Dynamics of Classical Swine Fever Spread in Wild Boar in 2018–2019, Japan

 and
and
Abstract
1. Introduction
2. Results
2.1. Temporal Trend of CSF Cases in Wild Boar
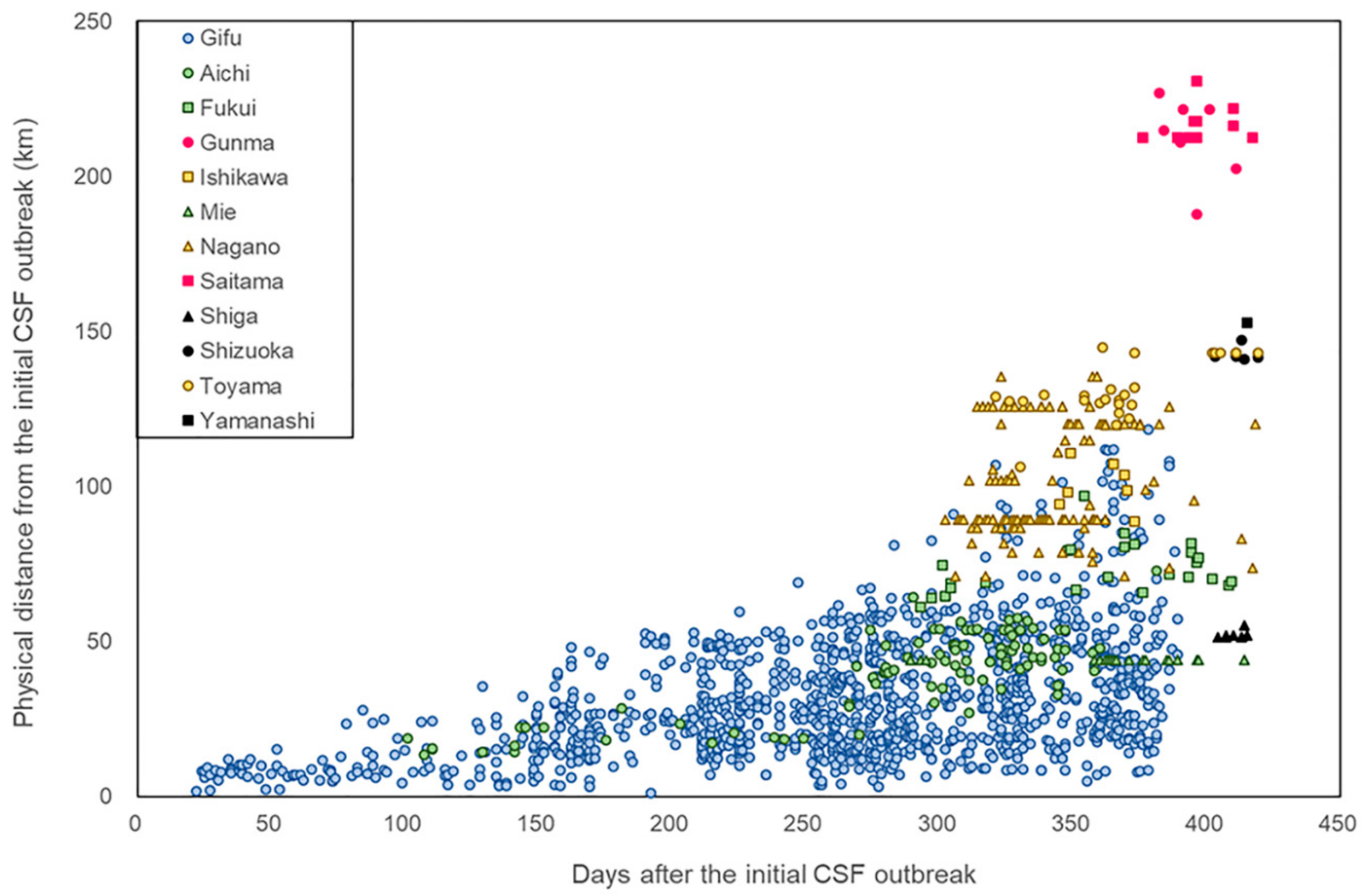
2.2. Distance of CSF Cases in Wild Boar from the Initial Outbreak Point
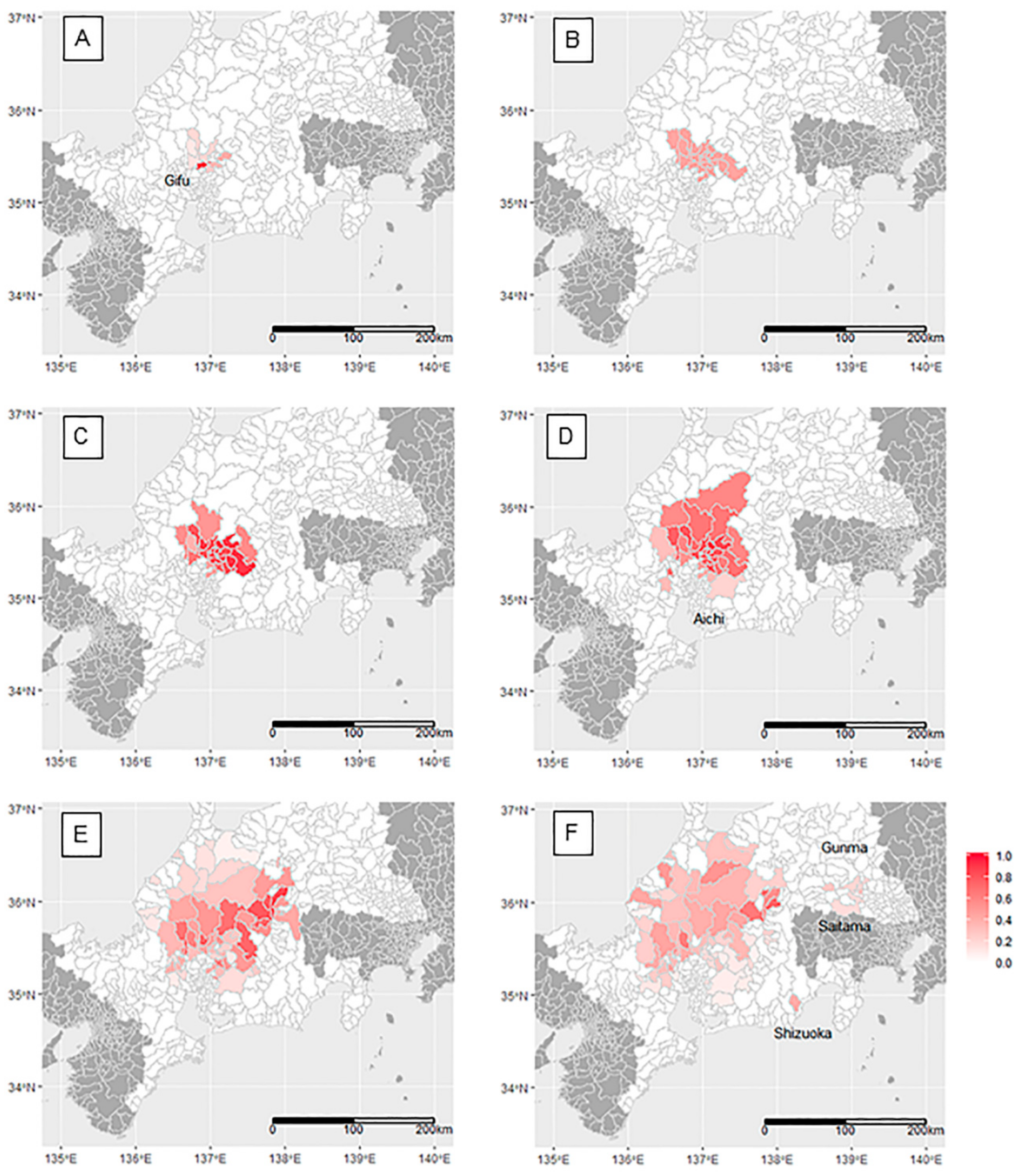
2.3. Spatial Change of CSF Period Prevalence Over Time
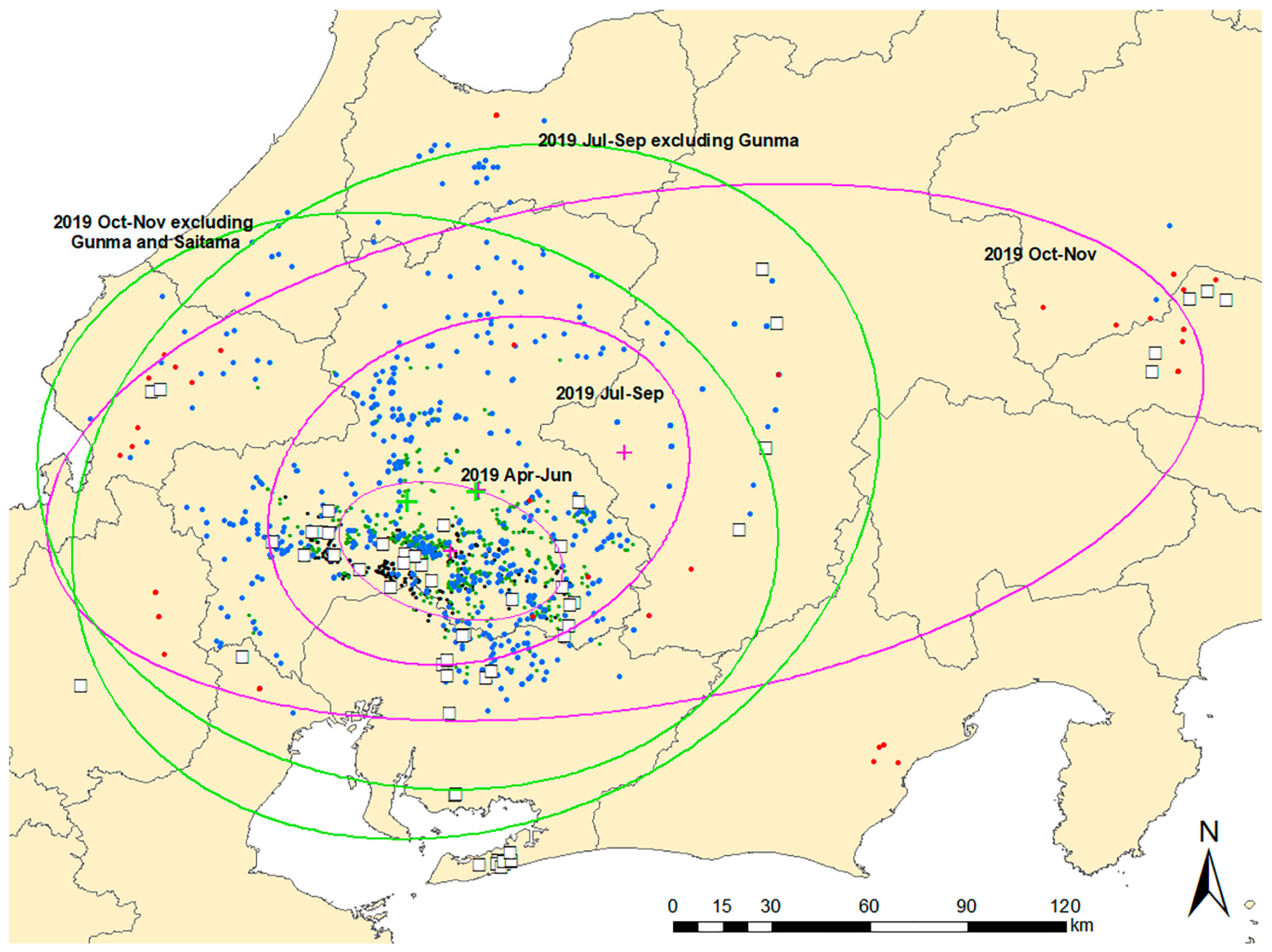
2.4. Standard Deviational Ellipse Analysis
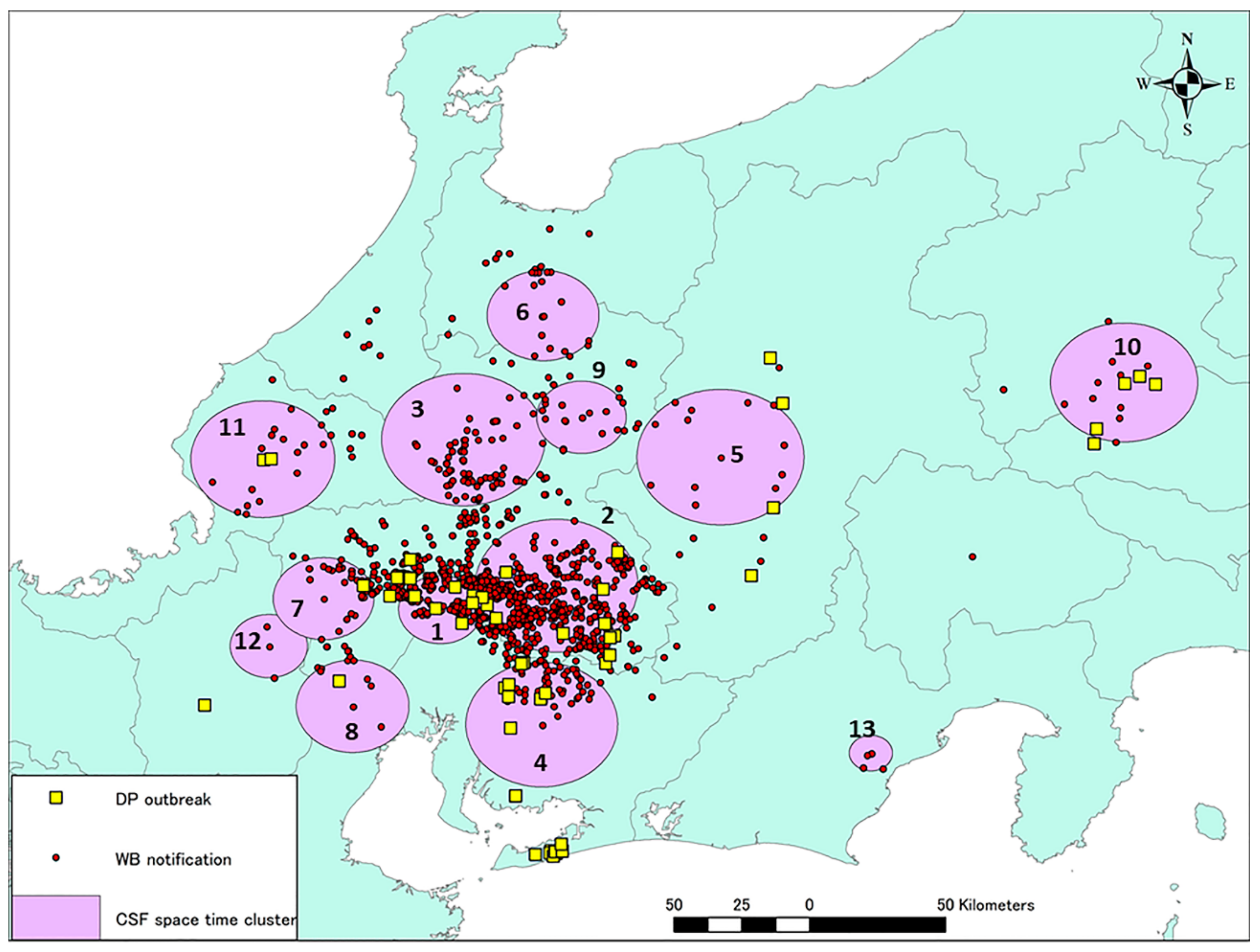
2.5. Space–Time Cluster Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Data and Data Sources
5.2. Temporal Trend and Linear Distance of CSF Cases in Wild Boar from the Initial Case
5.3. Description of Spatial Change of CSF Prevalence Over Time
5.4. SDE Analysis
5.5. Multi-Distance Spatial Cluster Analysis and Space–Time Cluster Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Edwards, S.; Fukusho, A.; Lefevre, P.C.; Lipowski, A.; Pejsak, Z.; Roehe, P.; Westergaard, J. Classical swine fever: The global situation. Vet. Microbiol. 2000, 73, 103–119. [Google Scholar] [CrossRef]
- Enkhbold, B.; Shatar, M.; Wakamori, S.; Tamura, T.; Hiono, T.; Matsuno, K.; Okamatsu, M.; Umemura, T.; Damdinjav, B.; Sakoda, Y. Genetic and virulence characterization of classical swine fever viruses isolated in Mongolia from 2007 to 2015. Virus Genes 2017, 53, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Ji, S.; Liu, Y.; Lei, J.L.; Xia, S.L.; Wang, Y.; Du, M.L.; Shao, L.; Meng, X.Y.; Zhou, M.; et al. Isolation and Characterization of a Moderately Virulent Classical Swine Fever Virus Emerging in China. Transbound. Emerg. Dis. 2017, 64, 1848–1857. [Google Scholar] [CrossRef] [PubMed]
- Kameyama, K.I.; Nishi, T.; Yamada, M.; Masujin, K.; Morioka, K.; Kokuho, T.; Fukai, K. Experimental infection of pigs with a classical swine fever virus isolated in Japan for the first time in 26 years. J. Vet. Med. Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Jurado, C.; Bosch, J.; Ito, M.; Sanchez-Vizcaino, J.M.; Isoda, N.; Sakoda, A.Y. Role of Wild Boar in the Spread of Classical Swine Fever in Japan. Pathogens 2019, 8, 206. [Google Scholar] [CrossRef]
- Ministry of Agriculture, Forestry and Fisheries, Japan (MAFF). Update of Classical Swine Fever in Japan. Available online: http://www.maff.go.jp/j/syouan/douei/csf/index.html (accessed on 10 February 2020).
- OIE. World Animal Health Information System. Available online: http://www.oie.int/wahis_2/public/wahid.php/Diseaseinformation/Diseasetimelines (accessed on 10 February 2020).
- International Steering Committee for Global Mapping. Global Map Version 1.2.1. Specifications. Available online: https://www.geospatial.jp/ckan/dataset/a5424d4b-dba5-4e68-83c3-0a6c29faf734/resource/a8f375d5-ed82-4fca-b77d-19205a972960/download/1.x.pdf (accessed on 10 February 2020).
- Hayama, Y.; Shimizu, Y.; Murato, Y.; Sawai, K.; Yamamoto, T. Estimation of infection risk on pig farms in infected wild boar areas-Epidemiological analysis for the reemergence of classical swine fever in Japan in 2018. Prev. Vet. Med. 2019, 175, 104873. [Google Scholar] [CrossRef] [PubMed]
- Moennig, V. The control of classical swine fever in wild boar. Front. Microbiol. 2015, 6, 1211. [Google Scholar] [CrossRef]
- Fritzemeier, J.; Teuffert, J.; Greiser-Wilke, I.; Staubach, C.; Schluter, H.; Moennig, V. Epidemiology of classical swine fever in Germany in the 1990s. Vet. Microbiol. 2000, 77, 29–41. [Google Scholar] [CrossRef]
- Rossi, S.; Fromont, E.; Pontier, D.; Cruciere, C.; Hars, J.; Barrat, J.; Pacholek, X.; Artois, M. Incidence and persistence of classical swine fever in free-ranging wild boar (Sus scrofa). Epidemiol. Infect. 2005, 133, 559–568. [Google Scholar] [CrossRef]
- Artois, M.; Depner, K.R.; Guberti, V.; Hars, J.; Rossi, S.; Rutili, D. Classical swine fever (hog cholera) in wild boar in Europe. Revue Scientifique et Technique 2002, 21, 287–303. [Google Scholar] [CrossRef]
- Rossi, S.; Staubach, C.; Blome, S.; Guberti, V.; Thulke, H.H.; Vos, A.; Koenen, F.; Le Potier, M.F. Controlling of CSFV in European wild boar using oral vaccination: A review. Front. Microbiol. 2015, 6, 1141. [Google Scholar] [CrossRef]
- Kaden, V.; Lange, E.; Fischer, U.; Strebelow, G. Oral immunisation of wild boar against classical swine fever: Evaluation of the first field study in Germany. Vet. Microbiol. 2000, 73, 239–252. [Google Scholar] [CrossRef]
- Rossi, S.; Pol, F.; Forot, B.; Masse-Provin, N.; Rigaux, S.; Bronner, A.; Le Potier, M.F. Preventive vaccination contributes to control classical swine fever in wild boar (Sus scrofa sp.). Vet. Microbiol. 2010, 142, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Hone, J.; Pech, R.; Yip, P. Estimation of the dynamics and rate of transmission of classical swine fever (hog cholera) in wild pigs. Epidemiol. Infect. 1992, 108, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Inayatullah, C. Wild boar in West Pakistan; Pakistan Forest Institute, Peshawar, Division of Forestry Research, Wildlife Management Branch: Peshawar, Pakistan, 1973; 17p. [Google Scholar]
- Ministry of Agriculture, Forestry and Fisheries, Japan (MAFF). Summary of 1st Meeting of Oral Vaccination to Classical Swine Fever (In Japanese). Available online: http://www.maff.go.jp/j/syouan/douei/csf/attach/pdf/domestic-26.pdf. (accessed on 10 February 2020).
- Takao, Y. Wild Boars and the Protection of Farm Crops at the Foot of Mt. Hakusan in Gifu Prefecture, Japan. Res. Rep. Hakusan Nat. Conserv. Cent. Ishikawa 1997, 24, 57–66. (In Japanese) [Google Scholar]
- Mauget, R. Seasonality of Reproduction in the Wild Boar; Butterworth Scientific: London, UK, 1982; pp. 509–526. [Google Scholar]
- Kodera, Y.; Takeda, T.; Tomaru, S.; Sugita, S. The estimation of birth periods in wild boar by detailed aging. Mamm. Sci. 2012, 52, 185–191. (In Japanese) [Google Scholar]
- Shimizu, Y.; Furuuchi, S.; Kumagai, T.; Sasahara, J. A mutant of hog cholera virus inducing interference in swine testicle cell cultures. Am. J. Vet. Res. 1970, 31, 1787–1794. [Google Scholar]
- Vilcek, S.; Herring, A.J.; Herring, J.A.; Nettleton, P.F.; Lowings, J.P.; Paton, D.J. Pestiviruses isolated from pigs, cattle and sheep can be allocated into at least three genogroups using polymerase chain reaction and restriction endonuclease analysis. Arch. Virol. 1994, 136, 309–323. [Google Scholar] [CrossRef]
- Bivand, R.S.; Gomez-Rubio, V.; Rue, H. Spatial Data Analysis with R-INLA with Some Extensions. J. Stat. Softw. 2015, 63, 1–31. [Google Scholar] [CrossRef]
- Fonseca, O.; Coronado, L.; Amaran, L.; Perera, C.L.; Centelles, Y.; Montano, D.N.; Alfonso, P.; Fernandex, O.; Santoro, K.R.; Frias-Lepoureau, M.T.; et al. Descriptive epidemiology of endemic Classical Swine Fever in Cuba. Span. J. Agric. Res. 2018, 16, e0508. [Google Scholar] [CrossRef]
- Lu, Y.; Deng, X.J.; Chen, J.H.; Wang, J.Y.; Chen, Q.; Niu, B. Risk analysis of African swine fever in Poland based on spatio-temporal pattern and Latin hypercube sampling, 2014–2017. BMC Vet. Res. 2019, 15, 160. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, I.; Rodriguez, A.; Feliziani, F.; Rolesu, S.; de la Torre, A. Spatio-temporal Analysis of African Swine Fever in Sardinia (2012-2014): Trends in Domestic Pigs and Wild Boar. Transbound. Emerg. Dis. 2017, 64, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Kulldorff, M.; Heffernan, R.; Hartman, J.; Assuncao, R.; Mostashari, F. A space-time permutation scan statistic for disease outbreak detection. PLoS Med. 2005, 2, e59. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster | Duration (Days) | Start Date | End Date | Radius (km) | Main Land Covers * |
|---|---|---|---|---|---|
| 1 | 148 | 2018/9/9 | 2019/2/4 | 12.53 | Closed shrublands, Cropland/Natural vegetation mosaic |
| 2 | 132 | 2019/2/12 | 2019/6/24 | 24.41 | Mixed forest |
| 3 | 69 | 2019/6/25 | 2019/9/2 | 24.33 | Mixed forest, Deciduous broadleaf forest, Cropland/Natural vegetation mosaic |
| 4 | 48 | 2019/7/2 | 2019/8/19 | 22.88 | Mixed forest, Deciduous broadleaf forest |
| 5 | 48 | 2019/7/16 | 2019/9/2 | 24.89 | Cropland/Natural vegetation mosaic, Mixed forest |
| 6 | 13 | 2019/9/3 | 2019/9/16 | 16.59 | Deciduous broadleaf forest |
| 7 | 27 | 2019/9/3 | 2019/9/30 | 15.11 | Mixed forest, Deciduous broadleaf forest |
| 8 | 55 | 2019/9/3 | 2019/10/28 | 16.99 | Closed shrublands, Mixed forest, Croplands |
| 9 | 20 | 2019/9/3 | 2019/9/23 | 13.30 | Cropland/Natural vegetation mosaic, Mixed forest |
| 10 | 41 | 2019/10/1 | 2019/11/11 | 21.87 | Closed shrublands, Croplands, Mixed forest |
| 11 | 27 | 2019/10/1 | 2019/10/28 | 21.46 | Croplands, Deciduous broadleaf forest |
| 12 | 20 | 2019/10/15 | 2019/11/4 | 11.61 | Water bodies, Croplands, Mixed forest |
| 13 | 20 | 2019/10/15 | 2019/11/4 | 6.52 | Mixed forest |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isoda, N.; Baba, K.; Ito, S.; Ito, M.; Sakoda, Y.; Makita, K. Dynamics of Classical Swine Fever Spread in Wild Boar in 2018–2019, Japan. Pathogens 2020, 9, 119. https://doi.org/10.3390/pathogens9020119
Isoda N, Baba K, Ito S, Ito M, Sakoda Y, Makita K. Dynamics of Classical Swine Fever Spread in Wild Boar in 2018–2019, Japan. Pathogens. 2020; 9(2):119. https://doi.org/10.3390/pathogens9020119
Chicago/Turabian StyleIsoda, Norikazu, Kairi Baba, Satoshi Ito, Mitsugi Ito, Yoshihiro Sakoda, and Kohei Makita. 2020. "Dynamics of Classical Swine Fever Spread in Wild Boar in 2018–2019, Japan" Pathogens 9, no. 2: 119. https://doi.org/10.3390/pathogens9020119
APA StyleIsoda, N., Baba, K., Ito, S., Ito, M., Sakoda, Y., & Makita, K. (2020). Dynamics of Classical Swine Fever Spread in Wild Boar in 2018–2019, Japan. Pathogens, 9(2), 119. https://doi.org/10.3390/pathogens9020119