Comparison of the Pathogenicity of Two Different Branches of Senecavirus a Strain in China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Genomic Sequence Analysis and Phylogenetic Analysis
2.3. One-Step Growth Curve
2.4. Electron Microscopy
2.5. Experimental Infection of Pigs
2.6. Quantitative Real-Time PCR
2.7. Cross-Neutralization Test
2.8. Histopathological Examination and Immunohistochemistry (IHC)
2.9. Statistical Analysis
3. Results
3.1. Genomic Sequence Comparison between SVA HB-CH-2016 and SVA CH/AH-02/2017 Strains
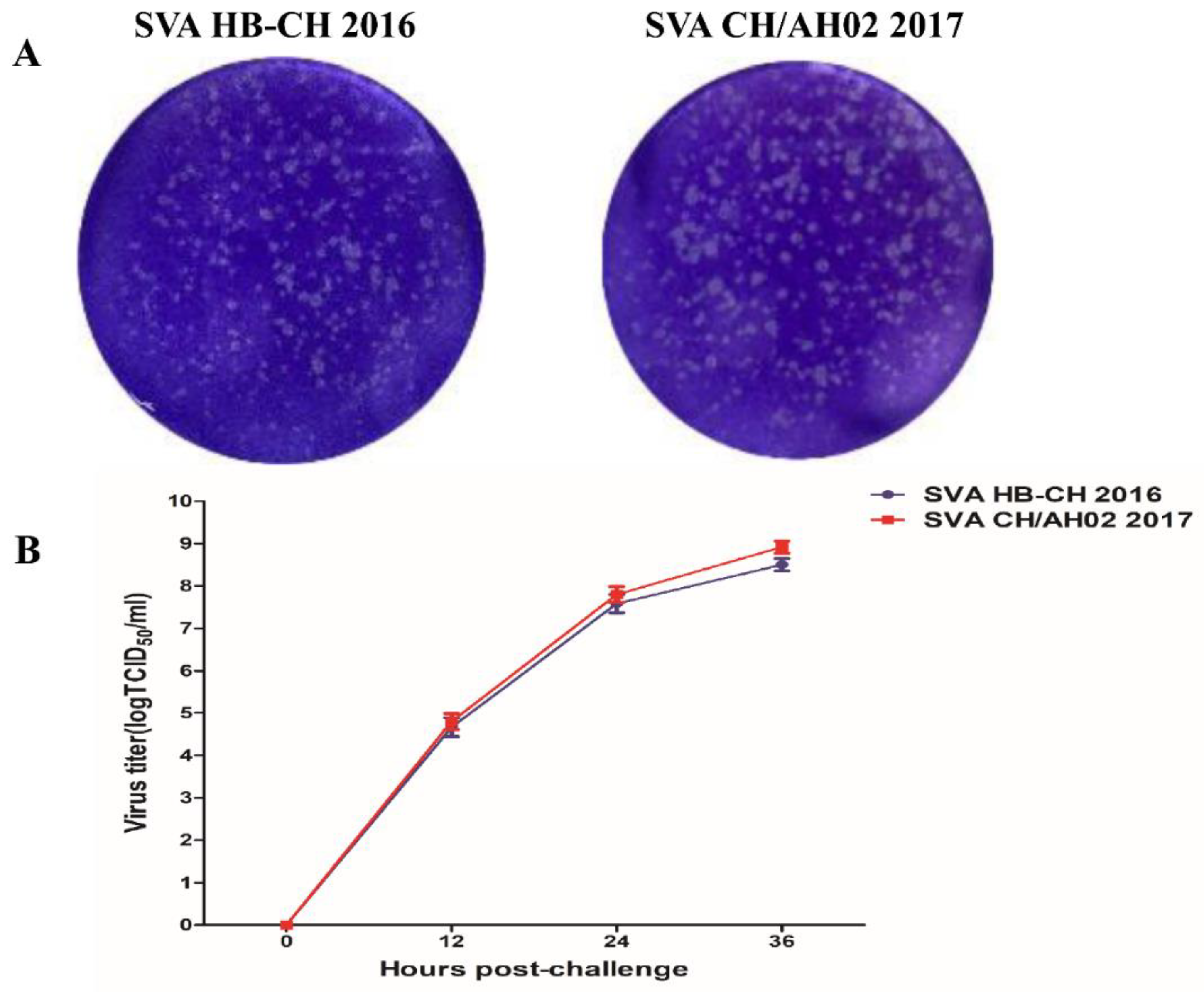
3.2. Replication Kinetics of the SVA HB-CH-2016 and SVA CH/AH-02/2017 Strains
3.3. Electron Microscopy
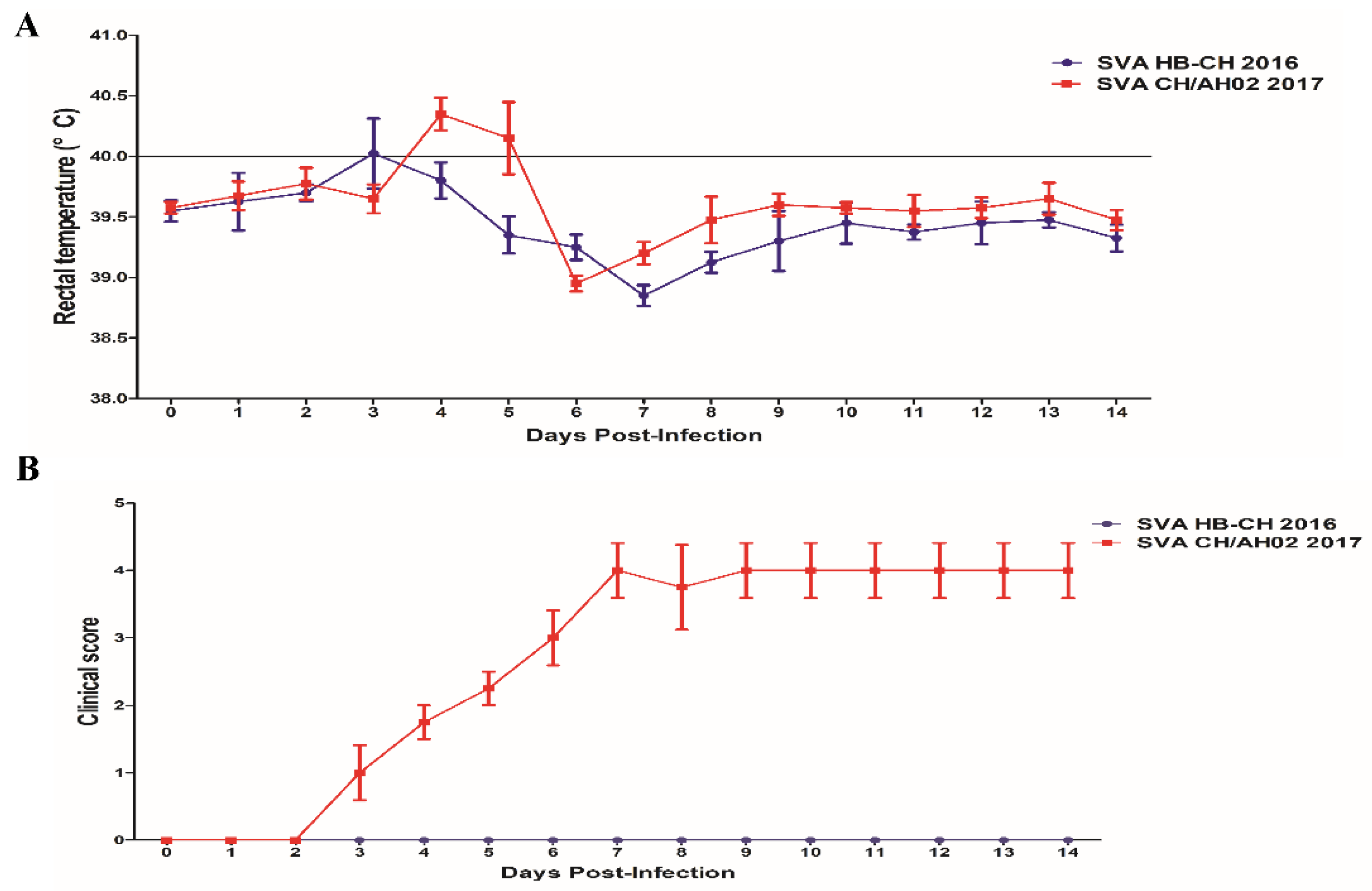
3.4. Pathogenicity of the SVA HB-CH-2016 and SVA CH/AH-02/2017 Strains
3.5. Histopathological Lesions
3.6. Detection of SVA Viremia Levels and Viral Shedding
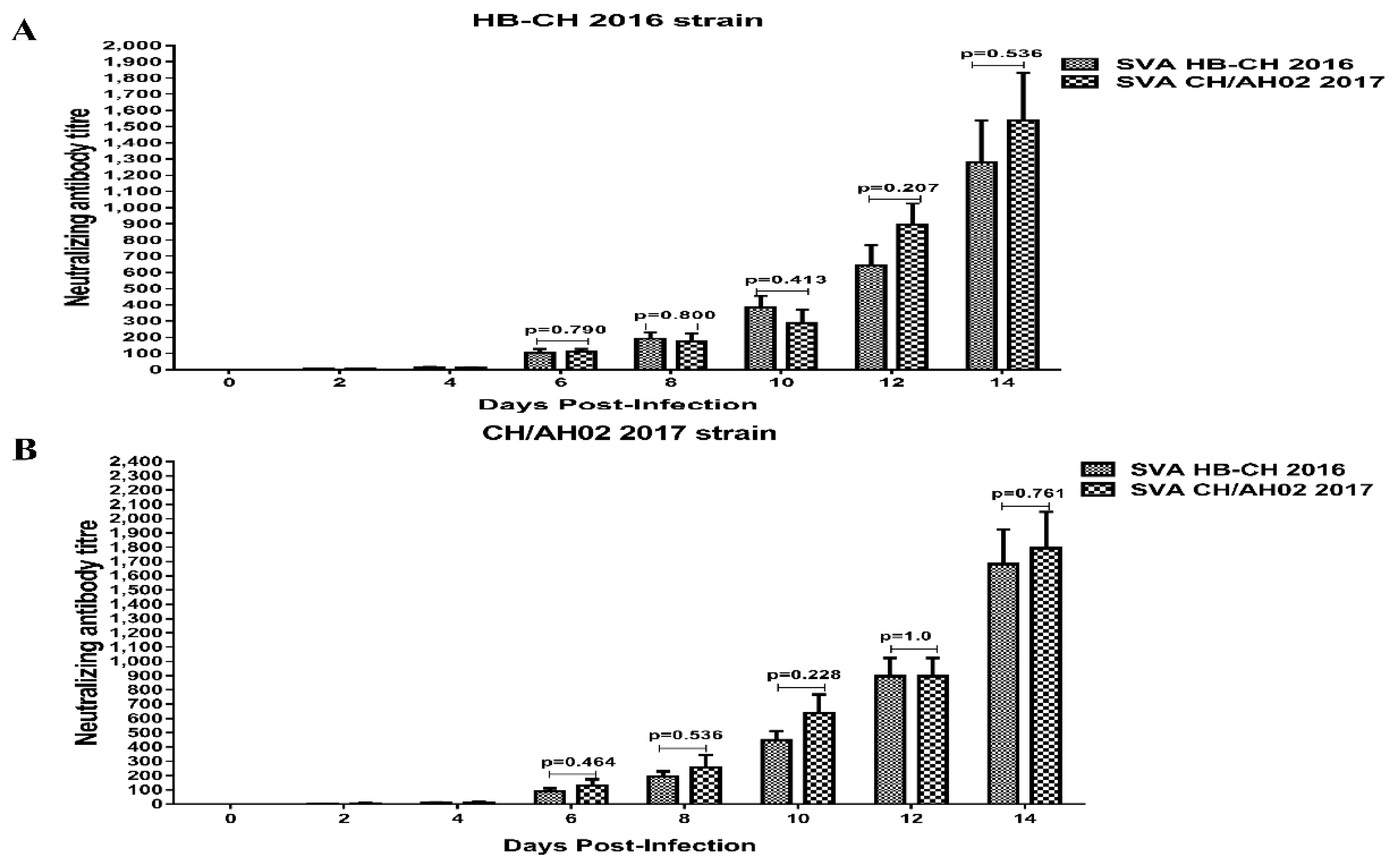
3.7. Cross-Neutralization between SVA HB-CH-2016 and SVA CH/AH-02/2017
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Hales, L.M.; Knowles, N.J.; Reddy, P.S.; Xu, L.; Hay, C.; Hallenbeck, P.L. Complete genome sequence analysis of Seneca Valley virus-001, a novel oncolytic picornavirus. J. Gen. Virol. 2008, 89, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, S.; Reddy, S.P.; Loo, J.; Idamakanti, N.; Hallenbeck, P.L.; Reddy, V.S. Structure of Seneca Valley Virus-001: An oncolytic picornavirus representing a new genus. Structure 2008, 16, 1555–1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkataraman, S.; Reddy, S.P.; Loo, J.; Idamakanti, N.; Hallenbeck, P.L.; Reddy, V.S. Crystallization and preliminary X-ray diffraction studies of Seneca Valley virus-001, a new member of the Picornaviridae family. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2008, 64, 293–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhu, Z.; Yang, F.; Cao, W.; Tian, H.; Zhang, K.; Zheng, H.; Liu, X. Review of seneca valley virus: A call for increased surveillance and research. Front. Microbiol. 2018, 9, 940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montiel, N.; Buckley, A.; Guo, B.; Kulshreshtha, V.; VanGeelen, A.; Hoang, H.; Rademacher, C.; Yoon, K.-J.; Lager, K. Vesicular disease in 9-week-old pigs experimentally infected with senecavirus A. Emerg. Infect. Dis. 2016, 22, 1246–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Yang, F.; Cao, W.; Liu, H.; Zhang, K.; Liu, X.; Xu, Z.; Zhu, Z.; Zheng, H. The Distribution of Different Clades of Seneca Valley Viruses in Guangdong Province, China. Virol. Sin. 2018, 33, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Buckley, A.; Kulshreshtha, V.; van Geelen, A.; Montiel, N.; Guo, B.; Yoon, K.J.; Lager, K. Experimental Seneca Valley virus infection in market-weight gilts. Vet. Microbiol. 2019, 231, 7–10. [Google Scholar] [CrossRef]
- Gimenez-Lirola, L.G.; Rademacher, C.; Linhares, D.; Harmon, K.; Rotolo, M.; Sun, Y.; Baum, D.H.; Zimmerman, J.; Piñeyro, P. Serological and molecular detection of senecavirus a associated with an outbreak of swine idiopathic vesicular disease and neonatal mortality. J. Clin. Microbiol. 2016, 54, 2082–2089. [Google Scholar] [CrossRef] [Green Version]
- Guo, B.; Piñeyro, P.E.; Rademacher, C.J.; Zheng, Y.; Li, G.; Yuan, J.; Hoang, H.; Gauger, P.C.; Madson, D.M.; Schwartz, K.J.; et al. Novel senecavirus a in swine with vesicular disease, United States, July 2015. Emerg. Infect. Dis. 2016, 22, 1325–1327. [Google Scholar] [CrossRef]
- Yang, F.; Zhu, Z.; Cao, W.; Liu, H.; Zhang, K.; Tian, H.; Liu, X.; Zheng, H. Immunogenicity and protective efficacy of an inactivated cell culture-derived Seneca Valley virus vaccine in pigs. Vaccine 2018, 36, 841–846. [Google Scholar] [CrossRef]
- Burke, M. Oncolytic Seneca Valley Virus: Past perspectives and future directions. Oncolytic Virother. 2016, 5, 81–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirier, J.T.; Dobromilskaya, I.; Moriarty, W.F.; Peacock, C.D.; Hann, C.L.; Rudin, C.M. Selective tropism of seneca valley virus for variant subtype small cell lung cancer. J. Natl. Cancer Inst. 2013, 105, 1059–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, P.S.; Burroughs, K.D.; Hales, L.M.; Ganesh, S.; Jones, B.H.; Idamakanti, N.; Hay, C.; Li, S.S.; Skele, K.L.; Vasko, A.J.; et al. 2007: Seneca Valley virus, a systemically deliverable oncolytic picornavirus, and the treatment of neuroendocrine cancers. J. Natl. Cancer Inst. 2007, 99, 1623–1633. [Google Scholar] [CrossRef] [PubMed]
- Rudin, C.M.; Poirier, J.T.; Senzer, N.N.; Stephenson, J.; Loesch, D.; Burroughs, K.D.; Reddy, P.S.; Hann, C.L.; Hallenbeck, P.L. 2011: Phase I clinical study of Seneca Valley Virus (SVV-001), a replication-competent picornavirus, in advanced solid tumors with neuroendocrine features. Clin. Cancer Res. 2011, 17, 888–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, M.H.V.; Maggioli, M.F.; Joshi, L.R.; Clement, T.; Faccin, T.C.; Rauh, R.; Bauermann, F.V.; Diel, D.G. Pathogenicity and cross-reactive immune responses of a historical and a contemporary Senecavirus A strains in pigs. Virology 2018, 522, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Zhao, X.; Chen, Y.; He, X.; Zhang, G.; Ma, J. Complete Genome Sequence of Seneca Valley Virus CH-01-2015 Identified in China. Genome Announc. 2016, 4, e01509-15. [Google Scholar] [CrossRef] [Green Version]
- Qian, S.; Fan, W.; Qian, P.; Chen, H.; Li, X. Isolation and full-genome sequencing of Seneca Valley virus in piglets from China, 2016. Virol. J. 2016, 13, 4–7. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Yang, F.; Chen, P.; Liu, H.; Cao, W.; Zhang, K.; Liu, X.; Zheng, H. Emergence of novel Seneca Valley virus strains in China, 2017. Transbound. Emerg. Dis. 2017, 64, 1024–1029. [Google Scholar] [CrossRef]
- Zhao, X.; Wu, Q.; Bai, Y.; Chen, G.; Zhou, L.; Wu, Z.; Li, Y.; Zhou, W.; Yang, H.; Ma, J. Phylogenetic and genome analysis of seven senecavirus A isolates in China. Transbound. Emerg. Dis. 2017, 64, 2075–2082. [Google Scholar] [CrossRef]
- Sun, Y.; Cheng, J.; Wu, R.-T.; Wu, Z.-X.; Chen, J.-W.; Luo, Y.; Xie, Q.-M.; Ma, J.-Y. Phylogenetic and Genome Analysis of 17 Novel Senecavirus A Isolates in Guangdong Province, 2017. Front. Vet. Sci. 2018, 5, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Yuan, F.; Li, Y.; Shang, P.; Schroeder, R.; Lechtenberg, K.; Henningson, J.; Hause, B.; Bai, J.; Rowland, R.R.R.; et al. Construction and characterization of a full-length cDNA infectious clone of emerging porcine Senecavirus A. Virology 2016, 497, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Joshi, L.R.; Fernandes, M.H.V.; Clement, T.; Lawson, S.; Pillatzki, A.; Resende, T.P.; Vannucci, F.A.; Kutish, G.F.; Nelson, E.A.; Diel, D.G. Pathogenesis of Senecavirus A infection in finishing pigs. J. Gen. Virol. 2016, 3267–3279. [Google Scholar] [CrossRef] [PubMed]
- Dall Agnol, A.M.; Miyabe, F.M.; Leme, R.A.; Oliveira, T.E.S.; Headley, S.A.; Alfieri, A.A.; Alfieri, A.F. Quantitative analysis of senecavirus A in tissue samples from naturally infected newborn piglets. Arch. Virol. 2018, 163, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Maggioli, M.F.; Lawson, S.; de Lima, M.; Joshi, L.R.; Faccin, T.C.; Bauermann, F.V.; Diel, D.G. Adaptive Immune Responses following Senecavirus A Infection in Pigs. J. Virol. 2018, 92, e01717-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Positions (AA) | SVA HB-CH2016 | SVACH/AH-02 2017 |
|---|---|---|---|
| protein L | 56 | R | K |
| protein VP4 | 83 | Q | H |
| protein VP2 | 426 | Y | F |
| 427 | K | T | |
| protein VP3 | 494 | P | S |
| 511 | T | A | |
| 575 | T | A | |
| protein VP1 | 738 | T | A |
| 767 | S | N | |
| 845 | A | 7 | |
| 904 | A | V | |
| 941 | V | I | |
| protein 2C | 1079 | K | T |
| 1148 | S | G | |
| protein 3A | 1401 | N | T |
| 1428 | Q | P | |
| 1429 | E | D | |
| 1462 | R | N | |
| 1469 | A | T | |
| protein 3C | 1589 | V | L |
| 1647 | E | D | |
| 1649 | T | S | |
| 1685 | L | M | |
| protein 3D | 1850 | V | A |
| 1856 | A | P | |
| 2023 | I | N | |
| 2035 | L | P | |
| 2079 | K | R | |
| 2158 | R | H |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Chen, P.; Hao, G.; Liu, W.; Chen, H.; Qian, P.; Li, X. Comparison of the Pathogenicity of Two Different Branches of Senecavirus a Strain in China. Pathogens 2020, 9, 39. https://doi.org/10.3390/pathogens9010039
Zhang H, Chen P, Hao G, Liu W, Chen H, Qian P, Li X. Comparison of the Pathogenicity of Two Different Branches of Senecavirus a Strain in China. Pathogens. 2020; 9(1):39. https://doi.org/10.3390/pathogens9010039
Chicago/Turabian StyleZhang, Huawei, Pin Chen, Genxi Hao, Wenqiang Liu, Huanchun Chen, Ping Qian, and Xiangmin Li. 2020. "Comparison of the Pathogenicity of Two Different Branches of Senecavirus a Strain in China" Pathogens 9, no. 1: 39. https://doi.org/10.3390/pathogens9010039
APA StyleZhang, H., Chen, P., Hao, G., Liu, W., Chen, H., Qian, P., & Li, X. (2020). Comparison of the Pathogenicity of Two Different Branches of Senecavirus a Strain in China. Pathogens, 9(1), 39. https://doi.org/10.3390/pathogens9010039