Correlation of NHR-48 Transcriptional Modulator Expression with Selected CYP Genes’ Expression during Thiabendazole Treatment of Anisakis simplex (s.l.)?—An In Vitro Study

 ,
,  , and
, and 
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
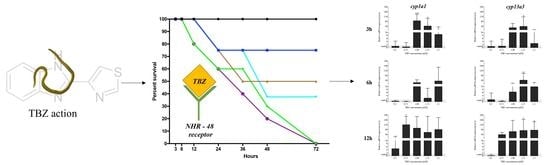
2.1. Survival of Larvae after TBZ Exposure
2.2. Bioinformatical Analyses
2.3. qReal-Time PCR
3. Discussion
4. Materials and Methods
4.1. Anisakis simplex (s.l.) Larvae
4.2. In Vitro Culture
4.3. Total RNA Isolation and cDNA Synthesis
4.4. Bioinformatical Analyses
4.5. qReal-Time PCR
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mattiucci, S.; Cipriani, P.; Levsen, A.; Paoletti, M.; Nascetti, G. Molecular Epidemiology of Anisakis and Anisakiasis: An Ecological and Evolutionary Road Map. Adv. Parasitol. 2018, 99, 93–263. [Google Scholar] [CrossRef]
- Valls, A.; Pascual, C.Y.; Martín Esteban, M. Anisakis allergy: An update. Rev. Fr. d’Allergol. d’Immunol. Clin. 2005, 45, 108–113. [Google Scholar] [CrossRef]
- Asaishi, K.; Nishino, C.; Ebata, T.; Totsuka, M.; Hayasaka, H.; Suzuki, T. Studies on the etiologic mechanism of anisakiasis. --1. Immunological reactions of digestive tract induced by Anisakis larva. Gastroenterol. Jpn. 1980, 15, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Sakanari, J.A.; McKerrow, J.H. Anisakiasis. Clin. Microbiol. Rev. 1989, 2, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Sakanari, J.A.; McKerrow, J.H. Identification of the Secreted Neutral Proteases from Anisakis simplex. J. Parasitol. 1990, 76, 625. [Google Scholar] [CrossRef]
- Mattiucci, S.; Fazii, P.; De Rosa, A.; Paoletti, M.; Megna, A.S.; Glielmo, A.; De Angelis, M.; Costa, A.; Meucci, C.; Calvaruso, V.; et al. Anisakiasis and Gastroallergic Reactions Associated with Anisakis pegreffii Infection, Italy. Emerg. Infect. Dis. 2013, 19, 496–499. [Google Scholar] [CrossRef]
- Audicana, M.T.; Kennedy, M.W. Anisakis simplex: From Obscure Infectious Worm to Inducer of Immune Hypersensitivity. Clin. Microbiol. Rev. 2008, 21, 360–379. [Google Scholar] [CrossRef]
- Audicana, M.T.; Ansotegui, I.J.; de Corres, L.F.; Kennedy, M.W. Anisakis simplex: Dangerous—Dead and alive? Trends Parasitol. 2002, 18, 20–25. [Google Scholar] [CrossRef]
- Moneo, I.; Caballero, M.L.; González-Muñoz, M.; Rodríguez-Mahillo, A.I.; Rodríguez-Perez, R.; Silva, A. Isolation of a heat-resistant allergen from the fish parasite Anisakis simplex. Parasitol. Res. 2005, 96, 285–289. [Google Scholar] [CrossRef]
- Bucci, C.; Gallotta, S.; Morra, I.; Fortunato, A.; Ciacci, C.; Iovino, P. Anisakis, just think about it in an emergency! Int. J. Infect. Dis. 2013, 17, e1071–e1072. [Google Scholar] [CrossRef]
- Dziekońska-Rynko, J.; Rokicki, J.; Jablonowski, Z. Effects of ivermectin and albendazole against Anisakis simplex in vitro and in guinea pigs. J. Parasitol. 2002, 88, 395–398. [Google Scholar] [CrossRef]
- Moore, D.A.; Girdwood, R.; Chiodini, P.L. Treatment of anisakiasis with albendazole. Lancet 2002, 360, 54. [Google Scholar] [CrossRef]
- Pacios, E.; Arias-Diaz, J.; Zuloaga, J.; Gonzalez-Armengol, J.; Villarroel, P.; Balibrea, J.L. Albendazole for the Treatment of Anisakiasis Ileus. Clin. Infect. Dis. 2005, 41, 1825–1826. [Google Scholar] [CrossRef] [PubMed]
- Ariasdiaz, J.; Zuloaga, J.; Vara, E.; Balibrea, J.; Balibrea, J. Efficacy of albendazole against Anisakis simplex larvae in vitro. Dig. Liver Dis. 2006, 38, 24–26. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.C.; Navarro, M.C.; Martín-Sánchez, J.; Valero, A. Peppermint (Mentha piperita) and albendazole against anisakiasis in an animal model. Trop. Med. Int. Health 2014, 19, 1430–1436. [Google Scholar] [CrossRef]
- Hierro, I.; Valero, A.; Pérez, P.; González, P.; Cabo, M.M.; Montilla, M.P.; Navarro, M.C. Action of different monoterpenic compounds against Anisakis simplex s.l. L3 larvae. Phytomedicine 2004, 11, 77–82. [Google Scholar] [CrossRef]
- Lin, R.-J.; Chen, C.-Y.; Lee, J.-D.; Lu, C.-M.; Chung, L.-Y.; Yen, C.-M. Larvicidal Constituents of Zingiber officinale (Ginger) against Anisakis simplex. Planta Med. 2010, 76, 1852–1858. [Google Scholar] [CrossRef]
- Del Carmen Romero, M.; Valero, A.; Martín-Sánchez, J.; Navarro-Moll, M.C. Activity of Matricaria chamomilla essential oil against anisakiasis. Phytomedicine 2012, 19, 520–523. [Google Scholar] [CrossRef]
- Jones, L.M.; Flemming, A.J.; Urwin, P.E. NHR-176 regulates cyp-35d1 to control hydroxylation-dependent metabolism of thiabendazole in Caenorhabditis elegans. Biochem. J. 2015, 466, 37–44. [Google Scholar] [CrossRef]
- Borgers, M.; Nollin, S. De Ultrastructural Changes in Ascaris suum Intestine after Mebendazole Treatment In vivo. J. Parasitol. 1975, 61, 110. [Google Scholar] [CrossRef]
- Sakai, C.; Tomitsuka, E.; Esumi, H.; Harada, S.; Kita, K. Mitochondrial fumarate reductase as a target of chemotherapy: From parasites to cancer cells. Biochim. Biophys. Acta Gen. Subj. 2012, 1820, 643–651. [Google Scholar] [CrossRef]
- Shompole, S.; Yao, C.; Cheng, X.; Knox, D.; Johnson, S.; Jasmer, D.P. Distinct characteristics of two intestinal protein compartments discriminated by using fenbendazole and a benzimidazole resistant isolate of Haemonchus contortus. Exp. Parasitol. 2002, 101, 200–209. [Google Scholar] [CrossRef]
- Kotze, A.C.; Cowling, K.; Bagnall, N.H.; Hines, B.M.; Ruffell, A.P.; Hunt, P.W.; Coleman, G.T. Relative level of thiabendazole resistance associated with the E198A and F200Y SNPs in larvae of a multi-drug resistant isolate of Haemonchus contortus. Int. J. Parasitol. Drugs Drug Resist. 2012, 2, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Stuchlíková, L.R.; Matoušková, P.; Vokřál, I.; Lamka, J.; Szotáková, B.; Sečkařová, A.; Dimunová, D.; Nguyen, L.T.; Várady, M.; Skálová, L. Metabolism of albendazole, ricobendazole and flubendazole in Haemonchus contortus adults: Sex differences, resistance-related differences and the identification of new metabolites. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 50–58. [Google Scholar] [CrossRef]
- Laing, S.T.; Ivens, A.; Laing, R.; Ravikumar, S.; Butler, V.; Woods, D.J.; Gilleard, J.S. Characterization of the xenobiotic response of Caenorhabditis elegans to the anthelmintic drug albendazole and the identification of novel drug glucoside metabolites. Biochem. J. 2010, 432, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Prichard, R.K.; Hennessy, D.R.; Steel, J.W.; Lacey, E. Metabolite concentrations in plasma following treatment of cattle with five anthelmintics. Res. Vet. Sci. 1985, 39, 173–178. [Google Scholar] [CrossRef]
- Chukwudebe, A.C.; Wislocki, P.G.; Sanson, D.R.; Halls, T.D.J.; VandenHeuvel, W.J.A. Metabolism of Thiabendazole in Laying Hen and Lactating Goats. J. Agric. Food Chem. 1994, 42, 2964–2969. [Google Scholar] [CrossRef]
- Gonzalez, F.; Tukey, R. Drug metabolism. In Goodman & Gilman’s the Pharmacological Basis of Therapeutics; Parker, K., Ed.; McGraw-Hill Medical Publishing Division: New York, NY, USA, 2006; pp. 71–91. [Google Scholar]
- Zamanian, M.; Cook, D.E.; Zdraljevic, S.; Brady, S.C.; Lee, D.; Lee, J.; Andersen, E.C. Discovery of genomic intervals that underlie nematode responses to benzimidazoles. PLoS Negl. Trop. Dis. 2018, 12, e0006368. [Google Scholar] [CrossRef]
- Stasiuk, S.J.; MacNevin, G.; Workentine, M.L.; Gray, D.; Redman, E.; Bartley, D.; Morrison, A.; Sharma, N.; Colwell, D.; Ro, D.K.; et al. Similarities and differences in the biotransformation and transcriptomic responses of Caenorhabditis elegans and Haemonchus contortus to five different benzimidazole drugs. Int. J. Parasitol. Drugs Drug Resist. 2019, 11, 13–29. [Google Scholar] [CrossRef]
- Jones, L.M.; Rayson, S.J.; Flemming, A.J.; Urwin, P.E. Adaptive and Specialised Transcriptional Responses to Xenobiotic Stress in Caenorhabditis elegans Are Regulated by Nuclear Hormone Receptors. PLoS ONE 2013, 8, e69956. [Google Scholar] [CrossRef]
- Chai, X.; Zeng, S.; Xie, W. Nuclear receptors PXR and CAR: Implications for drug metabolism regulation, pharmacogenomics and beyond. Expert Opin. Drug Metab. Toxicol. 2013, 9, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Ménez, C.; Alberich, M.; Courtot, E.; Guegnard, F.; Blanchard, A.; Aguilaniu, H.; Lespine, A. The transcription factor NHR-8: A new target to increase ivermectin efficacy in nematodes. PLoS Pathog. 2019, 15, e1007598. [Google Scholar] [CrossRef] [PubMed]
- Laing, R.; Bartley, D.J.; Morrison, A.A.; Rezansoff, A.; Martinelli, A.; Laing, S.T.; Gilleard, J.S. The cytochrome P450 family in the parasitic nematode Haemonchus contortus. Int. J. Parasitol. 2015, 45, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Mejía-Jaramillo, A.M.; Fernández, G.J.; Palacio, L.; Triana-Chávez, O. Gene expression study using real-time PCR identifies an NTR gene as a major marker of resistance to benznidazole in Trypanosoma cruzi. Parasit. Vectors 2011, 4, 169. [Google Scholar] [CrossRef]
- Kotze, A.C.; Hunt, P.W.; Skuce, P.; von Samson-Himmelstjerna, G.; Martin, R.J.; Sager, H.; Krücken, J.; Hodgkinson, J.; Lespine, A.; Jex, A.R.; et al. Recent advances in candidate-gene and whole-genome approaches to the discovery of anthelmintic resistance markers and the description of drug/receptor interactions. Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 164–184. [Google Scholar] [CrossRef]
- Edwards, D.P. The role of coactivators and corepressors in the biology and mechanism of action of steroid hormone receptors. J. Mammary Gland Biol. Neoplasia 2000, 5, 307–324. [Google Scholar] [CrossRef]
- Magner, D.B.; Antebi, A. Caenorhabditis elegans nuclear receptors: Insights into life traits. Trends Endocrinol. Metab. 2008, 19, 153–160. [Google Scholar] [CrossRef]
- Ruaud, A.-F.; Bessereau, J.-L. Activation of nicotinic receptors uncouples a developmental timer from the molting timer in C. elegans. Development 2006, 133, 2211–2222. [Google Scholar] [CrossRef]
- Holden-Dye, L.; Walker, R.J. How relevant is Caenorhabditis elegans as a model for the analysis of parasitic nematode biology? In Parasitic Helminths: Targets, Screens, Drugs and Vaccines; Caffrey, C.R., Ed.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2012; pp. 23–41. [Google Scholar]
- AlGusbi, S.; Krücken, J.; Ramünke, S.; von Samson-Himmelstjerna, G.; Demeler, J. Analysis of putative inhibitors of anthelmintic resistance mechanisms in cattle gastrointestinal nematodes. Int. J. Parasitol. 2014, 44, 647–658. [Google Scholar] [CrossRef]
- Menzel, R.; Rödel, M.; Kulas, J.; Steinberg, C.E.W. CYP35: Xenobiotically induced gene expression in the nematode Caenorhabditis elegans. Arch. Biochem. Biophys. 2005, 438, 93–102. [Google Scholar] [CrossRef]
- Chakrapani, B.P.S.; Kumar, S.; Subramaniam, J.R. Development and evaluation of an in vivo assay in Caenorhabditis elegans for screening of compounds for their effect on cytochrome P450 expression. J. Biosci. 2008, 33, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Albérich, M.; Ménez, C.; Sutra, J.-F.; Lespine, A. Ivermectin exposure leads to up-regulation of detoxification genes in vitro and in vivo in mice. Eur. J. Pharmacol. 2014, 740, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Lindblom, T.H.; Pierce, G.J.; Sluder, A.E. A C. elegans orphan nuclear receptor contributes to xenobiotic resistance. Curr. Biol. 2001, 11, 864–868. [Google Scholar] [CrossRef]
- Iglesisas, L.; Valero, A.; Benitez, R.; Adroher, F.J. In vitro cultivation of Anisakis simplex: Pepsin increases survival and moulting from fourth larval to adult stage. Parasitology 2001, 123, 285–291. [Google Scholar] [CrossRef]
- Łopieńska-Biernat, E.; Paukszto, Ł.; Jastrzębski, J.P.; Myszczyński, K.; Polak, I.; Stryiński, R. Genome-wide analysis of Anisakis simplex sensu lato: The role of carbohydrate metabolism genes in the parasite’s development. Int. J. Parasitol. 2019, 49, 933–943. [Google Scholar] [CrossRef]
- Stanke, M.; Morgenstern, B. AUGUSTUS: A web server for gene prediction in eukaryotes that allows user-defined constraints. Nucleic Acids Res. 2005, 33, W465–W467. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Katoh, K. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3--new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Łopieńska-Biernat, E.; Stryiński, R.; Paukszto, Ł.; Jastrzębski, J.P.; Makowczenko, K. The Selection of Reliable Reference Genes for RT-qPCR Analysis of Anisakis simplex Sensu Stricto Gene Expression from Different Developmental Stages. Acta Parasitol. 2020, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Łopieńska-Biernat, E.; Stryiński, R.; Paukszto, Ł.; Jastrzębski, J.P. Correlation of NHR-48 Transcriptional Modulator Expression with Selected CYP Genes’ Expression during Thiabendazole Treatment of Anisakis simplex (s.l.)?—An In Vitro Study. Pathogens 2020, 9, 1030. https://doi.org/10.3390/pathogens9121030
Łopieńska-Biernat E, Stryiński R, Paukszto Ł, Jastrzębski JP. Correlation of NHR-48 Transcriptional Modulator Expression with Selected CYP Genes’ Expression during Thiabendazole Treatment of Anisakis simplex (s.l.)?—An In Vitro Study. Pathogens. 2020; 9(12):1030. https://doi.org/10.3390/pathogens9121030
Chicago/Turabian StyleŁopieńska-Biernat, Elżbieta, Robert Stryiński, Łukasz Paukszto, and Jan P. Jastrzębski. 2020. "Correlation of NHR-48 Transcriptional Modulator Expression with Selected CYP Genes’ Expression during Thiabendazole Treatment of Anisakis simplex (s.l.)?—An In Vitro Study" Pathogens 9, no. 12: 1030. https://doi.org/10.3390/pathogens9121030
APA StyleŁopieńska-Biernat, E., Stryiński, R., Paukszto, Ł., & Jastrzębski, J. P. (2020). Correlation of NHR-48 Transcriptional Modulator Expression with Selected CYP Genes’ Expression during Thiabendazole Treatment of Anisakis simplex (s.l.)?—An In Vitro Study. Pathogens, 9(12), 1030. https://doi.org/10.3390/pathogens9121030