Pantoea ananatis, A New Bacterial Pathogen Affecting Wheat Plants (Triticum L.) in Poland



Abstract
1. Introduction
2. Results
2.1. Field Observations
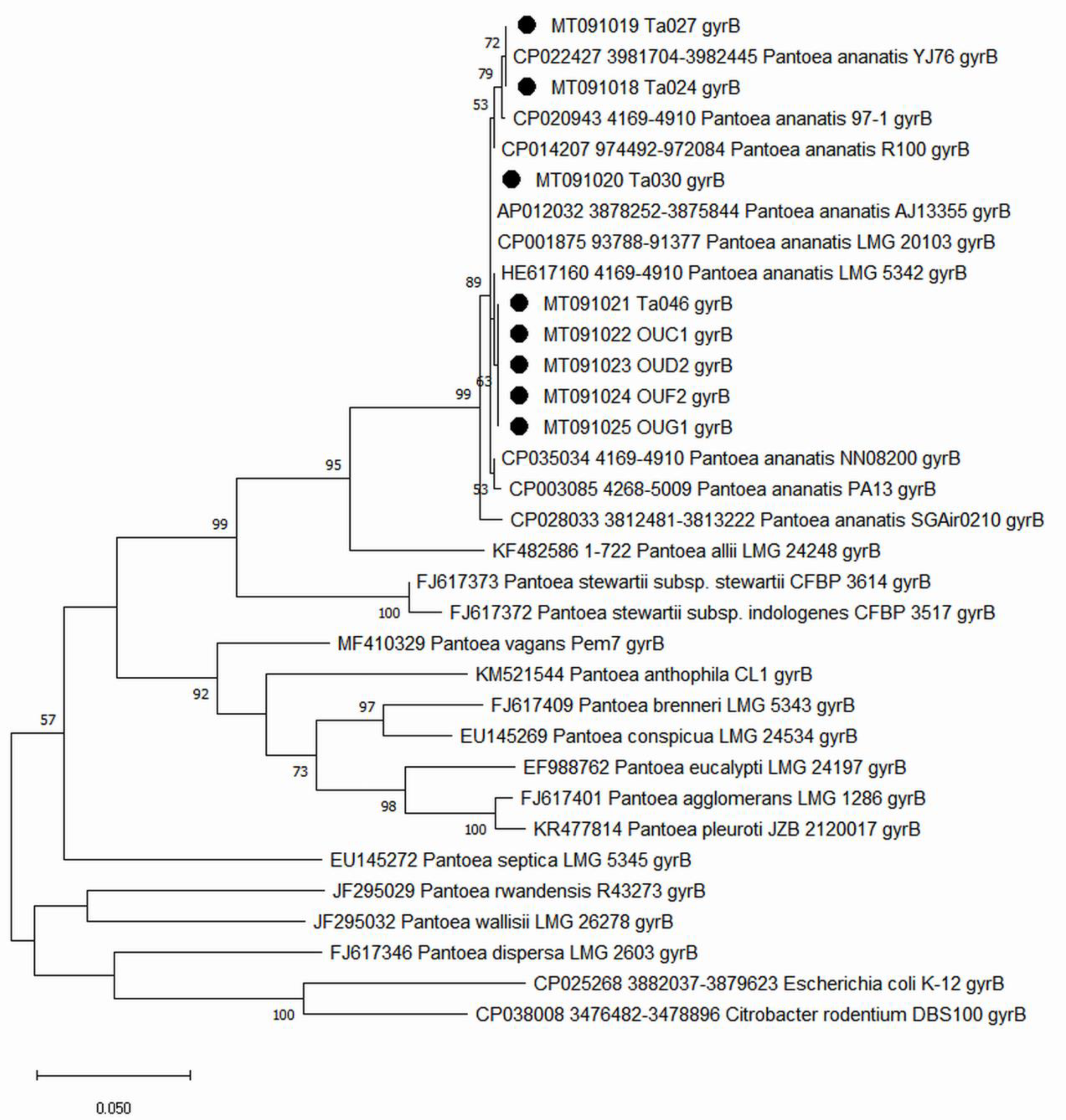
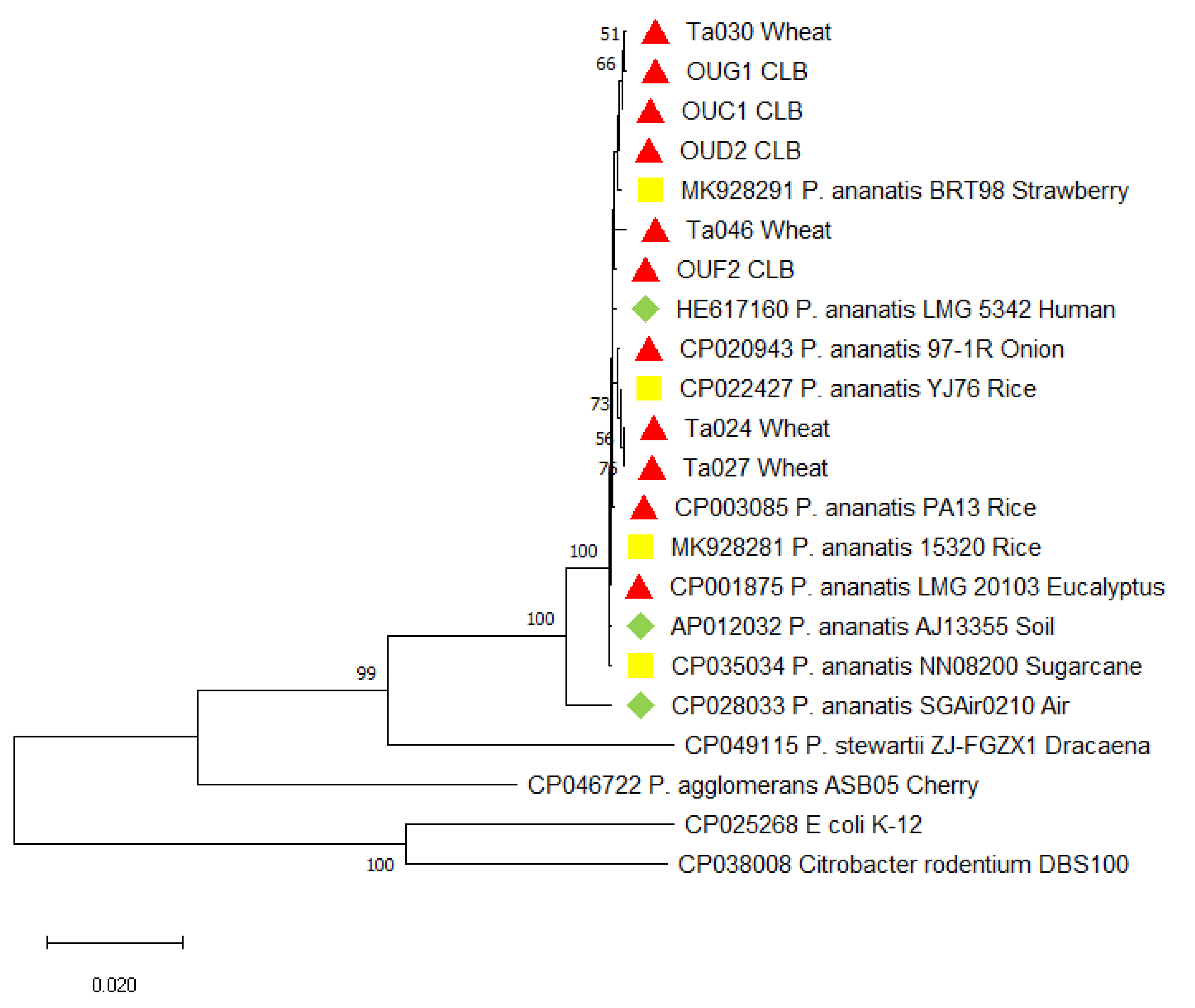
2.2. Identificaiton and Characteristics of Bacterial Strains
2.3. Pathogenicity Assessment of Strains
3. Discussion
4. Materials and Methods
4.1. Plant and Insect Material
4.2. Bacteria Isolation from Plants
4.3. Bacteria Isolation from Insects
4.4. Bacterial Identification, DNA Extraction, and Sequence Analysis
4.5. Phylogenetic Analysis of the Tested Pantoea Ananatis Strains
4.6. Assessing Pathogenicity of Bacterial Strains
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Peña-Bautista:, R.J.; Hernandez-Espinosa, N.; Jones, J.M.; Guzmán, C.; Braun, H.J. CIMMYT Series on Carbohydrates, Wheat, Grains, and Health: Wheat-Based Foods: Their Global and Regional Importance in the Food Supply, Nutrition, and Health. Cereal Foods World 2017, 62, 231–249. [Google Scholar]
- Niszczota, S.; Miziołek, D. Result estimate of the main agricultural and horticultural crops in 2019. Cent. Stat. Off. 2019, 12, 1–8. [Google Scholar]
- Kher, S.V.; Dosdall, L.M.; Cárcamo, H.A. The cereal leaf beetle: Biology, distribution and prospects for control. Prairie Soils Crop. 2011, 4, 32–41. [Google Scholar]
- Woods, S.; Olfert, O.; Philip, H.; Dosdall, L.; Weiss, R.M. Potential distribution and relative abundance of an invasive cereal crop pest, Oulema melanopus (Coleoptera: Chrysomelidae), in Canada. Can. Entomol. 2009, 136, 277–287. [Google Scholar]
- Nault, L.R. Transmission of Maize Chlorotic Mottle Virus by Chrysomelid Beetles. Phytopathology 1978, 68, 1071–1074. [Google Scholar] [CrossRef]
- Schaad, N.W.; Forster, R.L. A semiselective agar medium for isolating xanthomonas campestris.pdf. Phytopathology 1985, 75, 260–263. [Google Scholar]
- Toben, H.; Mavridis, A.; Rudolph, K.W.E. Basal glume rot (Pseudomonas syringae pv. atrofaciens) on wheat and barley in FRG and resistance screening of wheat. EPPO Bull. 1989, 19, 119–125. [Google Scholar]
- Hosford, R.M.J. White Blotch Incited in Wheat by Bacillus megaterium pv. cerealis. Phytopathology 1982, 72, 1453–1459. [Google Scholar]
- Piening, L.J.; Macpherson, D.J. Stem melanosis, a disease of spring wheat caused by pseudomonas cichorii. Can. J. Plant Pathol. 1985, 7, 168–172. [Google Scholar]
- Roberts, P. Erwinia rhapontici (Millard) Burkholder Associated with Pink Grain of Wheat. J. Appl. Bacteriol. 1974, 37, 353–358. [Google Scholar]
- Valencia-Botín, A.J.; Cisneros-Opez, M.E. Pseudomonas syringae Pathovars on Wheat. Int. J. Agron. 2012, 5, 1–5. [Google Scholar]
- Bach, H.J.; Jessen, I.; Schloter, M.; Munch, J.C. A TaqMan-PCR protocol for quantification and differentiation of the phytopathogenic Clavibacter michiganensis subspecies. J. Microbiol. Methods 2003, 52, 85–91. [Google Scholar] [PubMed]
- Midha, S.K.; Swarup, G. Factors affecting development of ear-cockle and tundu diseases of wheat. Indian J. Nematol. 1972, 2, 97–104. [Google Scholar]
- Krawczyk, K.; Kamasa, J.; Zwolinska, A.; Pospieszny, H. First report of Pantoea ananatis associated with leaf spot disease of maize in Poland. J. Plant Pathol. 2010, 92, 807–811. [Google Scholar]
- Azad, H.R.; Holmes, G.J.; Cooksey, D.A. A New Leaf Blotch Disease of Sudangrass Caused by Pantoea ananas and Pantoea stewartii. Plant Dis. 2007, 84, 973–979. [Google Scholar]
- Kaga, H.; Mano, H.; Tanaka, F.; Watanabe, A.; Kaneko, S.; Morisaki, H. Rice Seeds as Sources of Endophytic Bacteria. Microbes Environ. 2009, 24, 154–162. [Google Scholar]
- Stice, S.P.; Stumpf, S.D.; Gitaitis, R.D.; Kvitko, B.H.; Dutta, B. Pantoea ananatis Genetic Diversity Analysis Reveals Limited Genomic Diversity as Well as Accessory Genes Correlated with Onion Pathogenicity. Front. Microbiol. 2018, 9, 1–18. [Google Scholar]
- Choi, O.; Lim, J.Y.; Seo, Y.S.; Hwang, I.; Kim, J. Complete genome sequence of the rice pathogen Pantoea ananatis strain PA13. J. Bacteriol. 2012, 194, 531. [Google Scholar]
- Yu, X.; Jiang, J.; Liang, C.; Zhang, X.; Wang, J.; Shen, D.; Feng, Y. Indole affects the formation of multicellular aggregate structures in Pantoea agglomerans YS19. J. Gen. Appl. Microbiol. 2016, 62, 31–37. [Google Scholar]
- Nadarasah, G.; Stavrinides, J. Quantitative evaluation of the host-colonizing capabilities of the enteric bacterium Pantoea using plant and insect hosts. Microbiol. (U.K.) 2014, 160, 602–615. [Google Scholar]
- Hara, Y.; Kadotani, N.; Izui, H.; Katashkina, J.I.; Kuvaeva, T.M.; Andreeva, I.G.; Golubeva, L.I.; Malko, D.B.; Makeev, V.J.; Mashko, S.V.; et al. The complete genome sequence of Pantoea ananatis AJ13355, an organism with great biotechnological potential. Appl. Microbiol. Biotechnol. 2012, 93, 331–341. [Google Scholar] [CrossRef] [PubMed]
- De Maayer, P.; Chan, W.Y.; Rezzonico, F.; Bühlmann, A.; Venter, S.N.; Blom, J.; Goesmann, A.; Frey, J.E.; Smits, T.H.M.; Duffy, B.; et al. Complete genome sequence of clinical isolate Pantoea ananatis LMG 5342. J. Bacteriol. 2012, 194, 1615–1616. [Google Scholar] [CrossRef] [PubMed]
- Luhung, I.; Junqueira, A.C.M.; Uchida, A.; Purbojati, R.W.; Houghton, J.N.I.; Chénard, C.; Wong, A.; Clare, M.E.; Kushwaha, K.K.; Panicker, D.; et al. Genome sequence of Pantoea ananatis SGAir0210, isolated from outdoor air in Singapore. Genome Announc. 2018, 6, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, K.; Foryś, J.; Nakonieczny, M.; Tarnawska, M.; Bereś, P.K. Transmission of Pantoea ananatis, the causal agent of leaf spot disease of maize ( Zea mays ), by western corn rootworm ( Diabrotica virgifera virgifera LeConte ). Crop Prot. 2020. [Google Scholar] [CrossRef]
- Gitaitis, R.D.; Walcott, R.R.; Wells, M.L.; Perez, J.C.D.; Sanders, F.H. Transmission of Pantoea ananatis, Causal Agent of Center Rot of Onion, by Tobacco Thrips, Frankliniella fusca. Plant Dis. 2003, 87, 675–678. [Google Scholar] [CrossRef]
- Gitaitis, R.D.; Walcott, R.R.; Culpepper, S.; Sanders, H.; Zolobowska, L.; Langston, D. Recovery of Pantoea ananatis, causal agent of center rot of onion, from weeds and crops in Georgia, USA. Crop Prot. 2002, 21, 983–989. [Google Scholar] [CrossRef]
- Wells, M.L.; Gitaitis, R.D.; Sanders, F.H. Association of Tobacco Thrips, Frankliniella fusca (Thysanoptera: Thripidae) with Two Species of Bacteria of the Genus Pantoea. Ann. Entomol. Soc. Am. 2002, 95, 719–723. [Google Scholar] [CrossRef]
- Dutta, B.; Barman, A.K.; Srinivasan, R.; Avci, U.; Ullman, D.E.; Langston, D.B.; Gitaitis, R.D. Transmission of Pantoea ananatis and P. agglomerans, Causal Agents of Center Rot of Onion ( Allium cepa ), by Onion Thrips ( Thrips tabaci ) Through Feces. Phytopathology 2014, 104, 812–819. [Google Scholar] [CrossRef]
- Gullino, M.L.; Gilardi, G.; Sanna, M.; Garibaldi, A. Epidemiology of Pseudomonas syringae pv. syringae on tomato. Phytoparasitica 2009, 37, 461–466. [Google Scholar] [CrossRef]
- Nadarasah, G.; Stavrinides, J. Insects as alternative hosts for phytopathogenic bacteria. FEMS Microbiol. Rev. 2011, 35, 555–575. [Google Scholar] [CrossRef]
- Coutinho, T.A.; Venter, S.N. Pantoea ananatis: An unconventional plant pathogen. Mol. Plant Pathol. 2009, 10, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Wantanabe, K.; Sato, M. Survival and Characteristics of Ice Nucleation-active Bacteria on Mulberry Trees (Morus spp.) and in Mulberry Pyralid (Glyphodes pyloalis). Jpn. J. Phytopathol. 1995, 61, 439–443. [Google Scholar] [CrossRef]
- Murrell, A.; Dobson, S.J.; Yang, X.; Lacey, E.; Barker, S.C. A survey of bacterial diversity in ticks, lice and fleas from Australia. Parasitol. Res. 2003, 89, 326–334. [Google Scholar] [CrossRef] [PubMed]
- de Maayer, P.; Aliyu, H.; Vikram, S.; Blom, J.; Duffy, B.; Cowan, D.A.; Smits, T.H.M.; Venter, S.N.; Coutinho, T.A. Phylogenomic, pan-genomic, pathogenomic and evolutionary genomic insights into the agronomically relevant enterobacteria Pantoea ananatis and Pantoea stewartii. Front. Microbiol. 2017, 8, 1–14. [Google Scholar] [CrossRef]
- Bezděk, J.; Baselga, A. Revision of western Palaearctic species of the Oulema melanopus group, with description of two new species from Europe (Coleoptera: Chrysomelidae: Criocerinae). Acta Entomol. Musei Natl. Pragae 2015, 55, 273–304. [Google Scholar]
- García-González, T.; Sáenz-Hidalgo, H.K.; Silva-Rojas, H.V.; Morales-Nieto, C.; Vancheva, T.; Koebnik, R.; Ávila-Quezada, G.D. Enterobacter cloacae, an emerging plant-pathogenic bacterium affecting chili pepper seedlings. Plant Pathol. J. 2018, 34, 1–10. [Google Scholar]
- Korbie, D.J.; Mattick, J.S. Touchdown PCR for increased specificity and sensitivity in PCR amplification. Nat. Protoc. 2008, 3, 13–15. [Google Scholar] [CrossRef]
- Brady, C.; Cleenwerck, I.; Venter, S.; Vancanneyt, M.; Swings, J.; Coutinho, T. Phylogeny and identification of Pantoea species associated with plants, humans and the natural environment based on multilocus sequence analysis (MLSA). Syst. Appl. Microbiol. 2008, 31, 447–460. [Google Scholar] [CrossRef]
- Gerischer, U. Acinetobacter Molecular Microbiology; Gerischer, U., Ed.; Caister Academic Press: Ulm, Germany, 2008; ISBN 97819044552021904455204. [Google Scholar]
- Lee, H.; Churey, J.J.; Worobo, R.W. Biosynthesis and transcriptional analysis of thurincin H, a tandem repeated bacteriocin genetic locus, produced by Bacillus thuringiensis SF361. FEMS Microbiol. Lett. 2009, 299, 205–213. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Strain Tested | Symptoms Group 1/Season | Biolog Gen III Identification Result | Wheat Pathogenic (Yes/No) | 16S rDNA Sequencing Result (GenBank Accession Number) | gyrB Sequencing Result (GenBank Accession Number) |
|---|---|---|---|---|---|---|
| Season 2018 | ||||||
| 1 | Ta015 | 1/2018 | Bacillus licheniformis | No | nt | nt |
| 2 | Ta016 | 1/2018 | Bacillus licheniformis | No | nt | nt |
| 3 | Ta017 | 1/2018 | Pantoea agglomerans | No | nt | nt |
| 4 | Ta018 | 1/2018 | Bacillus licheniformis | No | nt | nt |
| 5 | Ta019 | 1/2018 | Macrococcus brunensis | No | nt | nt |
| 6 | Ta020 | 1/2018 | Macrococcus brunensis | No | nt | nt |
| 7 | Ta021 | 1/2018 | Pantoea agglomerans | No | nt | nt |
| 8 | Ta022 | 1/2018 | Pantoea agglomerans | No | nt | nt |
| 9 | Ta023 | 2/2018 | Bacillus licheniformis | No | nt | nt |
| 10 | Ta024 | 3/2018 | Pantoea ananatis | Yes | MH973236 | MT091018 |
| 11 | Ta025 | 3/2018 | Bacillus licheniformis | No | nt | nt |
| 12 | Ta026 | 3/2018 | Pseudomonas fluorescens | No | nt | nt |
| 13 | Ta027 | 3/2018 | Pantoea ananatis | Yes | MH973237 | MT091019 |
| 14 | Ta028 | 2/2018 | Pantoea agglomerans | No | nt | nt |
| 15 | Ta029 | 3/2018 | Pseudomonas fluorescens | No | nt | nt |
| 16 | Ta030 | 3/2018 | Pantoea ananatis | Yes | MH973238 | MT091020 |
| 17 | Ta031 | 2/2018 | Bacillus licheniformis | No | nt | nt |
| 18 | Ta032 | 2/2018 | Macrococcus brunensis | No | nt | nt |
| 19 | Ta033 | 2/2018 | Bacillus pseudomycoides | No | nt | nt |
| 20 | Ta034 | 2/2018 | Pseudomonas flavescens | No | nt | nt |
| 21 | Ta035 | 2/2018 | Pseudomonas flavescens | No | nt | nt |
| 22 | Ta036 | 2/2018 | Pantoea agglomerans | No | nt | nt |
| Season 2019 | ||||||
| 23 | Ta037 | 2/2019 | Pantoea agglomerans | No | nt | nt |
| 24 | Ta038 | 2/2019 | Pantoea agglomerans | No | nt | nt |
| 25 | Ta039 | 2/2019 | Pseudomonas synthaxa | No | nt | nt |
| 26 | Ta040 | 2/2019 | Pseudomonas synthaxa | No | nt | nt |
| 27 | Ta041 | 1/2019 | Pantoea agglomerans | No | nt | nt |
| 28 | Ta042 | 1/2019 | Pantoea agglomerans | No | nt | nt |
| 29 | Ta043 | 1/2019 | Pantoea agglomerans | No | nt | nt |
| 30 | Ta044 | 1/2019 | Pantoea agglomerans | No | nt | nt |
| 31 | Ta045 | 3/2019 | Pseudomonas fluorescens | No | nt | nt |
| 32 | Ta046 | 3/2019 | Pantoea ananatis | Yes | MH973239 | MT091021 |
| No | Strain Tested | Sample/Season | Biolog Gen III identification Result | Wheat Pathogenic (Yes/No) | 16S rDNA Sequencing Result (GenBank Accession Number) | gyrB Sequencing Result (GenBank Accession Number) |
|---|---|---|---|---|---|---|
| Season 2018 | ||||||
| 1 | OUC1 | 1/2018 | Pantoea ananatis | Yes | MT234395 | MT091022 |
| 2 | OUC2 | 1/2018 | Pantoea agglomerans | No | nt | nt |
| 3 | OUC3 | 1/2018 | Serratia liquefaciens | No | nt | nt |
| 4 | OUD1 | 2/2018 | Pantoea agglomerans | No | nt | nt |
| 5 | OUD2 | 2/2018 | Pantoea ananatis | Yes | MT234396 | MT091023 |
| 6 | OUD3 | 2/2018 | Serratia marcescens | No | nt | nt |
| 7 | OUE1 | 3/2018 | No ID | No | nt | nt |
| 8 | OUE2 | 3/2018 | No ID | No | nt | nt |
| 9 | OUE3 | 3/2018 | No ID | No | nt | nt |
| Season 2019 | ||||||
| 10 | OUF1 | 1/2019 | Pantoea agglomerans | No | nt | nt |
| 11 | OUF2 | 1/2019 | Pantoea ananatis | Yes | MT234397 | MT091024 |
| 12 | OUF3 | 1/2019 | Acinetobacter johnstonii | No | nt | nt |
| 13 | OUG1 | 2/2019 | Pantoea ananatis | Yes | MT234398 | MT091025 |
| 14 | OUG2 | 2/2019 | Serratia marcescens | No | nt | nt |
| 15 | OUG3 | 2/2019 | Enterobacter aerogenes | No | nt | nt |
| 16 | OUH1 | 3/2019 | Acinetobacter johnstonii | No | nt | nt |
| 17 | OUH2 | 3/2019 | No ID | No | nt | nt |
| 18 | OUH3 | 3/2019 | No ID | No | nt | nt |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krawczyk, K.; Wielkopolan, B.; Obrępalska-Stęplowska, A. Pantoea ananatis, A New Bacterial Pathogen Affecting Wheat Plants (Triticum L.) in Poland. Pathogens 2020, 9, 1079. https://doi.org/10.3390/pathogens9121079
Krawczyk K, Wielkopolan B, Obrępalska-Stęplowska A. Pantoea ananatis, A New Bacterial Pathogen Affecting Wheat Plants (Triticum L.) in Poland. Pathogens. 2020; 9(12):1079. https://doi.org/10.3390/pathogens9121079
Chicago/Turabian StyleKrawczyk, Krzysztof, Beata Wielkopolan, and Aleksandra Obrępalska-Stęplowska. 2020. "Pantoea ananatis, A New Bacterial Pathogen Affecting Wheat Plants (Triticum L.) in Poland" Pathogens 9, no. 12: 1079. https://doi.org/10.3390/pathogens9121079
APA StyleKrawczyk, K., Wielkopolan, B., & Obrępalska-Stęplowska, A. (2020). Pantoea ananatis, A New Bacterial Pathogen Affecting Wheat Plants (Triticum L.) in Poland. Pathogens, 9(12), 1079. https://doi.org/10.3390/pathogens9121079