Investigation of Camphor Effects on Fusarium graminearum and F. culmorum at Different Molecular Levels

 and
and
Abstract
:1. Introduction
2. Results
2.1. Antifungal Application Analysis
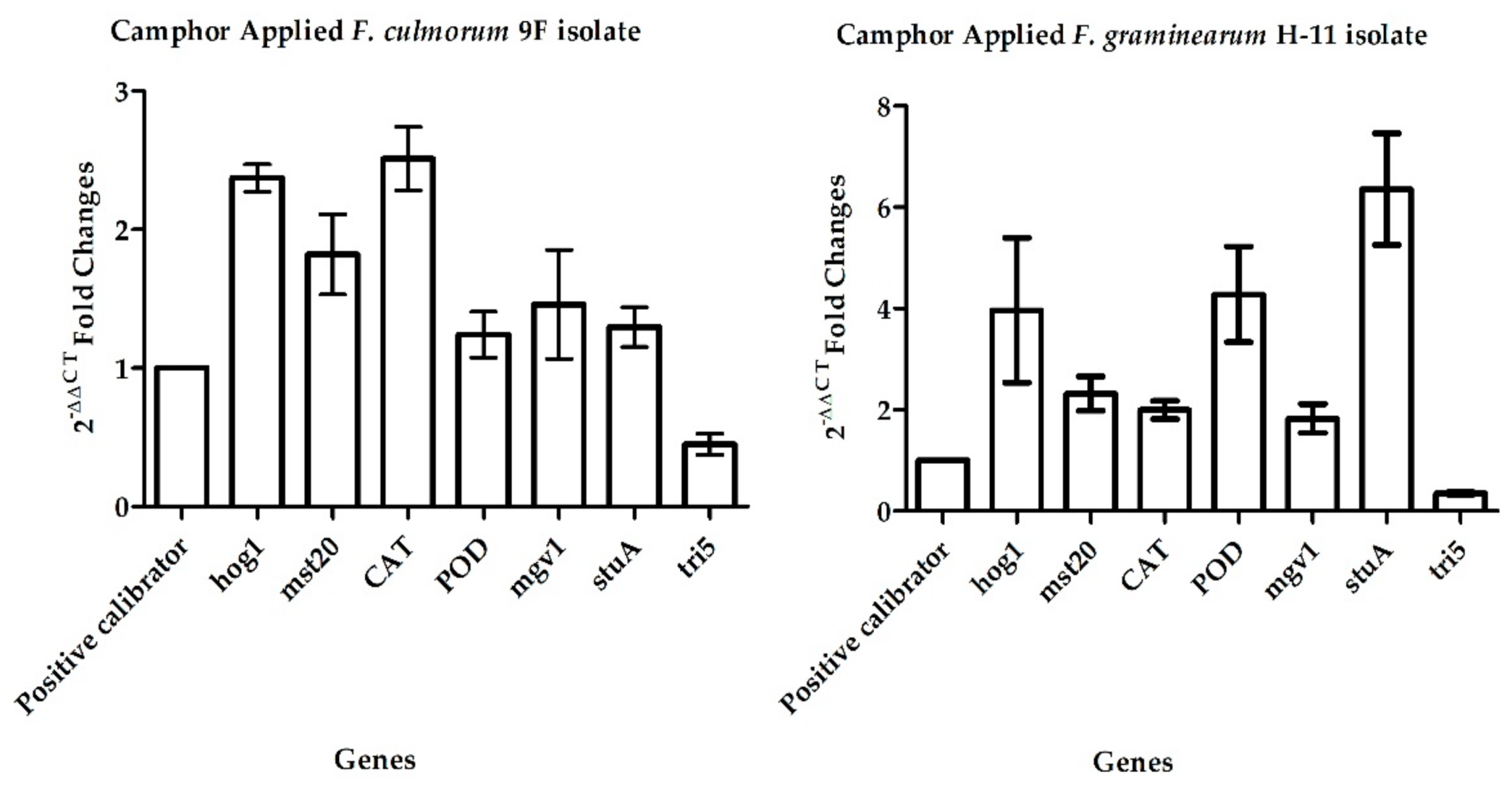
2.2. Gene Expression Analysis
2.3. Staining Analysis
2.4. Antioxidant Activities and Thin-Layer Chromatography (TLC)
2.5. Coupled Digestion-Random Amplification (CRED-RA) and Random Amplified Polymorphic DNA (RAPD) Analyses
3. Discussion
4. Materials and Methods
4.1. Antifungal Treatment
4.2. Total RNA Extraction, cDNA Synthesis, and Gene Expression Assays
4.3. Fluorescence Microscopy Assays
4.4. Antioxidant Activity and TLC
4.5. CRED-RA Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miedaner, T.; Cumagun, C.J.R.; Chakraborty, S. Population genetics of three important head blight pathogens Fusarium graminearum, F. pseudograminearum and F. culmorum. J. Phytopathol. 2008, 156, 129–139. [Google Scholar] [CrossRef]
- Matny, O.N. Fusarium head blight and crown rot on wheat & barley: Losses and health risks. APAR 2015, 2, 38–43. [Google Scholar] [CrossRef]
- Pasquali, M.; Migheli, Q. Genetic approaches to chemotype determination in type B-trichothecene producing Fusaria. Int. J. Food Microbiol. 2014, 189, 164–182. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.S.; Kistler, H.C. Heading for disaster: Fusarium graminearum on cereal crop. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Tunali, B.; Özseven, İ.; Büyük, O.; Erdurmuş, D.; Demirci, A. Fusarium head blight and deoxynivalenol accumulation of wheat in Marmara region and reactions of wheat cultivars and lines to F. graminearum and Fusarium culmorum. Plant Pathol. J. 2006, 2, 150–156. [Google Scholar]
- Desjardins, A.E.; Proctor, R.H. Molecular biology of Fusarium mycotoxins. Int. J. Food Microbiol. 2007, 119, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Kerényi, Z.; Moretti, A.; Waalwijk, C.; Olah, B.; Hornok, L. Mating type sequences in asexually reproducing Fusarium species. Appl. Environ. Microbiol. 2004, 70, 4419–4423. [Google Scholar] [CrossRef] [PubMed]
- Przemieniecki, S.W.; Kurowski, T.P.; Korzekwa, K. Chemotypes and geographic distribution of the Fusarium graminearum species complex. Environ. Biotechnol. 2015, 10, 45–59. [Google Scholar] [CrossRef]
- Trail, F. For blighted waves of grain: Fusarium graminearum in the postgenomics era. Plant Physiol. 2009, 149, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Obanor, F.; Erginbas-Orakci, G.; Tunali, B.; Nicol, J.M.; Chakraborty, S. Fusarium culmorum is a single phylogenetic species based on multilocus sequence analysis. Fungal Biol. 2010, 114, 753–765. [Google Scholar] [CrossRef] [PubMed]
- Dal Bello, G.M.; Monaco, C.I.; Simon, M.R. Biological control of seedling blight of wheat caused by Fusarium graminearum with beneficial rhizosphere microorganisms. World J. Microbiol. Biotechnol. 2002, 18, 627–636. [Google Scholar] [CrossRef]
- Anand, A.; Zhou, T.; Trick, H.N.; Gill, B.S.; Bockus, W.W.; Muthukrishnan, S. Greenhouse and field testing of transgenic wheat plants stably expressing genes for thaumatin-like protein, chitinase and glucanase against Fusarium graminearum. J. Exp. Bot. 2003, 54, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- Bai, G.; Shaner, G. Management and resistance in wheat and barley to Fusarium head blight. Annu. Rev. Phytopathol. 2004, 42, 135–161. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.H.; Ishii, H.; Nishimura, K.; Ohshima, M.; Iwama, T.; Yoshimatsu, H. Genetic analysis and PCR-based identification of major Fusarium species causing head blight on wheat in Japan. JGPP 2008, 10, 110–118. [Google Scholar] [CrossRef]
- Talas, F.; McDonald, B.A. Significant variation in sensitivity to a DMI fungicide in field populations of F usarium graminearum. Plant Pathol. 2015, 64, 664–670. [Google Scholar] [CrossRef]
- Yang, Y.; Li, M.; Duan, Y.; Li, T.; Shi, Y.; Zhao, D.; Zhou, Z. A new point mutation in β2 -tubulin confers resistance to carbendazim in Fusarium asiaticum. Pest. Biochem. Physiol. 2018, 145, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Arif, T.; Bhosale, J.D.; Kumar, N.; Mandal, T.K.; Bendre, R.S.; Lavekar, G.S.; Dabur, R. Natural products–antifungal agents derived from plants. J. Asian Nat. Prod. Res. 2009, 11, 621–638. [Google Scholar] [CrossRef] [PubMed]
- Sefer, Ö.; Yörük, E.; Develi, E.S.; Sezer, A.S.; Konukcu, Z. 2,4-dimetilpirol’ün başak yanikliği hastaliği etmeni Fusarium culmorum’un üzerine etkilerinin incelenmesi. IJSM 2017, 4, 103–113. [Google Scholar] [CrossRef]
- Yörük, E. Tetraconazole leads to alterations in Fusarium graminearum at different molecular levels. Appl. Ecol. Environ. 2018, 16, 6155–6167. [Google Scholar] [CrossRef]
- Green, B.G. Sensory characteristics of camphor. J. Investig. Dermatol. 1990, 94, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Irzykowska, L.; Bocianowski, J.; Baturo-Cieśniewska, A. Association of mating-type with mycelium growth rate and genetic variability of Fusarium culmorum. Cent. Eur. J. Biol. 2013, 8, 701–711. [Google Scholar] [CrossRef]
- Ponts, N.; Couedelo, L.; Pinson-Gadais, L.; Verdal-Bonnin, M.N.; Barreau, C.; Richard-Forget, F. Fusarium response to oxidative stress by H2O2 is trichothecene chemotype-dependent. FEMS Microbiol. Lett. 2009, 293, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, A.; Bai, G.; Guo, P.; Xiao, K.; Guenzi, A.C.; Ayoubi, P. Fusarium graminearum-induced changes in gene expression between Fusarium head blight-resistant and susceptible wheat cultivars. Funct. Integr. Genom. 2007, 7, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Scherm, B.; Orru, M.; Balmas, V.; Spanu, F.; Azara, E.; Delogu, G.; Hammond, T.M.; Keller, N.P.; Migheli, Q. Altered trichothecene biosynthesis in TRI6-silenced transformants of Fusarium culmorum influences the severity of crown and foot rot on durum wheat seedlings. Mol. Plant Pathol. 2011, 12, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Kazan, K.; Gardiner, D.M.; Manners, J.M. On the trail of a cereal killer: Recent advances in Fusarium graminearum pathogenomics and host resistance. Mol. Plant Pathol. 2012, 13, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Yaguchi, A.; Yoshinari, T.; Tsuyuki, R.; Takahashi, H.; Nakajima, T.; Sugita-Konishi, Y.; Nagasawa, H.; Sakuda, S. Isolation and identification of precocenes and piperitone from essential oils as specific inhibitors of trichothecene production by Fusarium graminearum. J. Agric. Food Chem. 2009, 57, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Yörük, E.; Sefer, Ö.; Sezer, A.S.; Konukcu, Z.; Develi, E.S. Eugenol’ün Fusarium culmorum üzerindeki etkilerinin incelenmesi. FEBED 2018, 8, 215–221. [Google Scholar] [CrossRef]
- Zemek, J.; Košíková, B.; Augustin, J.; Joniak, D. Antibiotic properties of lignin components. Folia Microbiol. 1979, 24, 483–486. [Google Scholar] [CrossRef]
- Boonchird, C.; Flegel, T.W. In vitro antifungal activity of eugenol and vanillin against Candida albicans and Cryptococcus neoformans. Can. J. Microbiol. 1982, 28, 1235–1241. [Google Scholar] [CrossRef] [PubMed]
- Kollu, N.; Baireddy, V.; Sivadevuni, G.; Solipuram, M. Influence of volatile compounds and food preservatives in the management of fumonisins (B1) production by Fusarium moniliforme. Afr. J. Biotechnol. 2009, 8, 4207–4210. [Google Scholar]
- Livak, J.K.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Harris, E.L.V.; Angal, S. Protein Purifications Method: A Practical Approach, 1st ed.; IRL Press: Oxford, UK, 1989; pp. 178–186. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Chem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Cho, U.H.; Park, J.O. Mercury-induced oxidative stress in tomato seedlings. Plant Sci. 2000, 156, 1–9. [Google Scholar] [CrossRef]
- Panda, S.K.; Chaudhury, I.; Khan, M.H. Heavy metals induce lipid peroxidation and affect antioxidants in wheat leaves. Biol. Plant. 2003, 46, 289–294. [Google Scholar] [CrossRef]
- Cvetnic, Z.; Pepeljnjak, S.; Šegvi, M. Toxigenıc potentıal of Fusarium specıes isolated from non-harvested maize. Arh. Hig. Rada. Toksikol. 2005, 56, 275–280. [Google Scholar] [PubMed]
- Niu, C.; Kebede, H.; Auld, D.L.; Woodward, J.E.; Burow, G.; Wright, R.J. A safe inexpensive method to isolate high quality plant and fungal DNA in an open laboratory environment. Afr. J. Biotechnol. 2008, 7, 2818–2822. [Google Scholar]
- Yörük, E.; Tunali, B.; Kansu, B.; Ölmez, F.; Uz, G.; Zümrüt, I.M.; Sarıkaya, A.; Meyva, G. Characterization of high-level deoxynivalenol producer Fusarium graminearum and F. culmorum isolates caused head blight and crown rot diseases in Turkey. JPDP 2016, 123, 177–186. [Google Scholar] [CrossRef]
- Yörük, E.; Albayrak, G. Genetic characterization of Fusarium graminearum and F. culmorum isolates from Turkey by using random-amplified polymorphic DNA. Genet. Mol. Res. 2013, 12, 1360–1372. [Google Scholar] [CrossRef] [PubMed]
- Nardemir, G.; Agar, G.; Arslan, E.; Erturk, F.A. Determination of genetic and epigenetic effects of glyphosate on Triticum aestivum with RAPD and CRED-RA techniques. TxPP 2015, 27, 131–139. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | 2−ΔΔCT for 9F | 2−ΔΔCT for H-11 |
|---|---|---|
| hog1 (DQ065608.1) | 2.178 ± 0.09 | 3.965 ± 1.42 |
| mst20 (XM_011329927.1) | 1.821 ± 0.29 | 2.318 ± 0.33 |
| mgv1 (AF492766.1) | 1.460 ± 0.39 | 1.826 ± 0.28 |
| stuA (HG970332.2) | 1.297 ± 0.14 | 6.359 ± 1.01 |
| CAT (XM_011328072.1) | 2.512 ± 0.22 | 1.996 ± 0.18 |
| POD (XM_011329011.1) | 1.241 ± 0.16 | 4.280 ± 0.94 |
| tri5 (AY130290.1) | 0.449 ± 0.07 | 0.349 ± 0.02 |
| Primer | Sequence (5´–3´) | % GC | Total Band No. |
|---|---|---|---|
| OPA-01 | CAGGCCCTTC | 70% | 13 |
| OPA-03 | AGTCAGCCAC | 60% | 9 |
| OPA-04 | AATCGGGCTG | 60% | - |
| OPA-05 | AGGGGTCTTG | 60% | - |
| OPA-06 | GGTCCCTGAC | 70% | - |
| OPA-07 | GAAACGGGTG | 60% | 9 |
| OPA-08 | GTGACGTAGG | 60% | 7 |
| OPB-06 | TGCTCTGCCC | 70% | 8 |
| OPB-07 | GGTGACGCAG | 70% | 5 |
| OPB-09 | TGGGGGACTC | 70% | 6 |
| OPB-10 | CTGCTGGGAC | 70% | 12 |
| OPB-13 | TTCCCCCGCT | 70% | 8 |
| OPB-14 | TCCGCTCTGG | 70% | 9 |
| OPC-5 | GATGACCGCC | 70% | - |
| OPC-4 | CCGCATCTAC | 60% | - |
| Primer | 9F Control | 9F Experiment Total Band Number | 9F Experiment Total Polymorphic Band Number | 9F Polymorphism (%) | H-11 Control | H-11 Experiment Total Band Number | H-11 Experiment Total Polymorphic Band Number | H-11 Polymorphism (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IC50 | IC50 | IC50 | IC50 | IC50 | IC50 | |||||||||||
| HapII | MspI | HapII | MspI | HapII | MspI | HapII | MspI | HapII | MspI | HapII | MspI | HapII | MspI | HapII | MspI | |
| OPA1 | 3 | 4 | 3 | 3 | 0 | 1 | 0 | 25 | 4 | 5 | 8 | 2 | 5 | 3 | 100 | 60 |
| OPA3 | 5 | 7 | 5 | 6 | 0 | 1 | 0 | 14.28 | 5 | 5 | 5 | 5 | 0 | 0 | 0 | 0 |
| OPA7 | 3 | 3 | 3 | 4 | 0 | 1 | 0 | 33.33 | 3 | 3 | 6 | 3 | 3 | 0 | 100 | 0 |
| OPA8 | 3 | 5 | 2 | 2 | 1 | 3 | 33.33 | 60 | 2 | 4 | 2 | 3 | 0 | 1 | 0 | 25 |
| OPB6 | 5 | 7 | 4 | 5 | 1 | 2 | 20 | 28.57 | 4 | 5 | 5 | 6 | 3 | 1 | 75 | 20 |
| OPB7 | 3 | 4 | 3 | 3 | 0 | 1 | 0 | 25 | 4 | 4 | 4 | 4 | 2 | 2 | 50 | 50 |
| OPB9 | 3 | 3 | 3 | 3 | 0 | 0 | 0 | 0 | 4 | 4 | 4 | 4 | 2 | 2 | 50 | 50 |
| OPB10 | 6 | 7 | 5 | 6 | 1 | 1 | 16.67 | 14.28 | 5 | 5 | 5 | 6 | 4 | 3 | 80 | 60 |
| OPB13 | 5 | 5 | 5 | 5 | 0 | 0 | 0 | 0 | 5 | 5 | 5 | 4 | 0 | 1 | 0 | 20 |
| OPB14 | 3 | 2 | 1 | 2 | 2 | 0 | 66.67 | 0 | 2 | 3 | 4 | 3 | 4 | 4 | 100 | 100 |
| Mean | 3.9 | 4.7 | 3.4 | 3.9 | 0.5 | 1 | 2.857143 | 18.33333 | 3.8 | 4.3 | 4.8 | 4 | 2.3 | 1.7 | 55.5 | 38.5 |
| Primer | Primer Sequence (5′-3′) | Size (bp) |
|---|---|---|
| betaF/betaR | agggtcattacaccgagggt/gtaccaccaccaagagagtgg | 121 |
| MgvRTF/MgvRTR | aggttcaacgattccgacag/gaccattaccctgaggcaga | 100 |
| StuARTF/StuARTR | gcccctactggatacgatca/ttgccttctagggacattgg | 100 |
| Hog1F/Hog1R | cctggcaaaaatacgacgtt/tgatggagaattggttgacg | 117 |
| PodrtF/PodrtR | tggatcaaggacattggtga/gttggtagcatcctgctggt | 117 |
| tri5fullF/tri5fullR | atggagaactttcccaccgagtatt/agtccatagtgctacggataaggttcaa | 469 |
| CatrtF/CatrtR | aattccacgttcgtttcgtc/ccatactaggctcgctttgc | 130 |
| Mst20rtF/Mst20rtR | cctgaaaaggaacgacgaga/gccagcatgatggaatttct | 117 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gazdağlı, A.; Sefer, Ö.; Yörük, E.; Varol, G.İ.; Teker, T.; Albayrak, G. Investigation of Camphor Effects on Fusarium graminearum and F. culmorum at Different Molecular Levels. Pathogens 2018, 7, 90. https://doi.org/10.3390/pathogens7040090
Gazdağlı A, Sefer Ö, Yörük E, Varol Gİ, Teker T, Albayrak G. Investigation of Camphor Effects on Fusarium graminearum and F. culmorum at Different Molecular Levels. Pathogens. 2018; 7(4):90. https://doi.org/10.3390/pathogens7040090
Chicago/Turabian StyleGazdağlı, Aylin, Özlem Sefer, Emre Yörük, Gülin İnci Varol, Tuğba Teker, and Gülruh Albayrak. 2018. "Investigation of Camphor Effects on Fusarium graminearum and F. culmorum at Different Molecular Levels" Pathogens 7, no. 4: 90. https://doi.org/10.3390/pathogens7040090
APA StyleGazdağlı, A., Sefer, Ö., Yörük, E., Varol, G. İ., Teker, T., & Albayrak, G. (2018). Investigation of Camphor Effects on Fusarium graminearum and F. culmorum at Different Molecular Levels. Pathogens, 7(4), 90. https://doi.org/10.3390/pathogens7040090