulapalli, C.; Stone, C.D. Efalizumab, a human monoclonal anti-CD11a antibody, in the treatment of moderate to severe Crohn’s Disease: An open-label pilot study. Dig. Dis. Sci. 2011, 56, 1806–1810. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kachlany, S.C.; Vega, B.A. Therapeutic Applications of Aggregatibacter actinomycetemcomitans Leukotoxin. Pathogens 2024, 13, 354. https://doi.org/10.3390/pathogens13050354
Kachlany SC, Vega BA. Therapeutic Applications of Aggregatibacter actinomycetemcomitans Leukotoxin. Pathogens. 2024; 13(5):354. https://doi.org/10.3390/pathogens13050354
Chicago/Turabian StyleKachlany, Scott C., and Brian A. Vega. 2024. "Therapeutic Applications of Aggregatibacter actinomycetemcomitans Leukotoxin" Pathogens 13, no. 5: 354. https://doi.org/10.3390/pathogens13050354
APA StyleKachlany, S. C., & Vega, B. A. (2024). Therapeutic Applications of Aggregatibacter actinomycetemcomitans Leukotoxin. Pathogens, 13(5), 354. https://doi.org/10.3390/pathogens13050354