
First Genomic Evidence of California Hare Coltivirus from Natural Populations of Ixodes persulcatus Ticks in Northeast China

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Sample Collection
2.2. Extraction of Viral RNA and Next-Generation Sequencing (NGS)
2.3. Real-Time RT-PCR for CHCV
2.4. Phylogenetic Analysis
2.5. Relationship between Coltiviruses and Host
3. Results
3.1. Identification of CHCV in Ha. concinna by NGS
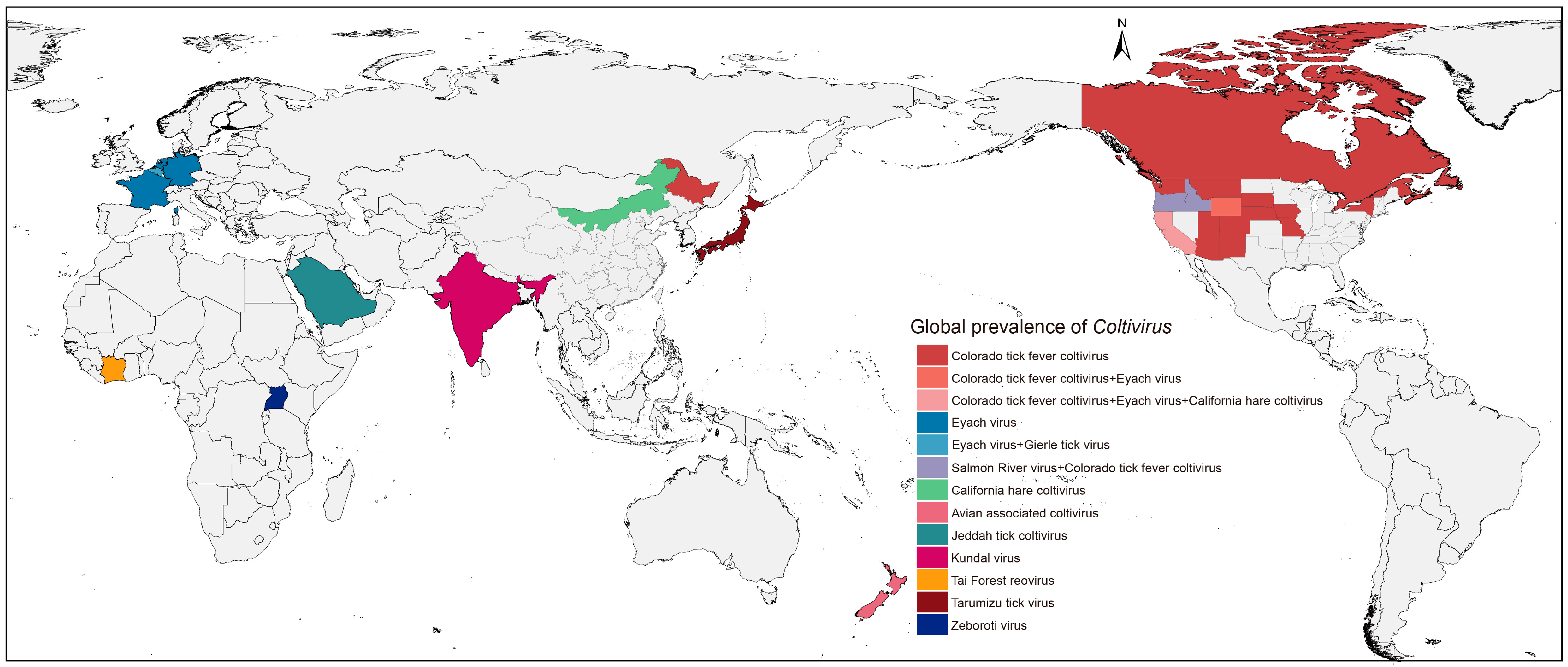
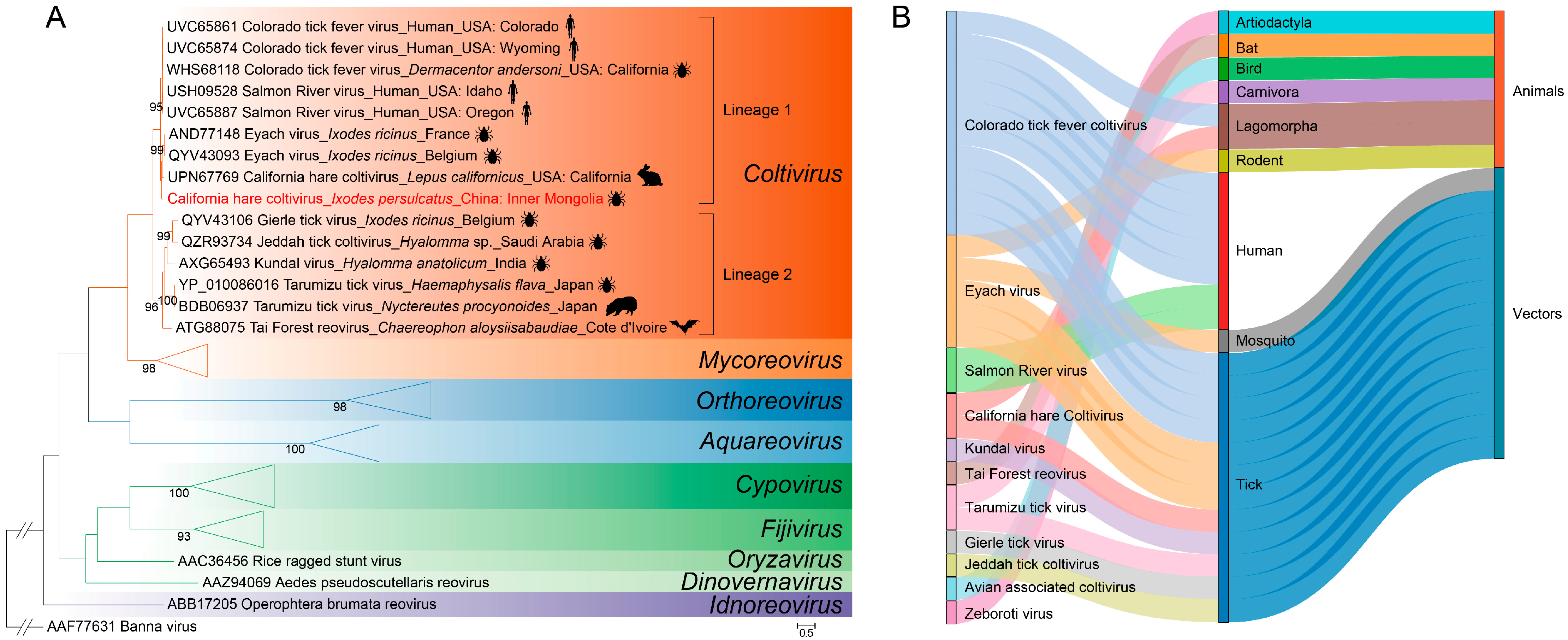
3.2. Phylogenetic Analysis of CHCV Sequences
3.3. CHCV Screening in Ticks by Real-Time RT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hughes, H.R.; Velez, J.O.; Fitzpatrick, K.; Davis, E.H.; Russell, B.J.; Lambert, A.J.; Staples, J.E.; Brault, A.C. Genomic Evaluation of the Genus Coltivirus Indicates Genetic Diversity among Colorado Tick Fever Virus Strains and Demarcation of a New Species. Diseases 2021, 9, 92. [Google Scholar] [CrossRef] [PubMed]
- Florio, L.; Stewart, M.O.; Mugrage, E.R. The etiology of Colorado tick fever. J. Exp. Med. 1946, 83, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Florio, L.; Miller, M.S.; Mugrage, E.R. Colorado tick fever; isolation of the virus from Dermacentar andersoni in Nature and a laboratory study of the transmission of the virus in the tick. J. Immunol. 1950, 64, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Rehse-Kupper, B.; Casals, J.; Rehse, E.; Ackermann, R. Eyach—An arthropod-borne virus related to Colorado tick fever virus in the Federal Republic of Germany. Acta Virol. 1976, 20, 339–342. [Google Scholar] [PubMed]
- Chastel, C.; Main, A.J.; Couatarmanac’h, A.; Le Lay, G.; Knudson, D.L.; Quillien, M.C.; Beaucournu, J.C. Isolation of Eyach virus (Reoviridae, Colorado tick fever group) from Ixodes ricinus and I. ventalloi ticks in France. Arch. Virol. 1984, 82, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.D.; Whitmer, S.L.M.; Sarkale, P.; Fei Fan Ng, T.; Goldsmith, C.S.; Nyayanit, D.A.; Esona, M.D.; Shrivastava-Ranjan, P.; Lakra, R.; Pardeshi, P.; et al. Characterization of Novel Reoviruses Wad Medani Virus (Orbivirus) and Kundal Virus (Coltivirus) Collected from Hyalomma anatolicum Ticks in India during Surveillance for Crimean Congo Hemorrhagic Fever. J. Virol. 2019, 93, 13. [Google Scholar] [CrossRef] [PubMed]
- Fujita, R.; Ejiri, H.; Lim, C.K.; Noda, S.; Yamauchi, T.; Watanabe, M.; Kobayashi, D.; Takayama-Ito, M.; Murota, K.; Posadas-Herrera, G.; et al. Isolation and characterization of Tarumizu tick virus: A new coltivirus from Haemaphysalis flava ticks in Japan. Virus Res. 2017, 242, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.; Dabrowski, P.W.; Kurth, A.; Leendertz, S.A.J.; Leendertz, F.H. A novel Coltivirus-related virus isolated from free-tailed bats from Cote d’Ivoire is able to infect human cells in vitro. Virol. J. 2017, 14, 181. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Li, S.S.; Liu, J.Z. Illustrated keys to families and genera of the superfamily Ixodoidea under new taxonomic system. Zhongguo Ji Sheng Chong Xue Yu Ji Sheng Chong Bing Za Zhi 2011, 29, 302–304, 309. [Google Scholar] [PubMed]
- Teng, K.F.; Jiang, Z.J. Economic Insect Fauna of China, Fasc 39 Acari: Ixodidae; Science Press: Beijing, China, 1991. (In Chinese) [Google Scholar]
- Mangold, A.J.; Bargues, M.D.; Mas-Coma, S. Mitochondrial 16S rDNA sequences and phylogenetic relationships of species of Rhipicephalus and other tick genera among Metastriata (Acari: Ixodidae). Parasitol. Res. 1998, 84, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.A.; Li, H.; Jiang, F.C.; Zhu, F.; Zhang, Y.F.; Chen, J.J.; Tan, C.W.; Anderson, D.E.; Fan, H.; Dong, L.Y.; et al. A Zoonotic Henipavirus in Febrile Patients in China. N. Engl. J. Med. 2022, 387, 470–472. [Google Scholar] [CrossRef] [PubMed]
- Lole, K.S.; Bollinger, R.C.; Paranjape, R.S.; Gadkari, D.; Kulkarni, S.S.; Novak, N.G.; Ingersoll, R.; Sheppard, H.W.; Ray, S.C. Full-length human immunodeficiency virus type 1 genomes from subtype C-infected seroconverters in India, with evidence of intersubtype recombination. J. Virol. 1999, 73, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [PubMed]
- Málková, D.; Holubová, J.; Kolman, J.M.; Marhoul, Z.; Hanzal, F.; Kulková, H.; Markvart, K.; Simková, L. Antibodies against some arboviruses in persons with various neuropathies. Acta Virol. 1980, 24, 298. [Google Scholar] [PubMed]
- Wang, S.S.; Liu, J.Y.; Wang, B.Y.; Wang, W.J.; Cui, X.M.; Jiang, J.F.; Sun, Y.; Guo, W.B.; Pan, Y.S.; Zhou, Y.H.; et al. Geographical distribution of Ixodes persulcatus and associated pathogens: Analysis of integrated data from a China field survey and global published data. One Health 2023, 16, 100508. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tick Species | Inner Mongolia | Heilongjiang | Jilin | Liaoning | Total |
|---|---|---|---|---|---|
| NPP/NTP (NTT) | |||||
| Ixodes persulcatus | 10/59 (311) | 2/100 (104) | 1/41 (180) | nd | 13/200 (595) |
| Devmacentor silvarum | 0/48 (48) | nd | nd | nd | 0/48 (48) |
| Haemaphysalis concinna | 0/48 (48) | nd | nd | nd | 0/48 (48) |
| Haemaphysalis longicornis | nd | nd | nd | 0/67 (181) | 0/67 (181) |
| Total | 10/155 (407) | 2/100 (104) | 1/41 (180) | 0/67 (181) | 13/363 (872) |
| Prevalence (%, 95% CI) | |||||
| Ixodes persulcatus | 3.62 (1.85–6.46) | 1.94 (0.35–6.23) | 0.55 (0.03–2.65) | nd | 2.33 (1.30–3.88) |
| Devmacentor silvarum | 0 | nd | nd | nd | 0 |
| Haemaphysalis concinna | 0 | nd | nd | nd | 0 |
| Haemaphysalis longicornis | nd | nd | nd | 0 | 0 |
| Total | 2.69 (1.36–4.81) | 1.94 (0.35–6.23) | 0.55 (0.03–2.65) | 0 | 1.56 (0.87–2.59) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Z.; Zhang, J.; Liu, Y.; Liu, L.; Tang, F.; Si, G.; Zhang, M.; Li, S.; Zhang, Y.; Peng, C.; et al. First Genomic Evidence of California Hare Coltivirus from Natural Populations of Ixodes persulcatus Ticks in Northeast China. Pathogens 2024, 13, 614. https://doi.org/10.3390/pathogens13080614
Hu Z, Zhang J, Liu Y, Liu L, Tang F, Si G, Zhang M, Li S, Zhang Y, Peng C, et al. First Genomic Evidence of California Hare Coltivirus from Natural Populations of Ixodes persulcatus Ticks in Northeast China. Pathogens. 2024; 13(8):614. https://doi.org/10.3390/pathogens13080614
Chicago/Turabian StyleHu, Zhenyu, Jingtao Zhang, Yantao Liu, Liming Liu, Fang Tang, Guangqian Si, Meiqi Zhang, Shuang Li, Yunfa Zhang, Cong Peng, and et al. 2024. "First Genomic Evidence of California Hare Coltivirus from Natural Populations of Ixodes persulcatus Ticks in Northeast China" Pathogens 13, no. 8: 614. https://doi.org/10.3390/pathogens13080614
APA StyleHu, Z., Zhang, J., Liu, Y., Liu, L., Tang, F., Si, G., Zhang, M., Li, S., Zhang, Y., Peng, C., Zhang, L., Ma, X., Zhang, X., & Liu, W. (2024). First Genomic Evidence of California Hare Coltivirus from Natural Populations of Ixodes persulcatus Ticks in Northeast China. Pathogens, 13(8), 614. https://doi.org/10.3390/pathogens13080614