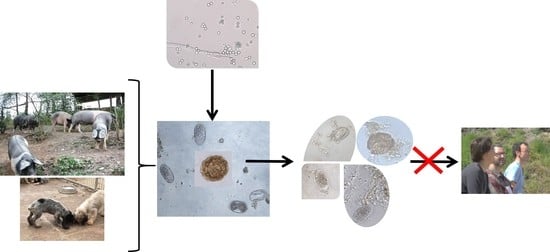
The Control of Zoonotic Soil-Transmitted Helminthoses Using Saprophytic Fungi


 , and
, and Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Share and Cite
Viña, C.; Silva, M.I.; Palomero, A.M.; Voinot, M.; Vilá, M.; Hernández, J.Á.; Paz-Silva, A.; Sánchez-Andrade, R.; Cazapal-Monteiro, C.F.; Arias, M.S. The Control of Zoonotic Soil-Transmitted Helminthoses Using Saprophytic Fungi. Pathogens 2020, 9, 1071. https://doi.org/10.3390/pathogens9121071
Viña C, Silva MI, Palomero AM, Voinot M, Vilá M, Hernández JÁ, Paz-Silva A, Sánchez-Andrade R, Cazapal-Monteiro CF, Arias MS. The Control of Zoonotic Soil-Transmitted Helminthoses Using Saprophytic Fungi. Pathogens. 2020; 9(12):1071. https://doi.org/10.3390/pathogens9121071
Chicago/Turabian StyleViña, Cándido, María Isabel Silva, Antonio Miguel Palomero, Mathilde Voinot, María Vilá, José Ángel Hernández, Adolfo Paz-Silva, Rita Sánchez-Andrade, Cristiana Filipa Cazapal-Monteiro, and María Sol Arias. 2020. "The Control of Zoonotic Soil-Transmitted Helminthoses Using Saprophytic Fungi" Pathogens 9, no. 12: 1071. https://doi.org/10.3390/pathogens9121071
APA StyleViña, C., Silva, M. I., Palomero, A. M., Voinot, M., Vilá, M., Hernández, J. Á., Paz-Silva, A., Sánchez-Andrade, R., Cazapal-Monteiro, C. F., & Arias, M. S. (2020). The Control of Zoonotic Soil-Transmitted Helminthoses Using Saprophytic Fungi. Pathogens, 9(12), 1071. https://doi.org/10.3390/pathogens9121071