Characterizing Virulence of the Pyrenophora tritici-repentis Isolates Lacking Both ToxA and ToxB Genes



Abstract
:1. Introduction
2. Results
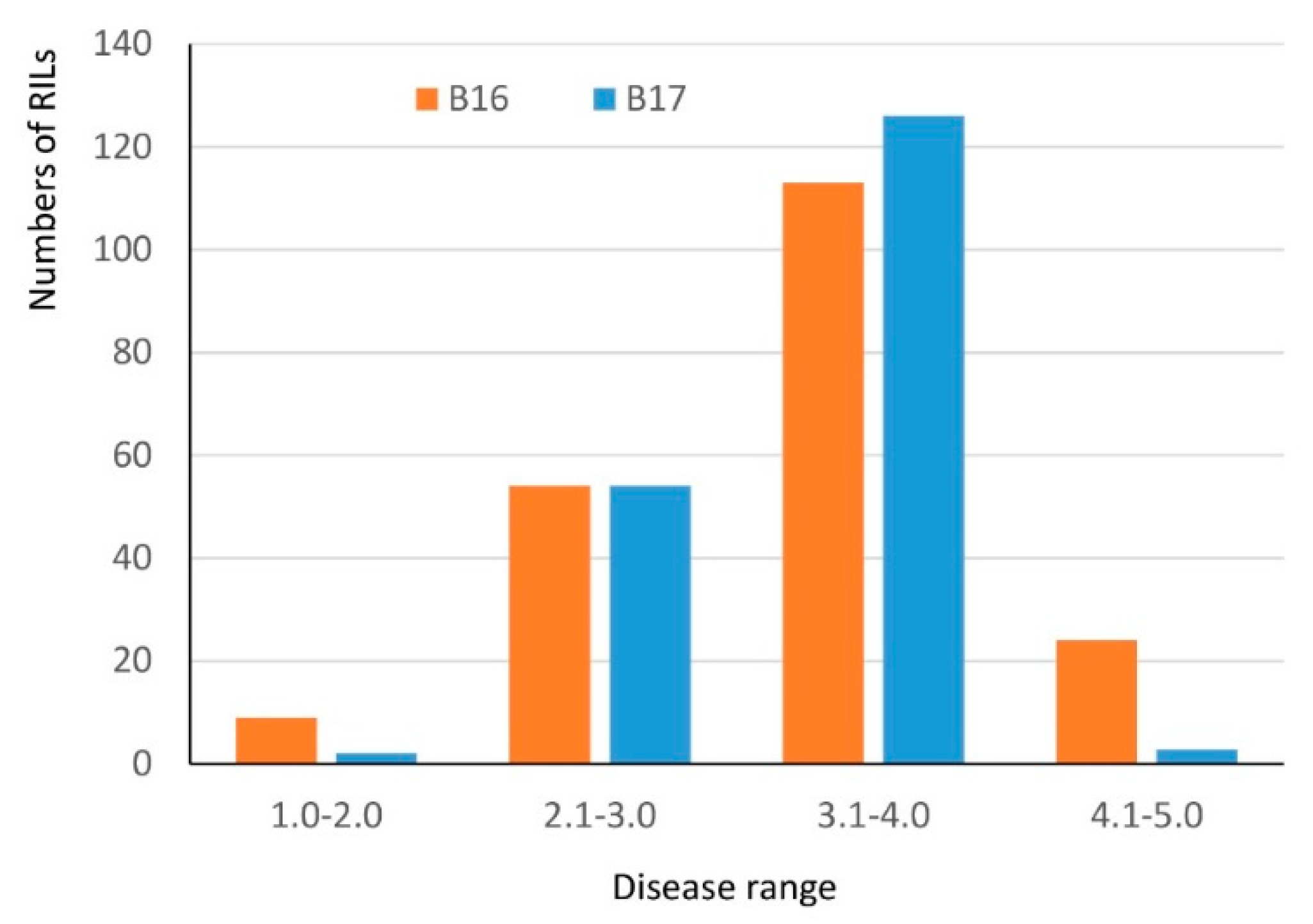
2.1. Pathogenicity Test
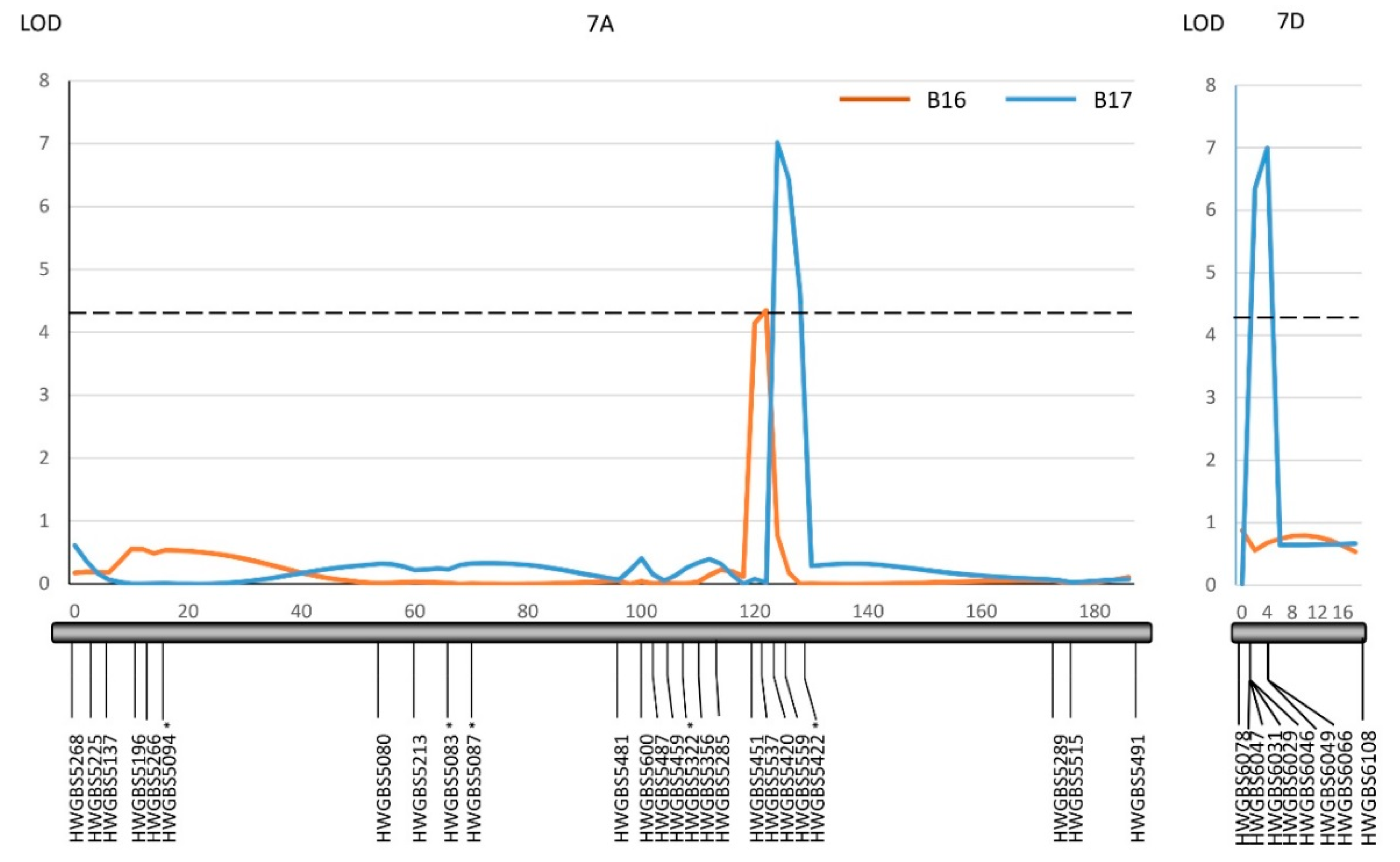
2.2. QTL Mapping
3. Discussion
4. Materials and Methods
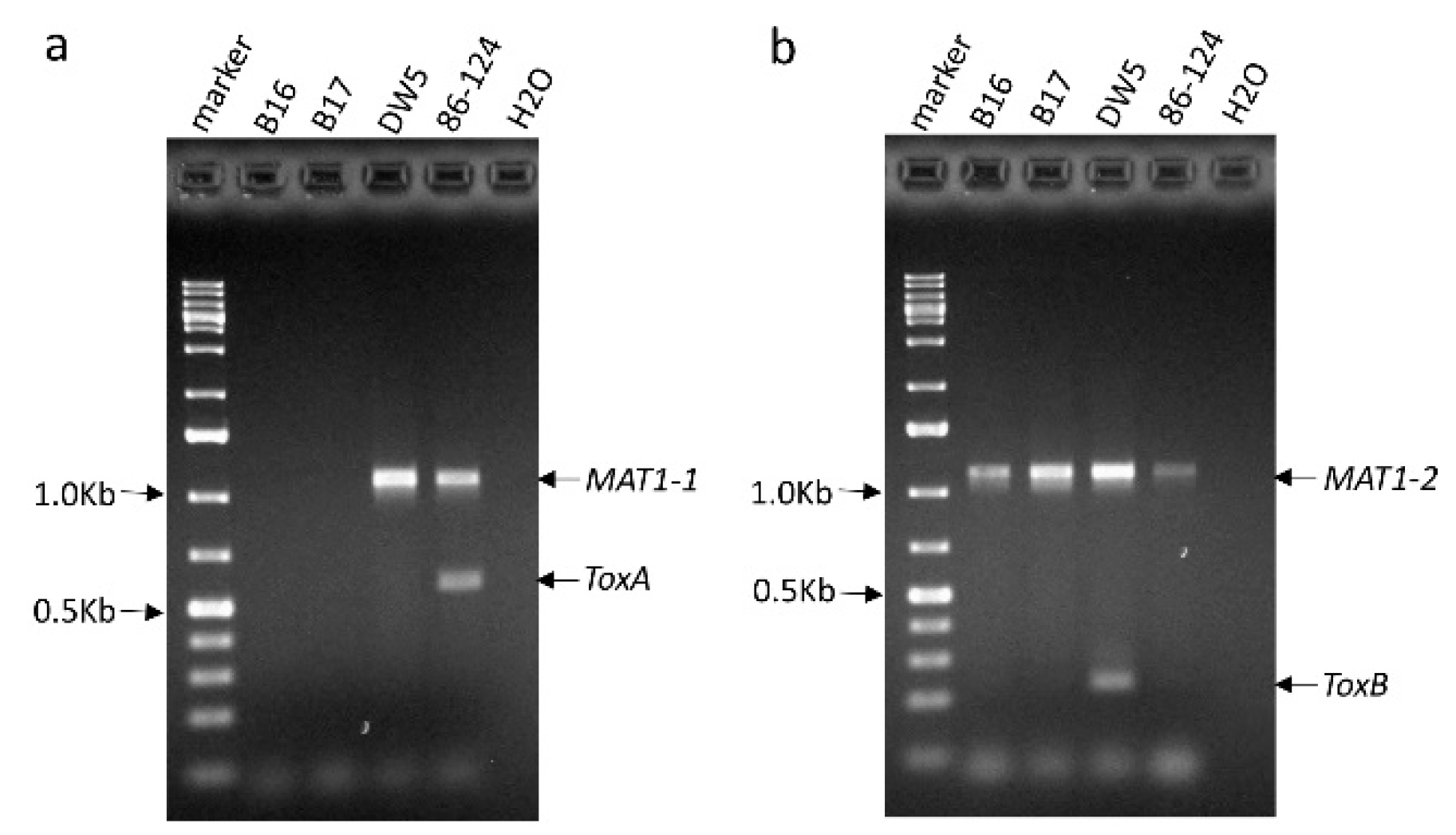
4.1. Fungal Isolates and PCR Testings
4.2. Plant Materials, Experimental Designs, and Disease Evaluations
4.3. Statistic Analysis and QTL Mapping
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Faris, J.D.; Liu, Z.; Xu, S.S. Genetics of tan spot resistance in wheat. Theor. Appl. Genet. 2013, 126, 2197–2217. [Google Scholar] [CrossRef] [PubMed]
- Friskop, A.; Liu, Z. Fungal Leaf Spot Diseases of Wheat: Tan Spot, Stagonospora nodorum Blotch and Septoria tritici Blotch; North Dakota State University Extension Service: Fargo, ND, USA, 2016; p. 1249. [Google Scholar]
- Lamari, L.; Bernier, C.C. Evaluation of wheat lines and cultivars to tan spot Pyrenophora tritici-repentis based on lesion type. Can. J. Plant Pathol. 1989, 11, 49–56. [Google Scholar] [CrossRef]
- Lamari, L.; Bernier, C.C. Genetics of tan necrosis and extensive chlorosis in tan spot of wheat caused by Pyrenophora tritici-repentis. Phytopathology 1991, 81, 1092–1095. [Google Scholar] [CrossRef]
- Lamari, L.; Strelkov, S.E.; Yahyaoui, A.; Orabi, J.; Smith, R.B. The identification of two new races of Pyrenophora tritici-repentis from the host center of diversity confirms a one-to-one relationship in tan spot of wheat. Phytopathology 2003, 93, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Ciuffetti, L.M.; Francl, L.J.; Ballance, G.M.; Bockus, W.W.; Lamari, L.; Meinhardt, S.W.; Rasmussen, J.B. Standardization of toxin nomenclature in the Pyrenophora tritici-repentis/wheat interaction. Can. J. Plant Pathol. 1998, 20, 421–424. [Google Scholar] [CrossRef]
- Ciuffetti, L.M.; Manning, V.A.; Pandelova, I.; Betts, M.F.; Martinez, J.P. Host-selective toxins, Ptr ToxA and Ptr ToxB, as necrotrophic effectors in the Pyrenophora tritici-repentis–wheat interaction. New Phytol. 2010, 187, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Strelkov, S.E.; Lamari, L. Host–parasite interactions in tan spot Pyrenophora tritici-repentis of wheat. Can. J. Plant Pathol. 2003, 25, 339–349. [Google Scholar] [CrossRef]
- Ali, S.; Francl, L.J. Population race structure of Pyrenophora tritici-repentis prevalent on wheat and noncereal grasses in the Great Plains. Plant Dis. 2003, 87, 418–422. [Google Scholar] [CrossRef]
- Ali, S.; Francl, L.J. Identification and characterization of novel isolates of Pyrenophora tritici-repentis from Akansas. Plant Dis. 2010, 94, 229–235. [Google Scholar] [CrossRef]
- Meinhardt, S.W.; Ali, S.; Ling, H.; Francl, L.J. A new race of Pyrenophora tritici-repentis that produces a putative host-selective toxin. In Proceedings of the Fourth International Wheat Tan Spot and Spot Blotch Workshop, Bemidji, MN, USA, 21–24 July 2002; Rasmussen, J.B., Friesen, T.L., Ali, S., Eds.; North Dakota Agricultural Experiment Station: Fargo, ND, USA, 2003; pp. 117–119. [Google Scholar]
- Ciuffetti, L.M.; Manning, V.A.; Martinez, J.P.; Pandelova, I.; Andrie, R.M. Proteinaceous toxins of Pyrenophora tritici-repentis and investigation of the site-of-action of Ptr ToxA. In Proceedings of the Fourth International Wheat Tan Spot and Spot Blotch Workshop, Bemidji, MN, USA, 21–24 July 2002; Rasmussen, J.B., Friesen, T.L., Ali, S., Eds.; North Dakota Agricultural Experiment Station: Fargo, ND, USA, 2003; pp. 96–102. [Google Scholar]
- Ameen, G.; Kariyawasam, G.; Shi, G.; Friesen, T.L.; Faris, J.D.; Ali, S.; Liu, Z. Molecular manipulation of the mating-type system and development of a new approach for characterizing pathogen virulence in Pyrenophora tritici-repentis. Fungal Genet. Biol. 2017, 109, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Virdi, S.; Liu, Z.; Overlander, M.; Zhang, Z.; Xu, S.S.; Friesen, T.; Faris, J.D. New insights into the roles of host gene-necrotrophic effector interactions in governing susceptibility of durum wheat to tan spot and Septoria nodorum blotch. G3 Genes Genomes Genet. 2016, 8, 3–116. [Google Scholar] [CrossRef] [PubMed]
- Kariyawasam, G.K.; Carter, A.H.; Rasmussen, J.B.; Faris, J.; Xu, S.S.; Mergoum, M.; Liu, Z. Genetic relationships between race-nonspecific and race-specific interactions in the wheat—Pyrenophora tritici-repentis pathosystem. Theor. Appl. Genet. 2016, 129, 897–908. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zurn, J.D.; Kariyawasam, G.; Faris, J.D.; Shi, G.; Hansen, J.; Acevedo, M. Inverse gene-for-gene interactions contribute additively to tan spot susceptibility in wheat. Theor. Appl. Genet. 2017, 130, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Friesen, T.L.; Ali, S.; Kianian, S.; Francl, L.J.; Rasmussen, J.B. Role of host sensitivity to Ptr ToxA in development of tan spot of wheat. Phytopathology 2003, 93, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Gamba, F.M.; Lamari, L. Mendelian inheritance of resistance to tan spot Pyrenophora tritici-repentis in selected genotypes of durum wheat (Triticum turgidum). Can. J. Plant Pathol. 1998, 20, 408–414. [Google Scholar] [CrossRef]
- Singh, P.K.; Mergoum, M.; Ali, S.; Adhikari, T.B.; Hughes, G.R. Genetic analysis of resistance to Pyrenophora tritici-repentis races 1 and 5 in tetraploid and hexaploid wheat. Phytopathology 2008, 98, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Mergoum, M.; Adhikari, T.B.; Shah, T.; Ghavami, F.; Kianian, S.F. Genetic and molecular analysis of wheat tan spot resistance effective against Pyrenophora tritici-repentis races 2 and 5. Mol. Breed. 2010, 25, 369–379. [Google Scholar] [CrossRef]
- See, P.T.; Marathamuthu, K.A.; Iagallo, E.M.; Oliver, R.P.; Moffat, C.S. Evaluating the importance of the tan spot ToxA–Tsn1 interaction in Australian wheat varieties. Plant Pathol. 2018, 67, 1066–1075. [Google Scholar] [CrossRef]
- Moreno, M.V.; Stenglein, S.; Perelló, A.E. Distribution of races and Tox genes in Pyrenophora tritici-repentis isolates from wheat in Argentina. Trop. Plant Pathol. 2015, 40, 141–146. [Google Scholar] [CrossRef]
- Abdullah, S.; Sehgal, S.K.; Ali, S.; Liatukas, Z.; Ittu, M.; Kaur, N. Characterization of Pyrenophora tritici-repentis (tan spot of wheat) races in Baltic States and Romania. Plant Pathol. J. 2017, 33, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, S.; Sehgal, S.K.; Ali, S. Race diversity of Pyrenophora tritici-repentis in South Dakota and response of predominant wheat cultivars to tan spot. J. Plant Pathol. Microbiol. 2017, 8, 409. [Google Scholar] [CrossRef]
- Gurung, S.; Mamidi, S.; Bonman, J.M.; Jackson, E.W.; Del Rio, L.E.; Acevedo, M.; Adhikari, T.B. Identification of novel genomic regions associated with resistance to Pyrenophora tritici-repentis races 1 and 5 in spring wheat landraces using association analysis. Theor. Appl. Genet. 2011, 123, 1029. [Google Scholar] [CrossRef] [PubMed]
- Faris, J.D.; Abeysekara, N.S.; McClean, P.E.; Xu, S.S.; Friesen, T.L. Tan spot susceptibility governed by the Tsn1 locus and race-nonspecific resistance quantitative trait loci in a population derived from the wheat lines Salamouni and Katepwa. Mol. Breed. 2012, 30, 1669–1678. [Google Scholar] [CrossRef]
- Chu, C.G.; Chao, S.; Friesen, T.L.; Faris, J.D.; Zhong, S.; Xu, S.S. Identification of novel tan spot resistance QTLs using an SSR-based linkage map of tetraploid wheat. Mol. Breed. 2010, 25, 327–338. [Google Scholar] [CrossRef]
- Liu, Z.; El-Basyoni, I.; Kariyawasam, G.; Zhang, G.; Fritz, A.; Hansen, J.; Baenziger, P.S. Evaluation and association mapping of resistance to tan spot and Stagonospora nodorum blotch in adapted winter wheat germplasm. Plant Dis. 2015, 99, 1333–1341. [Google Scholar] [CrossRef]
- Manning, V.A.; Ciuffetti, L.M. Necrotrophic effector epistasis in the Pyrenophora tritici-repentis-wheat interaction. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Shjerve, R.A.; Faris, J.D.; Brueggeman, R.S.; Yan, C.; Zhu, Y.; Koladia, V.; Friesen, T.L. Evaluation of a Pyrenophora teres f. teres mapping population reveals multiple independent interactions with the barley 6H chromosome region. Fungal Genet. Biol. 2014, 70, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Lepoint, P.; Renard, M.E.; Legrève, A.; Duveiller, E.; Maraite, H. Genetic diversity of the mating type and toxin production genes in Pyrenophora tritici-repentis. Phytopathology 2010, 100, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Andrie, R.M.; Pandelova, I.; Ciuffetti, L.M. A combination of phenotypic and genotypic characterization strengthens Pyrenophora tritici-repentis race identification. Phytopathology 2007, 97, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Hussain, W.; Baenziger, P.S.; Belamkar, V.; Guttieri, M.J.; Venegas, J.P.; Easterly, A.; Sallam, A.; Poland, J. Genotyping-by-sequencing derived high-density linkage map and its application to QTL mapping of flag leaf traits in bread wheat. Sci. Rep. 2017, 7, 16394. [Google Scholar] [CrossRef] [PubMed]
- Joehanes, R.; Nelson, J.C. QGene 4.0, An extensible Java QTL-analysis platform. Bioinformatics 2008, 24, 2788–2789. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes | Wheat Type | B16 a | B17 a | ||
|---|---|---|---|---|---|
| Disease Mean | Rank Mean | Disease Mean | Rank Mean | ||
| Rollag | Common wheat | 1.17 | 6.17O | 1.17 | 9.83JK |
| Velva | Common wheat | 1.17 | 6.17O | 1.50 | 17.50JK |
| Jenna | Common wheat | 1.50 | 11.50NO | 1.50 | 17.50JK |
| RB07 | Common wheat | 1.50 | 14.50MNO | 1.50 | 17.50JK |
| Brennan | Common wheat | 1.67 | 17.17MNO | 1.67 | 21.33IJK |
| Glenn | Common wheat | 1.83 | 25.33KLMN | 1.50 | 18.33IJK |
| Briggs | Common wheat | 2.00 | 28.50JKLM | 2.33 | 38.17GH |
| Chinese Spring | Common wheat | 2.00 | 28.50JKLM | 2.50 | 23.00FG |
| Prosper | Common wheat | 2.00 | 28.50JKLM | 1.33 | 13.67JK |
| Vantage | Common wheat | 2.00 | 28.50JKLM | 2.17 | 33.67GHI |
| Wesley | Common wheat | 2.00 | 28.50JKLM | 2.00 | 29.33HIJ |
| Howard | Common wheat | 2.17 | 33.67IJKL | 2.17 | 33.67GHI |
| Kelby | Common wheat | 2.50 | 42.83HIJ | 2.50 | 43.00FG |
| Select | Common wheat | 2.50 | 42.83HIJ | 2.50 | 43.00FG |
| SY Soren | Common wheat | 2.67 | 46.83GHI | 2.67 | 47.50EFG |
| SY Tyra | Common wheat | 3.00 | 56.00FGH | 3.50 | 67.50CD |
| Mott | Common wheat | 4.00 | 81.50BCD | 4.00 | 80.00BC |
| Steele-ND | Common wheat | 4.00 | 81.50BCD | 4.00 | 80.00BC |
| Harry | Common wheat | 4.00 | 81.50BCD | 4.50 | 96.00A |
| Barlow | Common wheat | 4.1 | 86.67ABC | 4.33 | 90.67AB |
| Alkabo | Durum wheat | 2.17 | 33.67IJK | 2.67 | 47.50EFG |
| Lebsock | Durum wheat | 3.00 | 56.00FGH | 3.50 | 66.00CD |
| Grenora | Durum wheat | 3.00 | 56.00FGH | 3.17 | 59.67DE |
| Carpio | Durum wheat | 3.1 | 60.00EFG | 3.17 | 59.67DE |
| Tioga | Durum wheat | 3.5 | 68.50DEF | 3.50 | 66.00CD |
| Ben | Durum wheat | 3.8 | 77.00BCD | 4.00 | 80.00BC |
| Dilse | Durum wheat | 4.00 | 81.50BCD | 4.00 | 80.00BC |
| Divide | Durum wheat | 4.00 | 81.50BCD | 4.00 | 80.00BC |
| Langdon | Durum wheat | 4.33 | 91.83AB | 4.50 | 96.00A |
| Rusty | Durum wheat | 4.33 | 91.83AB | 4.33 | 90.67AB |
| Mountrail | Durum wheat | 4.50 | 97.00A | 4.50 | 96.00A |
| Pierce | Durum wheat | 4.50 | 97.00A | 4.50 | 96.00A |
| Least Significant Difference | 15.0 | 15.70 | |||
| QTL | Interval (cM) | Flanking Markers | B16 a | B17 a | Source b | ||
|---|---|---|---|---|---|---|---|
| LOD | R2 | LOD | R2 | ||||
| QTs.zhl.-7A-1 | 120.0-122.0 | HWGBS5451-HWGBS5420 | 4.4 | 0.08 | - | - | W |
| QTs.zhl.-7A-2 | 124.0-130.0 | HWGBS5420-HWGBS5422 | - | - | 7.0 | 0.11 | W |
| QTs.zhl.-7D | 176.0-180.0 | HWGBS6047-HWGBS6066 | - | - | 7.0 | 0.09 | W |
| Primers | Sequences (from 5′ to 3′) | Purpose | Reference |
|---|---|---|---|
| PtrPLP7 | GCTTTACTACAACTTTCCTCTACC | Amplify the MAT1-2 | Lepoint et al. (2010) [31] |
| PtrPLP10 | GTACGGGCCAGCATGACGTGC | ||
| PtrPLP2 | CAGAACAAAGGCAGGACTGTGAGC | Amplify the MAT1-1 | Lepoint et al. (2010) [31] |
| PtrPLP4 | ATGCGCTCAGCAAGGAAGGTCG | ||
| TA51F | GCGTTCTATCCTCGTACTTC | Amplify the ToxA | Andrie et al. (2007) [32] |
| TA52R | GCATTCTCCAATTTTCACG | ||
| TB71F | GCTACTTGCTGTGGCTATC | Amplify the ToxB | Andrie et al. (2007) [32] |
| TB6R | ACGTCCTCCACTTTGCACACTCTC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, J.; Shi, G.; Liu, Z. Characterizing Virulence of the Pyrenophora tritici-repentis Isolates Lacking Both ToxA and ToxB Genes. Pathogens 2018, 7, 74. https://doi.org/10.3390/pathogens7030074
Guo J, Shi G, Liu Z. Characterizing Virulence of the Pyrenophora tritici-repentis Isolates Lacking Both ToxA and ToxB Genes. Pathogens. 2018; 7(3):74. https://doi.org/10.3390/pathogens7030074
Chicago/Turabian StyleGuo, Jingwei, Gongjun Shi, and Zhaohui Liu. 2018. "Characterizing Virulence of the Pyrenophora tritici-repentis Isolates Lacking Both ToxA and ToxB Genes" Pathogens 7, no. 3: 74. https://doi.org/10.3390/pathogens7030074
APA StyleGuo, J., Shi, G., & Liu, Z. (2018). Characterizing Virulence of the Pyrenophora tritici-repentis Isolates Lacking Both ToxA and ToxB Genes. Pathogens, 7(3), 74. https://doi.org/10.3390/pathogens7030074