
The Genomic Epidemiology of Clinical Burkholderia pseudomallei Isolates in North Queensland, Australia
 , and
, and
Abstract
:1. Introduction
2. Ethical Approval
3. Methods
Statistical Analysis
4. Results
4.1. Multilocus Sequence Type
4.2. Virulence Factors
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gassiep, I.; Armstrong, M.; Norton, R. Human Melioidosis. Clin. Microbiol. Rev. 2020, 33, e00006-19. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.D.; Smith, S.; Binotto, E.; McBride, W.J.; Currie, B.J.; Hanson, J. The epidemiology and clinical features of melioidosis in Far North Queensland: Implications for patient management. PLoS Negl. Trop. Dis. 2017, 11, e0005411. [Google Scholar] [CrossRef] [PubMed]
- Currie, B.J.; Mayo, M.; Ward, L.M.; Kaestli, M.; Meumann, E.M.; Webb, J.R.; Woerle, C.; Baird, R.W.; Price, R.N.; Marshall, C.S.; et al. The Darwin Prospective Melioidosis Study: A 30-year prospective, observational investigation. Lancet Infect. Dis. 2021, 21, 1737–1746. [Google Scholar] [CrossRef] [PubMed]
- Gassiep, I.; Ganeshalingam, V.; Chatfield, M.D.; Harris, P.N.A.; Norton, R.E. The epidemiology of melioidosis in Townsville, Australia. Trans. R. Soc. Trop. Med. Hyg. 2022, 116, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Pearson, T.; Giffard, P.; Beckstrom-Sternberg, S.; Auerbach, R.; Hornstra, H.; Tuanyok, A.; Price, E.P.; Glass, M.B.; Leadem, B.; Beckstrom-Sternberg, J.S.; et al. Phylogeographic reconstruction of a bacterial species with high levels of lateral gene transfer. BMC Biol. 2009, 7, 78. [Google Scholar] [CrossRef]
- McRobb, E.; Kaestli, M.; Price, E.P.; Sarovich, D.S.; Mayo, M.; Warner, J.; Spratt, B.G.; Currie, B.J. Distribution of Burkholderia pseudomallei in Northern Australia, a Land of Diversity. Appl. Environ. Microbiol. 2014, 80, 3463–3468. [Google Scholar] [CrossRef] [PubMed]
- Godoy, D.; Randle, G.; Simpson, A.J.; Aanensen, D.M.; Pitt, T.L.; Kinoshita, R.; Spratt, B.G. Multilocus sequence typing and evolutionary relationships among the causative agents of melioidosis and glanders, Burkholderia pseudomallei and Burkholderia mallei. J. Clin. Microbiol. 2003, 41, 2068–2079. [Google Scholar] [CrossRef]
- McRobb, E.; Sarovich, D.S.; Price, E.P.; Kaestli, M.; Mayo, M.; Keim, P.; Currie, B.J. Tracing Melioidosis Back to the Source: Using Whole-Genome Sequencing to Investigate an Outbreak Originating from a Contaminated Domestic Water Supply. J. Clin. Microbiol. 2015, 53, 1144–1148. [Google Scholar] [CrossRef]
- Chapple, S.N.; Price, E.P.; Sarovich, D.S.; McRobb, E.; Mayo, M.; Kaestli, M.; Spratt, B.G.; Currie, B.J. Burkholderia pseudomallei Genotype Distribution in the Northern Territory, Australia. Am. J. Trop. Med. Hyg. 2016, 94, 68–72. [Google Scholar] [CrossRef]
- Balaji, V.; Perumalla, S.; Perumal, R.; Inbanathan, F.Y.; Rajamani Sekar, S.K.; Paul, M.M.; Sahni, R.D.; Prakash, J.A.J.; Iyadurai, R. Multi locus sequence typing of Burkholderia pseudomallei isolates from India unveils molecular diversity and confers regional association in Southeast Asia. PLoS Neglected Trop. Dis. 2018, 12, e0006558. [Google Scholar] [CrossRef]
- Fang, Y.; Zhu, P.; Li, Q.; Chen, H.; Li, Y.; Ren, C.; Hu, Y.; Tan, Z.; Gu, J.; Mao, X. Multilocus sequence typing of 102 Burkholderia pseudomallei strains isolated from China. Epidemiol. Infect. 2016, 144, 1917–1923. [Google Scholar] [CrossRef] [PubMed]
- Zueter, A.R.; Rahman, Z.A.; Abumarzouq, M.; Harun, A. Multilocus sequence types of clinical Burkholderia pseudomallei isolates from peninsular Malaysia and their associations with disease outcomes. BMC Infect. Dis. 2018, 18, 5. [Google Scholar] [CrossRef] [PubMed]
- Tellapragada, C.; Kamthan, A.; Shaw, T.; Ke, V.; Kumar, S.; Bhat, V.; Mukhopadhyay, C. Unravelling the Molecular Epidemiology and Genetic Diversity among Burkholderia pseudomallei Isolates from South India Using Multi-Locus Sequence Typing. PLoS ONE 2016, 11, e0168331. [Google Scholar] [CrossRef] [PubMed]
- Sarovich, D.S.; Price, E.P.; Webb, J.R.; Ward, L.M.; Voutsinos, M.Y.; Tuanyok, A.; Mayo, M.; Kaestli, M.; Currie, B.J. Variable Virulence Factors in Burkholderia pseudomallei (Melioidosis) Associated with Human Disease. PLoS ONE 2014, 9, e91682. [Google Scholar] [CrossRef]
- Burnard, D.; Bauer, M.J.; Falconer, C.; Gassiep, I.; Norton, R.E.; Paterson, D.L.; Harris, P.N. Clinical Burkholderia pseudomallei isolates from north Queensland carry diverse bimABm genes that are associated with central nervous system disease and are phylogenomically distinct from other Australian strains. PLoS Negl. Trop. Dis. 2022, 16, e0009482. [Google Scholar] [CrossRef] [PubMed]
- Tuanyok, A.; Auerbach, R.K.; Brettin, T.S.; Bruce, D.C.; Munk, A.C.; Detter, J.C.; Pearson, T.; Hornstra, H.; Sermswan, R.W.; Wuthiekanun, V.; et al. A Horizontal Gene Transfer Event Defines Two Distinct Groups within Burkholderia pseudomallei That Have Dissimilar Geographic Distributions. J. Bacteriol. 2007, 189, 9044–9049. [Google Scholar] [CrossRef]
- Gassiep, I.; Burnard, D.; Permana, B.; Bauer, M.J.; Cuddihy, T.; Forde, B.M.; Chatfield, M.D.; Ling, W.; Norton, R.; Harris, P.N.A. Melioidosis Queensland: An analysis of clinical outcomes and genomic factors. PLoS Negl. Trop. Dis. 2023, 17, e0011697. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Treangen, T.J.; Ondov, B.D.; Koren, S.; Phillippy, A.M. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014, 15, 524. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. 2016. Abricate: Mass Screening of Contigs for Antibiotic Resistance Genes. GitHub: San Francisco, CA, USA. Available online: https://github.com/tseemann/abricate (accessed on 9 March 2023).
- Webb, J.R.; Sarovich, D.S.; Price, E.P.; Ward, L.M.; Mayo, M.; Currie, B.J. Burkholderia pseudomallei Lipopolysaccharide Genotype Does Not Correlate with Severity or Outcome in Melioidosis: Host Risk Factors Remain the Critical Determinant. Open Forum Infect. Dis. 2019, 6, ofz091. [Google Scholar] [CrossRef]
- Corkeron, M.L.; Norton, R.; Nelson, P.N. Spatial analysis of melioidosis distribution in a suburban area. Epidemiol. Infect. 2010, 138, 1346–1352. [Google Scholar] [CrossRef] [PubMed]
- Rachlin, A.; Mayo, M.; Webb, J.R.; Kleinecke, M.; Rigas, V.; Harrington, G.; Currie, B.J.; Kaestli, M. Whole-genome sequencing of Burkholderia pseudomallei from an urban melioidosis hot spot reveals a fine-scale population structure and localised spatial clustering in the environment. Sci. Rep. 2020, 10, 5443. [Google Scholar] [CrossRef]
- Australian Bureau of Statistics. 2021 Census. 2021. Available online: https://www.abs.gov.au/census (accessed on 9 March 2023).
- Gora, H.; Hasan, T.; Smith, S.; Wilson, I.; Mayo, M.; Woerle, C.; Webb, J.R.; Currie, B.J.; Hanson, J.; Meumann, E.M. Melioidosis of the central nervous system; impact of the bimABm allele on patient presentation and outcome. Clin. Infect. Dis. 2022, 78, 968–975. [Google Scholar] [CrossRef]
- Tuanyok, A.; Stone, J.K.; Mayo, M.; Kaestli, M.; Gruendike, J.; Georgia, S.; Warrington, S.; Mullins, T.; Allender, C.J.; Wagner, D.M.; et al. The genetic and molecular basis of O-antigenic diversity in Burkholderia pseudomallei lipopolysaccharide. PLoS Negl. Trop. Dis. 2012, 6, e1453. [Google Scholar] [CrossRef]
- Norris, M.H.; Schweizer, H.P.; Tuanyok, A. Structural diversity of Burkholderia pseudomallei lipopolysaccharides affects innate immune signaling. PLoS Negl. Trop. Dis. 2017, 11, e0005571. [Google Scholar] [CrossRef] [PubMed]
- Gassiep, I.; Ganeshalingam, V.; Chatfield, M.D.; Harris, P.N.A.; Norton, R.E. Melioidosis: Laboratory Investigations and Association with Patient Outcomes. Am. J. Trop. Med. Hyg. 2021, 106, 54–59. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Novel ST N (%) | Bivariate p-Value | aOR (95% CI) | Multivariate p-Value | |||
|---|---|---|---|---|---|---|
| Time period | 1997–2004 | 10/40 (20) | <0.001 | Ref | ||
| 2005–2012 | 13/30 (43) | 3.8 (0.9–16.1) | 0.07 | |||
| 2013–2020 | 28/48 (58) | 9.1 (2.5–32.9) | 0.001 | |||
| Clinical data | ||||||
| Age > 50 y | Yes | 33/91 (36) | 0.2 | |||
| No | 18/37 (49) | |||||
| Sex | Male | 36/88 (44) | 0.3 | |||
| Female | 15/46 (33) | |||||
| First Nations | Yes | 13/34 (38) | 0.7 | |||
| No | 35/81 (43) | |||||
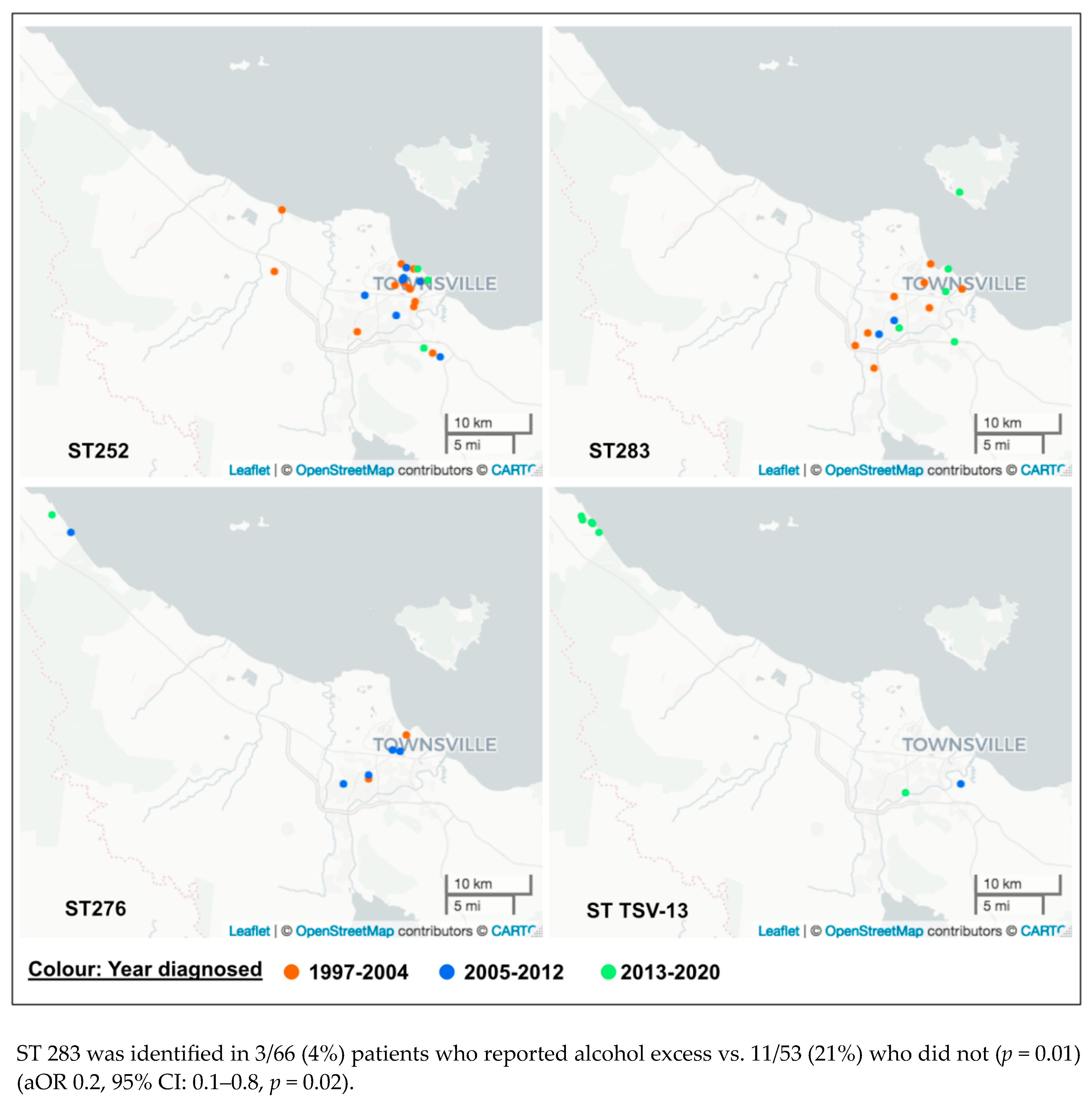
| Alcohol excess | Yes | 30/66 (45) | 0.5 | |||
| No | 20/53 (38) | |||||
| Diabetes mellitus | Yes | 23/59 (39) | 1.0 | |||
| No | 26/65 (40) | |||||
| Chronic kidney disease | Yes | 5/7 (71) | 0.1 | 3.1 (0.4–20.9) | 0.2 | |
| No | 42/107 (39) | |||||
| Malignancy | Yes | 5/16 (31) | 0.6 | |||
| No | 44/106 (42) | |||||
| Lung disease | Yes | 16/37 (43) | 0.7 | |||
| No | 33/87 (38) | |||||
| No risk factors | Yes | 2/13 (15) | 0.1 | |||
| No | 47/112 (42) | |||||
| Bacteraemia | Yes | 36/87 (41) | 0.55 | |||
| No | 13/37 (35) | |||||
| Pneumonia | Yes | 33/84 (39) | 0.68 | |||
| No | 16/37 (43) | |||||
| Solid organ abscess | Yes | 5/15 (33) | 1.0 | |||
| No | 42/103 (41) | |||||
| Skin and soft tissue | Yes | 7/17 (41) | 1.0 | |||
| No | 42/103 (41) | |||||
| Genitourinary | Yes | 8/19 (42) | 1.0 | |||
| No | 41/102 (40) | |||||
| Septic shock | Yes | 38/90 (42) | 1.0 | |||
| No | 10/24 (42) | |||||
| Virulence factors | ||||||
| LPS A | Yes | 28/89 (31) | 0.003 | 0.2 (0.1–0.6) | 0.004 | |
| No | 23/39 (59) | |||||
| YLF | Yes | 26/88 (30) | <0.001 | 0.8 (0.3–2.2) | 0.6 | |
| No | 25/40 (62) | |||||
| fhaB3 | Yes | 41/105 (39) | 0.69 | |||
| No | 10/23 (43) | |||||
| bimABm | Yes | 16/18 (89) | <0.001 | 41.2 (6.7–251) | <0.001 | |
| No | 35/110 (32) | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gassiep, I.; Chatfield, M.D.; Permana, B.; Burnard, D.; Bauer, M.J.; Cuddihy, T.; Forde, B.M.; Mayer-Coverdale, J.; Norton, R.E.; Harris, P.N.A. The Genomic Epidemiology of Clinical Burkholderia pseudomallei Isolates in North Queensland, Australia. Pathogens 2024, 13, 584. https://doi.org/10.3390/pathogens13070584
Gassiep I, Chatfield MD, Permana B, Burnard D, Bauer MJ, Cuddihy T, Forde BM, Mayer-Coverdale J, Norton RE, Harris PNA. The Genomic Epidemiology of Clinical Burkholderia pseudomallei Isolates in North Queensland, Australia. Pathogens. 2024; 13(7):584. https://doi.org/10.3390/pathogens13070584
Chicago/Turabian StyleGassiep, Ian, Mark D. Chatfield, Budi Permana, Delaney Burnard, Michelle J. Bauer, Thom Cuddihy, Brian M. Forde, Johanna Mayer-Coverdale, Robert E. Norton, and Patrick N. A. Harris. 2024. "The Genomic Epidemiology of Clinical Burkholderia pseudomallei Isolates in North Queensland, Australia" Pathogens 13, no. 7: 584. https://doi.org/10.3390/pathogens13070584