

Molecular Identification of Spotted Fever Group Rickettsiae in Ticks in the Republic of Korea


Abstract
:1. Introduction
2. Materials and Methods
2.1. Tick Collection and Identification
2.2. DNA Extraction
2.3. PCR Amplification
2.4. Phylogenetic Analysis
2.5. Statistical Analyses
3. Results
3.1. The Identification of Tick Species
3.2. Molecular Detection of SFGR in Ticks
3.3. SFGR Species Identification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rochlin, I.; Toledo, A. Emerging tick-borne pathogens of public health importance: A mini-review. J. Med. Microbiol. 2020, 69, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Kim, J.; Choi, Y.J.; Park, H.J.; Jang, W.J. Molecular genetic analysis and clinical characterization of Rickettsia species isolated from the Republic of Korea in 2017. Transbound. Emerg. Dis. 2020, 67, 1447–1452. [Google Scholar] [CrossRef] [PubMed]
- Socolovschi, C.; Mediannikov, O.; Raoult, D.; Parola, P. The relationship between spotted fever group Rickettsiae and ixodid ticks. Vet. Res. 2009, 40, 34. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; de la Fuente, J. The ecology of ticks and epidemiology of tick-borne viral diseases. Antivir. Res. 2014, 108, 104–128. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.E.; et al. Update on tick-borne rickettsioses around the world: A geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Y.; Sun, Y.-Q.; Chen, J.J.; Teng, A.Y.; Wang, T.; Li, H.; Hay, S.I.; Fang, L.Q.; Yang, Y.; Liu, W. Mapping the global distribution of spotted fever group rickettsiae: A systematic review with modelling analysis. Lancet Digit. Health 2023, 5, e5–e15. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Epidemiology and Statistics. Number of Reported Cases of Rocky Mountain Spotted Fever in USA. 2012–2019. Available online: https://www.cdc.gov/rmsf/stats/index.html (accessed on 23 January 2024).
- Luz, H.R.; Costa, F.B.; Benatti, H.R.; Ramos, V.N.; de ASerpa, M.C.; Martins, T.F.; Acosta, I.C.; Ramirez, D.G.; Munoz-Leal, S.; Ramirez-Hernandez, A.; et al. Epidemiology of capybara-associated Brazilian spotted fever. PLoS Negl. Trop. Dis. 2019, 13, e0007734. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Barroso, D.; Vescio, M.F.; Bella, A.; Ciervo, A.; Busani, L.; Rizzo, C.; Rezza, G.; Pezzotti, P. Mediterranean spotted fever rickettsiosis in Italy, 2001–2015: Spatio-temporal distribution based on hospitalization records. Ticks Tick-Borne Dis. 2019, 10, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, H.; Arima, Y.; Shigematsu, M.; Sunagawa, T.; Saijo, M.; Oishi, K.; Ando, S. Descriptive epidemiology of rickettsial infections in Japan: Scrub typhus and Japanese spotted fever, 2007–2016. Int. J. Infect. Dis. 2021, 105, 560–566. [Google Scholar] [CrossRef]
- Jia, N.; Liu, H.-B.; Zheng, Y.C.; Shi, W.Q.; Wei, R.; Chu, Y.L.; Ning, N.Z.; Jiang, B.G.; Jiang, R.R.; Li, T.; et al. Cutaneous immunoprofiles of three spotted fever group rickettsia cases. Infect. Immun. 2020, 88, e00686-19. [Google Scholar] [CrossRef]
- Jang, W.-J.; Kim, J.H.; Choi, Y.J.; Jung, K.D.; Kim, Y.G.; Lee, S.H.; Choi, M.S.; Kim, I.S.; Walker, D.H.; Park, K.H. First serologic evidence of human spotted fever group rickettsiosis in Korea. J. Clin. Microbiol. 2004, 42, 2310–2313. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.J.; Jang, W.J.; Kim, J.H.; Ryu, J.S.; Lee, S.H.; Park, K.H.; Paik, H.S.; Koh, Y.S.; Choi, M.S.; Kim, I.S. Spotted fever group and typhus group rickettsioses in humans, South Korea. Emerg. Infect. Dis. 2005, 11, 237–244. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, H.S.; Jung, K.D.; Jang, W.J.; Koh, S.E.; Kang, S.S.; Lee, I.Y.; Lee, W.J.; Kim, B.J.; Kook, Y.H.; et al. Identification of the spotted fever group rickettsiae detected from Haemaphysalis longicornis in Korea. Microbiol. Immunol. 2003, 47, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Choi, Y.J.; Shin, S.H.; Choi, M.K.; Song, H.J.; Kim, H.C.; Klein, T.A.; Richards, A.L.; Park, K.H.; Jang, W.J. Spotted fever group rickettsia closely related to Rickettsia monacensis isolated from ticks in South Jeolla province, Korea. Microbiol. Immunol. 2013, 57, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Noh, Y.; Lee, Y.S.; Kim, H.C.; Chong, S.T.; Klein, T.A.; Jiang, J.; Richards, A.L.; Lee, H.K.; Kim, S.Y. Molecular detection of Rickettsia species in ticks collected from the southwestern provinces of the Republic of Korea. Parasites Vectors 2017, 10, 20. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Kim, C.-M.; Kim, D.M.; Yun, N.R. Case report: Coinfection with Rickettsia monacensis and Orientia tsutsugamushi. Am. J. Trop. Med. Hyg. 2019, 101, 332–335. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Choi, Y.J.; Lee, K.M.; Ahn, K.J.; Kim, H.C.; Klein, T.; Jiang, J.; Richards, A.; Park, K.H.; Jang, W.J. First isolation of Rickettsia monacensis from a patient in South Korea. Microbiol. Immunol. 2017, 61, 258–263. [Google Scholar] [CrossRef]
- Jiang, J.; Choi, Y.J.; Kim, J.; Kim, H.C.; Klein, T.A.; Chong, S.T.; Richards, A.L.; Park, H.J.; Shin, S.H.; Song, D.; et al. Distribution of Rickettsia spp. in ticks from northwestern and southwestern provinces, Republic of Korea. Korean J. Parasitol. 2019, 57, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Lee, S.H.; Lee, G.S.; Seo, J.J.; Chung, J.K. Epidemiological characteristics of field tick-borne pathogens in Gwang-ju metropolitan area, South Korea, from 2014 to 2018. Osong Public Health Res. Perspect. 2020, 11, 177–184. [Google Scholar] [CrossRef]
- Seo, J.W.; Han, S.Y.; Sung, S.H.; Jung, E.Y.; Kim, J.H.; Lee, S.J.; Yoo, S.S. Survey on tick distribution and tick-borne pathogens in Daejeon and adjacent areas in South Korea. Ticks Tick-Borne Dis. 2021, 12, 101711. [Google Scholar] [CrossRef]
- Yamaguti, N.; Tipton, V.J.; Keegan, H.L.; Toshioka, S. Ticks of Japan, Korea, and the Ryukyu islands. Brigh. Young Univ. Sci. Bull. 1971, 15, 1. [Google Scholar]
- Seo, M.G.; Noh, B.E.; Lee, H.S.; Kim, T.K.; Song, B.G.; Lee, H.I. Nationwide temporal and geographical distribution of tick populations and phylogenetic analysis of severe fever with thrombocytopenia syndrome virus in ticks in Korea, 2020. Microorganisms 2021, 9, 1630. [Google Scholar] [CrossRef]
- Bang, M.S.; Kim, C.M.; Pyun, S.H.; Kim, D.M.; Yun, N.R. Molecular investigation of tick-borne pathogens in ticks removed from tick-bitten humans in the southwestern region of the Republic of Korea. PLoS ONE 2021, 16, e0252992. [Google Scholar] [CrossRef]
- Arai, R.; Sato, M.; Kato, M.; Aoki, J.; Nishida, A.; Watanabe, K.; Hirokawa, C.; Ikeda, S.; Watanabe, K.; Regilme, M.A.; et al. Spotted fever group rickettsiae (SFGR) detection in ticks following reported human case of Japanese spotted fever in Niigata Prefecture, Japan. Sci. Rep. 2021, 11, 2595. [Google Scholar] [CrossRef]
- Knoll, S.; Springer, A.; Hauck, D.; Schunack, B.; Pachnicke, S.; Strube, C. Regional, seasonal, biennial and landscape-associated distribution of Anaplasma phagocytophilum and Rickettsia spp. infections in Ixodes ticks in northern Germany and implications for risk assessment at larger spatial scales. Ticks Tick-Borne Dis. 2021, 12, 101657. [Google Scholar] [CrossRef]
- Okado, K.; Moumouni, P.F.A.; Lee, S.H.; Sivakumar, T.; Yokoyama, N.; Fujisaki, K.; Suzuki, H.; Xuan, X.; Umemiya-Shirafuji, R. Molecular detection of borrelia burgdorferi (Sensu lato) and Rickettsia spp. in hard ticks distributed in tokachi district, Eastern Hokkaido, Japan. Curr. Res. Parasitol. Vector Borne Dis. 2021, 1, 100059. [Google Scholar] [CrossRef]
- Wang, Q.; Guo, W.B.; Pan, Y.S.; Jiang, B.G.; Du, C.H.; Que, T.C.; Zhan, L.; Wu, J.H.; Yu, M.H.; Cui, X.M.; et al. Detection of novel spotted fever group Rickettsiae (Rickettsiales: Rickettsiaceae) in ticks (Acari: Ixodidae) in Southwestern China. J. Med. Entomol. 2021, 58, 1363–1369. [Google Scholar] [CrossRef]
- Špitalská, E.; Stanko, M.; Mošanský, L.; Kraljik, J.; Miklisová, D.; Mahríková, L.; Bona, M.; Kazimírová, M. Seasonal analysis of Rickettsia species in ticks in an agricultural site of Slovakia. Exp. Appl. Acarol. 2016, 68, 315–324. [Google Scholar] [CrossRef]
- Piranda, E.M.; Faccini, J.L.H.; Pinter, A.; Pacheco, R.C.; Cançado, P.H.; Labruna, M.B. Experimental infection of Rhipicephalus sanguineus ticks with the bacterium Rickettsia rickettsii, using experimentally infected dogs. Vector-Borne Zoonotic Dis. 2011, 11, 29–36. [Google Scholar] [CrossRef]
- Stanley, H.M.; Ford, S.L.; Snellgrove, A.N.; Hartzer, K.; Smith, E.B.; Krapiunaya, I.; Levin, M.L. The ability of the invasive Asian longhorned tick Haemaphysalis longicornis (Acari: Ixodidae) to acquire and transmit Rickettsia rickettsii (Rickettsiales: Rickettsiaceae), the agent of Rocky Mountain spotted fever, under laboratory conditions. J. Med. Entomol. 2020, 57, 1635–1639. [Google Scholar] [CrossRef]
- Hauck, D.; Jordan, D.; Springer, A.; Schunack, B.; Pachnicke, S.; Fingerle, V.; Strube, C. Transovarial transmission of Borrelia spp., Rickettsia spp. and Anaplasma phagocytophilum in Ixodes ricinus under field conditions extrapolated from DNA detection in questing larvae. Parasites Vectors 2020, 13, 176. [Google Scholar] [CrossRef]
- Matsumoto, K.; Ogawa, M.; Brouqui, P.; Raoult, D.; Parola, P. Transmission of Rickettsia massiliae in the tick, Rhipicephalus turanicus. Med. Vet. Entomol. 2005, 19, 263–270. [Google Scholar] [CrossRef]
- Climates to Travel. Average Weather, Temperature, Rainfall, Sunshine of Slovakia and Germany. Available online: https://www.climatestotravel.com/climate (accessed on 23 January 2024).
- Piedmonte, N.P.; Vinci, V.C.; Daniels, T.J.; Backenson, B.P.; Falco, R.C. Seasonal activity of Haemaphysalis longicornis (Acari: Ixodidae) in southern New York state. J. Med. Entomol. 2021, 58, 676–681. [Google Scholar] [CrossRef]
- Liu, H.; Li, Q.; Zhang, X.; Li, Z.; Wang, Z.; Song, M.; Wei, F.; Wang, S.; Liu, Q. Characterization of rickettsiae in ticks in northeastern China. Parasites Vectors 2016, 9, 498. [Google Scholar] [CrossRef]
- Liu, H.; Liang, X.; Wang, H.; Sun, X.; Bai, X.; Hu, B.; Shi, N.; Wang, N.; Zhang, X.; Huang, L.; et al. Molecular evidence of the spotted fever group Rickettsiae in ticks from Yunnan Province, Southwest China. Exp. Appl. Acarol. 2020, 80, 339–348. [Google Scholar] [CrossRef]
- Guo, W.P.; Wang, Y.H.; Lu, Q.; Xu, G.; Luo, Y.; Ni, X.; Zhou, E.M. Molecular detection of spotted fever group rickettsiae in hard ticks, northern China. Transbound. Emerg. Dis. 2019, 66, 1587–1596. [Google Scholar] [CrossRef]
- Lu, M.; Meng, C.; Zhang, B.; Wang, X.; Tian, J.; Tang, G.; Wang, W.; Li, N.; Li, M.; Xu, X.; et al. Prevalence of Spotted Fever Group Rickettsia and Candidatus Lariskella in Multiple Tick Species from Guizhou Province, China. Biomolecules 2022, 12, 1701. [Google Scholar] [CrossRef]
- Qi, Y.; Ai, L.; Jiao, J.; Wang, J.; Wu, D.; Wang, P.; Zhang, G.; Qin, Y.; Hu, C.; Lv, R.; et al. High prevalence of Rickettsia spp. in ticks from wild hedgehogs rather than domestic bovine in Jiangsu province, Eastern China. Front. Cell. Infect. Microbiol. 2022, 12, 954785. [Google Scholar] [CrossRef]
- Seo, J.Y.; Kim, Y.J.; Kim, S.Y.; Lee, H.I. Molecular Detection of Anaplasma, Ehrlichia and Rickettsia Pathogens in Ticks Collected from Humans in the Republic of Korea, 2021. Pathogens 2023, 12, 802. [Google Scholar] [CrossRef]
- Fournier, P.E.; Raoult, D. Tick-borne spotted fever Rickettsioses. In Hunter’s Tropical Medicine and Emerging Infectious Diseases; Elsevier: Amsterdam, The Netherlands, 2020; pp. 587–593. [Google Scholar]
- Shin, S.H.; Seo, H.J.; Choi, Y.J.; Choi, M.K.; Kim, H.C.; Klein, T.A.; Chong, S.T.; Richards, A.L.; Lee, K.H.; Jang, W.J. Detection of Rickettsia monacensis from Ixodes nipponensis collected from rodents in Gyeonggi and Gangwon Provinces, Republic of Korea. Exp. Appl. Acarol. 2013, 61, 337–347. [Google Scholar] [CrossRef]
- Truong, A.T.; Yun, B.R.; Yoo, M.S.; Lim, J.; Min, S.; Yoon, S.S.; Yun, Y.M.; Kim, J.T.; Cho, Y.S. Utility of ultra-rapid real-time PCR for detection and prevalence of Rickettsia spp. in ticks. BMC Vet. Res. 2022, 18, 199. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; An, H.; Lee, J.S.; O’Guinn, M.L.; Kim, H.C.; Chong, S.T.; Zhang, Y.; Song, D.; Burrus, R.G.; Bao, Y.; et al. Molecular characterization of Haemaphysalis longicornis-borne rickettsiae, Republic of Korea and China. Ticks Tick-Borne Dis. 2018, 9, 1606–1613. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; Lia, R.P.; Capelli, G.; Otranto, D. Efficiency of flagging and dragging for tick collection. Exp. Appl. Acarol. 2013, 61, 119–127. [Google Scholar] [CrossRef]
- Mays, S.E.; Houston, A.E.; Trout Fryxell, R.T. Comparison of novel and conventional methods of trapping ixodid ticks in the southeastern USA. Med. Vet. Entomol. 2016, 30, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Kho, J.W.; Lee, W.G.; Roh, J.Y.; Lee, D.H. Seasonal occurrence of Haemaphysalis longicornis (Acari: Ixodidae) and Haemaphysalis flava, vectors of severe fever with thrombocytopenia syndrome (SFTS) in South Korea. J. Med. Entomol. 2019, 56, 1139–1144. [Google Scholar] [CrossRef] [PubMed]
- Fracasso, G.; Grillini, M.; Grassi, L.; Gradoni, F.; Rold, G.; Bertola, M. Effective Methods of Estimation of Pathogen Prevalence in Pooled Ticks. Pathogens 2023, 12, 557. [Google Scholar] [CrossRef] [PubMed]
- Halos, L.; Jamal, T.; Vial, L.; Maillard, R.; Suau, A.; Le Menach, A.; Boulouis, H.J.; Vayssier-Taussat, M. Determination of an efficient and reliable method for DNA extraction from ticks. Vet. Res. 2004, 35, 709–713. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K. Rickettsia-host-tick interactions: Knowledge advances and gaps. Infect. Immun. 2022, 90, e00621-21. [Google Scholar] [CrossRef]
- Sul, H.; Kim, D.M. Present state and future of tick-borne infectious diseases in Korea. J. Korean Med. Assoc. 2017, 60, 475–483. [Google Scholar] [CrossRef]
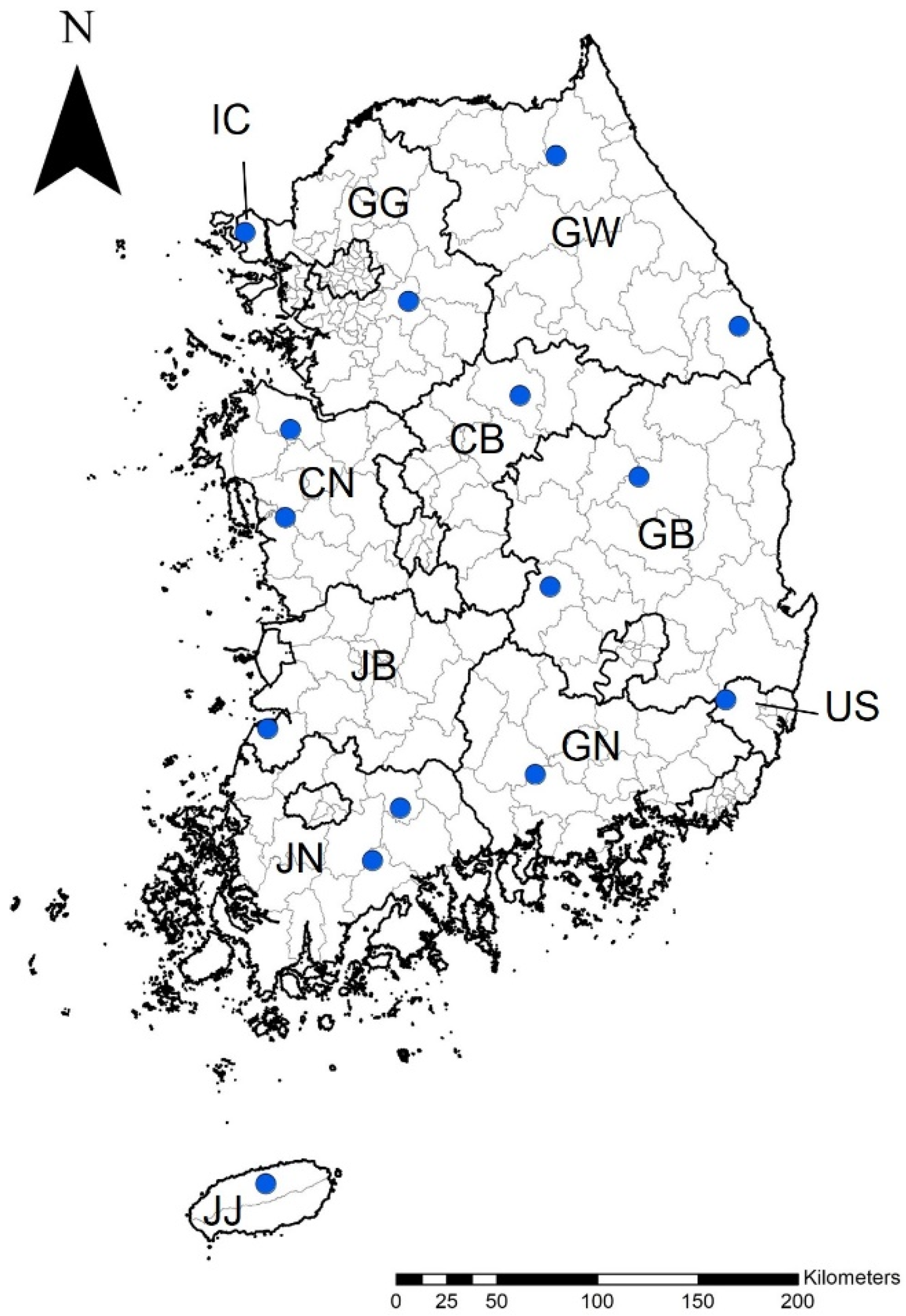
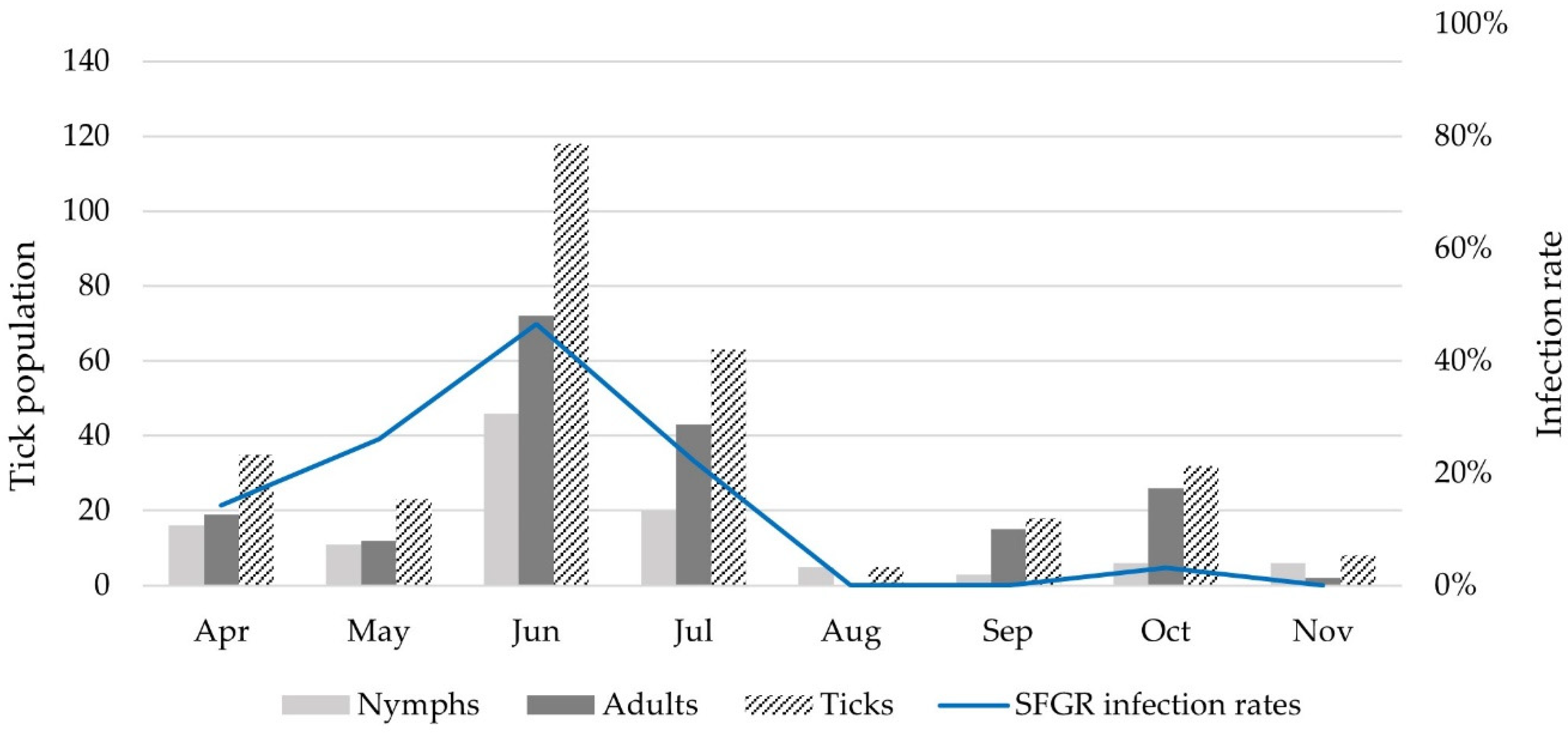


{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Name | Nucleotide Sequence (5’-3’) | Product Size (bp) | PCR Conditions | Reference | |
|---|---|---|---|---|---|---|
| 17 kDa | 1st | Rr17k.1p | TTTACAAAATTCTAAAAACCAT | 539 | 95 °C/5 m; 35 cycles: 95 °C/30 s, 57 °C/1 m, 72 °C/2 m; 72 °C/5 m | [24] |
| Rr17k.539n | TCAATTCACAACTTGCCATT | |||||
| 2nd | Rr17k.90p | GCTCTTGCAACTTCTATGTT | 450 | |||
| Rr17k.539n | TCAATTCACAACTTGCCATT | |||||
| ompA | 1st | R190.70F | ATGGCGAATATTTCTCCAAAA | 634 | 94 °C/5 m; 40 cycles: 94 °C/30 s, 50 °C/30 s, 72 °C/1 m; 72 °C/5 m | |
| RR190.701R | GTTCCGTTAATGGCAGCATCT | |||||
| 2nd | R190.70F | ATGGCGAATATTTCTCCAAAAA | 535 | 94 °C/5 m; 5 cycles: 94 °C/30 s, 50 °C/30 s, 72 °C/30 s; 30 cycles: 94 °C/30 s, 54 °C/30 s, 72 °C/30 s; 72 °C/5 m | ||
| RR190.602R | AGTGCAGCATTCGCTCCCCCT | |||||
| gltA | 1st | RpCS.780p | GACCATGAGCAGAATGCTTCT | 479 | 95 °C/5 m; 35 cycles: 95 °C/30 s, 44 °C/30 s, 65 °C/2 m; 65 °C/5 m | [25] |
| RpCS.1258n | ATTGCAAAAAGTACAGTGAACA | |||||
| 2nd | Rsfg.77p | GGGGGCCTGCTCACGGCGG | 382 | 95 °C/5 m; 35 cycles: 95 °C/30 s, 48 °C/30 s, 65 °C/2 m; 65 °C/5 m | ||
| Rsfg.1258n | ATTGCAAAAAGTACAGTGAACA | |||||
| Tick Species | Stages | Positive SFGR | Positive SFGR (%) | Total Positive SFGR (%) | p-Value | ||
|---|---|---|---|---|---|---|---|
| R. Monacensis | R. Heilong-Jiangensis | Candidatus R. Jingxinensis | |||||
| Haemaphysalis longicornis | Female | 0 | 0 | 19 | 19/73 (26.0) | 77/204 (37.7) | <0.0001 |
| Male | 0 | 0 | 36 | 36/54 (66.7) | |||
| Nymph | 0 | 0 | 22 | 22/77 (28.6) | |||
| Haemaphysalis flava | Female | 0 | 0 | 0 | 0/30 (0.0) | 3/83 (3.6) | - |
| Male | 0 | 1 | 0 | 1/24 (4.2) | |||
| Nymph | 0 | 0 | 2 | 2/29 (6.9) | |||
| Amblyomma testudinarium | Female | 0 | 0 | 0 | 0/4 (0.0) | 0/11 (0.0) | 0.5216 |
| Male | 0 | 0 | 0 | 0 | |||
| Nymph | 0 | 0 | 0 | 0/7 (0.0) | |||
| Ixodes nipponensis | Female | 0 | 0 | 0 | 0/2 (0.0) | 1/4 (25.0) | 0.0461 |
| Male | 1 | 0 | 0 | 1/2 (50.0) | |||
| Nymph | 0 | 0 | 0 | 0 | |||
| Total | 1 (0.3) | 1 (0.3) | 79 (26.2) | 81/302 (26.8) | - | ||
| Region | Tick Species | No. of Ticks | Positive SFGR (%) | p-Value | ||
|---|---|---|---|---|---|---|
| Metropolitan area | Incheon | Ganghwa-gun | H. longicornis | 15 | 9/19(47.4) | - |
| H.flava | 4 | |||||
| Ulsan | Ulju-gun | H. longicornis | 15 | 8/26 (30.8) | 0.2566 | |
| H.flava | 5 | |||||
| A. testudinarium | 5 | |||||
| I. nipponensis | 1 | |||||
| Provinces | Gyeonggi | Gwangju-si | H. longicornis | 10 | 0/20 (0.0) | 0.0004 |
| H.flava | 10 | |||||
| Gangwon | Inje-gun | H. longicornis | 13 | 2/18 (11.1) | 0.0159 | |
| H.flava | 5 | |||||
| Samcheok-si | H. longicornis | 15 | 9/20 (45.0) | 0.8821 | ||
| H.flava | 5 | |||||
| Chungcheong-buk | Chungju-si | H. longicornis | 10 | 6/14 (42.9) | 0.7970 | |
| H.flava | 3 * | |||||
| Chungcheong-nam | Dangjin-si | H. longicornis | 15 | 7/24 (29.2) | 0.2201 | |
| H.flava | 9 | |||||
| Boryeong-si | H. longicornis | 13 | 5/17 (29.4) | 0.2699 | ||
| H.flava | 4 | |||||
| Gyeongsang-buk | Andong-si | H. longicornis | 10 | 6/17 (35.3) | 0.4632 | |
| H.flava | 7 | |||||
| Gimcheon-si | H. longicornis | 15 | 8/20 (40.0) | 0.6428 | ||
| H.flava | 1 | |||||
| A. testudinarium | 3 | |||||
| I. nipponensis | 1 | |||||
| Gyeongsang-nam | Jinju-si | H. longicornis | 14 | 6/17 (35.3) | 0.4632 | |
| H.flava | 1 | |||||
| I. nipponensis | 12 * | |||||
| Jeolla-buk | Gochang-gun | H. longicornis | 15 | 6/19 (31.6) | 0.3194 | |
| H.flava | 1 | |||||
| A. testudinarium | 3 | |||||
| Jeolla-nam | Gokseong-gun | H. longicornis | 16 | 2/23 (8.7) | 0.0046 | |
| H.flava | 7 | |||||
| Boseong-gun | H. longicornis | 14 | 0/24 (0.0) | 0.0001 | ||
| H.flava | 10 | |||||
| Jeju | Jeju-si | H. longicornis | 14 | 7/24 (29.2) | 0.2201 | |
| H.flava | 10 | |||||
| Total | 302 | 81 (26.8) | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, J.-Y.; Park, J.-S.; Lee, H.-I.; Ju, J.-W. Molecular Identification of Spotted Fever Group Rickettsiae in Ticks in the Republic of Korea. Pathogens 2024, 13, 575. https://doi.org/10.3390/pathogens13070575
Seo J-Y, Park J-S, Lee H-I, Ju J-W. Molecular Identification of Spotted Fever Group Rickettsiae in Ticks in the Republic of Korea. Pathogens. 2024; 13(7):575. https://doi.org/10.3390/pathogens13070575
Chicago/Turabian StyleSeo, Ji-Ye, Jin-Seo Park, Hee-Il Lee, and Jung-Won Ju. 2024. "Molecular Identification of Spotted Fever Group Rickettsiae in Ticks in the Republic of Korea" Pathogens 13, no. 7: 575. https://doi.org/10.3390/pathogens13070575