Therapeutic Effects of Atranorin towards the Proliferation of Babesia and Theileria Parasites

 , , ,
, , ,
Abstract
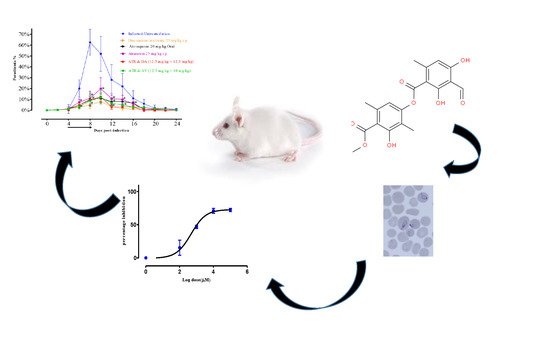
:
1. Introduction
2. Results
2.1. The Inhibition Assay of ATR In Vitro
2.2. In Vitro Potential of the Combination of ATR with DA, CF, and AV
2.3. Toxicity of ATR on Normal Cell Lines
2.4. The In Vivo Chemotherapeutic Potential of ATR in Mice
3. Discussion
4. Materials and Methods
4.1. Chemical Reagents
4.2. Ethical Approval
4.3. Cultivation Condition In Vitro
4.3.1. The Parasites Cultivation In Vitro
4.3.2. Evaluation of the Impacts of ATR on RBCs of Bovines and Equines
4.3.3. The In Vitro Growth Inhibitory Effects of ATR and Its Combination Efficacy with DA, CF, and AV
4.3.4. Parasite Viability Test In Vitro
4.4. Cytotoxicity Assay
4.4.1. Cell Cultures
4.4.2. Cytotoxic Action of ATR, DA, CF, and AV on Normal Cells
4.5. In Vivo Experiments
4.5.1. Chemotherapeutic Effects of ATR against B. Microti
4.5.2. PCR Identification of B. Microti DNA Extracted from All Treated Groups
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Homer, M.J.; Aguilar-Delfin, I.; Telford, S.R.; Krause, P.J.; Persing, D.H. Babesiosis. Clin. Microbiol. Rev. 2000, 13, 451–469. [Google Scholar] [CrossRef] [PubMed]
- Mosqueda, J.; Olvera-Ramirez, A.; Aguilar-Tipacamu, G.; Canto, G.J. Current Advances in Detection and Treatment of Babesiosis. Curr. Med. Chem. 2012, 19, 1504–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajapakshage, B.K.W.; Yamasaki, M.; Hwang, S.-J.; Sasaki, N.; Murakami, M.; Tamura, Y.; Lim, S.Y.; Nakamura, K.; Ohta, H.; Takiguchi, M. Involvement of mitochondrial genes of Babesia gibsoni in resistance to diminazene aceturate. J. Veter. Med. Sci. 2012, 74, 1139–1148. [Google Scholar] [CrossRef] [Green Version]
- A Hines, S.; Ramsay, J.D.; Kappmeyer, L.S.; Lau, A.O.; Ojo, K.K.; Van Voorhis, W.C.; Knowles, D.P.; Mealey, R.H. Theileria equi isolates vary in susceptibility to imidocarb dipropionate but demonstrate uniform in vitro susceptibility to a bumped kinase inhibitor. Parasites Vectors 2015, 8, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krause, P.J.; Lepore, T.; Sikand, V.K.; Gadbaw, J.; Brassard, P.; Pearl, D.; Azlanzadeh, J.; Christianson, D.; Telford, S.R.; Spielman, A.; et al. Atovaquone and Azithromycin for the Treatment of Babesiosis. N. Engl. J. Med. 2000, 343, 1454–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, D.A.; Urban, S. Phytochemical investigation of the Australian lichens Ramalina glaucescens and Xanthoria parietina. Nat. Prod. Commun. 2009, 4, 959–964. [Google Scholar] [CrossRef] [Green Version]
- Węgrzyn, M.H.; Wietrzyk-Pełka, P.; Galanty, A.; Cykowska-Marzencka, B.; Sundset, M.A. Incomplete degradation of lichen usnic acid and atranorin in Svalbard reindeer (Rangifer tarandus platyrhynchus). Polar Res. 2019, 38, 38. [Google Scholar] [CrossRef]
- De Melo, M.G.D.; Araújo, A.A.D.S.; Serafini, M.R.; Carvalho, L.F.; Bezerra, M.S.; Ramos, C.S.; Bonjardim, L.R.; Albuquerque-Júnior, R.L.C.; Lima, J.T.; Siqueira, R.S.; et al. Anti-inflammatory and toxicity studies of atranorin extracted from Cladina kalbii Ahti in rodents. Braz. J. Pharm. Sci. 2011, 47, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Yang, Y.; Park, S.-Y.; Nguyen, T.T.; Seo, Y.-W.; Lee, K.H.; Lee, J.H.; Kim, K.K.; Hur, J.-S.; Kim, H. The lichen secondary metabolite atranorin suppresses lung cancer cell motility and tumorigenesis. Sci. Rep. 2017, 7, 8136. [Google Scholar] [CrossRef] [Green Version]
- Thadhani, V.M.; Mesaik, M.A.; Asif, M.; Karunaratne, V.; Choudhary, I.M. Immunomodulatory activities of some common lichen metabolites. Int. J. Pharm. Pharm. Sci. 2015, 7, 144–147. [Google Scholar]
- Ranković, B.; Mišić, M.; Sukdolak, S. The antimicrobial activity of substances derived from the lichens Physcia aipola, Umbilicaria polyphylla, Parmelia caperata, and Hypogymnia physodes. World J. Microbiol. Biotechnol. 2008, 24, 1239–1242. [Google Scholar] [CrossRef]
- Vu, T.H.; Le Lamer, A.-C.; Lalli, C.; Boustie, J.; Samson, M.; Dévéhat, F.L.-L.; Le Seyec, J. Depsides: Lichen Metabolites Active against Hepatitis C Virus. PLoS ONE 2015, 10, e0120405. [Google Scholar] [CrossRef] [PubMed]
- Zofou, D.; Kengne, A.B.O.; Tene, M.; Ngemenya, M.N.; Tane, P.; Titanji, V.P.K. In vitro antiplasmodial activity and cytotoxicity of crude extracts and compounds from the stem bark of Kigelia africana (Lam.) Benth (Bignoniaceae). Parasitol. Res. 2011, 108, 1383–1390. [Google Scholar] [CrossRef]
- Zofou, D.; Tene, M.; Tane, P.; Titanji, V.P. Antimalarial drug interactions of compounds isolated from Kigelia africana (Bignoniaceae) and their synergism with artemether, against the multidrug-resistant W2mef Plasmodium falciparum strain. Parasitol. Res. 2012, 110, 539–544. [Google Scholar] [CrossRef]
- Marante, F.J.T.; Castellano, A.G.; Rosas, F.E.; Aguiar, J.Q.; Barrera, J.B. Identification and quantitation of allelochemicals from the lichen Lethariella canariensis: Phytotoxicity and antioxidative activity. J. Chem. Ecol. 2003, 29, 2049–2071. [Google Scholar] [CrossRef]
- Plsíkova, J.; Štêpánková, J.; Kasparkova, J.; Brabec, V.; Bačkor, M.; Kožurková, M. Lichen secondary metabolites as DNA-interacting agents. Toxicol. Vitr. 2014, 28, 182–186. [Google Scholar] [CrossRef]
- Shukla, V.; Joshi, G.P.; Rawat, M.S.M. Lichens as a potential natural source of bioactive compounds: A review. Phytochem. Rev. 2010, 9, 303–314. [Google Scholar] [CrossRef]
- Rizk, M.A.; El-Sayed, S.A.E.-S.; Aboulaila, M.; Yokoyama, N.; Igarashi, I. Evaluation of the inhibitory effect of N-acetyl-L-cysteine on Babesia and Theileria parasites. Exp. Parasitol. 2017, 179, 43–48. [Google Scholar] [CrossRef]
- Salama, A.A.; AbouLaila, M.; Terkawi, M.A.; Mousa, A.; El-Sify, A.; Allaam, M.; Zaghawa, A.; Yokoyama, N.; Igarashi, I. Inhibitory effect of allicin on the growth of Babesia and Theileria equi parasites. Parasitol. Res. 2014, 113, 275–283. [Google Scholar] [CrossRef]
- El-Sayed, S.A.E.-S.; Rizk, M.A.; Yokoyama, N.; Igarashi, I. Evaluation of the in vitro and in vivo inhibitory effect of thymoquinone on piroplasm parasites. Parasites Vectors 2019, 12, 37. [Google Scholar] [CrossRef] [PubMed]
- Rizk, M.A.; Aboulaila, M.; El-Sayed, S.A.E.-S.; Guswanto, A.; Yokoyama, N.; Igarashi, I. Inhibitory effects of fluoroquinolone antibiotics on Babesia divergens and Babesia microti, blood parasites of veterinary and zoonotic importance. Infect. Drug Resist. 2018, 11, 1605–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batiha, G.E.-S.; Beshbishy, A.M.; Tayebwa, D.S.; Adeyemi, O.S.; Shaheen, H.; Yokoyama, N.; Igarashi, I. The effects of trans-chalcone and chalcone 4 hydrate on the growth of Babesia and Theileria. PLoS Neglected Trop. Dis. 2019, 13, e0007030. [Google Scholar] [CrossRef] [Green Version]
- Bork, S.; Yokoyama, N.; Matsuo, T.; Claveria, F.G.; Fujisaki, K.; Igarashi, I. Clotrimazole, ketoconazole, and clodinafop-propargyl inhibit the in vitro growth of Babesia bigemina and Babesia bovis (Phylum Apicomplexa). Parasitology 2003, 127, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Salama, A.A.; Aboulaila, M.; Moussa, A.A.; Nayel, M.A.; El-Sify, A.; Terkawi, M.A.; Hassan, H.Y.; Yokoyama, N.; Igarashi, I. Evaluation of in vitro and in vivo inhibitory effects of fusidic acid on Babesia and Theileria parasites. Veter- Parasitology 2013, 191, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Batiha, G.E.-S.; Beshbishy, A.M.; Tayebwa, D.S.; Adeyemi, O.S.; Yokoyama, N.; Igarashi, I. Evaluation of the inhibitory effect of ivermectin on the growth of Babesia and Theileria parasites in vitro and in vivo. Trop. Med. Heal. 2019, 47, 42. [Google Scholar] [CrossRef] [PubMed]
- Beshbishy, A.M.; Batiha, G.E.-S.; Yokoyama, N.; Igarashi, I. Ellagic acid microspheres restrict the growth of Babesia and Theileria in vitro and Babesia microti in vivo. Parasites Vectors 2019, 12, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tayebwa, D.S.; Tuvshintulga, B.; Guswanto, A.; Nugraha, A.B.; Batiha, G.E.-S.; Gantuya, S.; Rizk, M.A.; Vudriko, P.; Sivakumar, T.; Yokoyama, N.; et al. The effects of nitidine chloride and camptothecin on the growth of Babesia and Theileria parasites. Ticks Tick-borne Dis. 2018, 9, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Guswanto, A.; Nugraha, A.B.; Tuvshintulga, B.; Tayebwa, D.S.; Rizk, M.A.; Batiha, G.E.-S.; Gantuya, S.; Sivakumar, T.; Yokoyama, N.; Igarashi, I. 17-DMAG inhibits the multiplication of several Babesia species and Theileria equi on in vitro cultures, and Babesia microti in mice. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 104–111. [Google Scholar] [CrossRef]
- Susithra, E.; Meena, S.; Chamundeeswari, D.; Chekkara, R.; Varalakshmi, E. Molecular docking studies of lichen metabolites as malarial protein inhibitors: Plasmepsin II protease and dihydrofolate reductase. J. Chem. Pharm. Res. 2014, 6, 501–512. [Google Scholar]
- Perry, D.P.G.; Benn, N.B.; Brennan, M.H.; Burgess, N.J.; Ellis, E.J.; Galloway, G.; Lorimer, S.D.; Tangney, R.S. Antimicrobial, antiviral and cytotoxic activity of New Zeland lichens. Lichenologist. 1999, 31, 627–636. [Google Scholar] [CrossRef]
- Correché, E.R.; Enriz, R.D.; Piovano, M.; Garbarino, J.; Gómez-Lechón, M.J. Cytotoxic and apoptotic effects on hepatocytes of secondary metabolites obtained from lichens. Altern. Lab. Anim. 2004, 32, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Baldissera, M.D.; Grando, T.H.; Souza, C.F.; Cossetin, L.F.; Sagrillo, M.R.; Nascimento, K.; Da Silva, A.P.; Lana, D.F.D.; Da Silva, A.S.; Stefani, L.M.; et al. Nerolidol nanospheres increases its trypanocidal efficacy against Trypanosoma evansi: New approach against diminazene aceturate resistance and toxicity. Exp. Parasitol. 2016, 166, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Gaikwad, S.; Verma, N.; Sarma, B.O.; Behera, B.C. Growth promoting effects of some lichen metabolites on probiotic bacteria. J. Food Sci. Technol. 2014, 51, 2624–2631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lengauer, H.; Just, F.T.; Edelhofer, R.; Pfister, K. Tick infestation and the prevalence of Borrelia burgdorferi and Babesia divergens in cattle in Bavaria. Berl. und Munchener tierarztliche Wochenschr. 2006, 119, 335–341. [Google Scholar]
- Igarashi, I.; Njonge, F.K.; Kaneko, Y.; Nakamura, Y. Babesia bigemina: In vitro and in vivo effects of curdlan sulfate on the growth of parasites. Exp. Parasitol. 1998, 90, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Batiha, G.E.-S.; Beshbishy, A.M.; Tayebwa, D.S.; Shaheen, H.M.; Yokoyama, N.; Igarashi, I. Inhibitory effects of Syzygium aromaticum and Camellia sinensis methanolic extracts on the growth of Babesia and Theileria parasites. Ticks Tick-borne Dis. 2019, 10, 949–958. [Google Scholar] [CrossRef] [PubMed]
- Beshbishy, A.M.; Batiha, G.E.S.; Adeyemi, O.S.; Yokoyama, N.; Igarashi, I. Inhibitory effects of methanolic Olea europaea and acetonic Acacia laeta on growth of Babesia and Theileria. Asian Pac. J. Trop. Med. 2019, 12, 425. [Google Scholar]
- Batiha, G.E.S.; Beshbishy, A.M.; Tayebwa, D.S.; Shaheen, M.H.; Yokoyama, N.; Igarashi, I. Inhibitory effects of Uncaria tomentosa bark, Myrtus communis roots, Origanum vulgare leaves and Cuminum cyminum seeds extracts against the growth of Babesia and Theileria in vitro. Jap. J. Vet. Parasitol. 2018, 17, 1–13. [Google Scholar]
- Chou, T.-C. Drug Combination Studies and Their Synergy Quantification Using the Chou-Talalay Method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [Green Version]
- Batiha, G.-S.; Beshbishy, A.M.; Adeyemi, O.S.; Nadwa, E.H.; Rashwan, E.M.; Alkazmi, L.M.; Elkelish, A.A.; Igarashi, I. Phytochemical screening and antiprotozoal effects of the methanolic Berberis vulgaris and acetonic Rhus coriaria extracts. Molecules 2020, 25, 550. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parasite | IC50 (µM) a | |||
|---|---|---|---|---|
| ATR | DA | AV | CF | |
| B. bovis | 98.4 ± 4.2 | 0.35 ± 0.06 | 0.04 ± 0.00 | 8.24 ± 1.7 |
| B. bigemina | 64.5 ± 3.9 | 0.68 ± 0.09 | 0.70 ± 0.04 | 5.73 ± 1.9 |
| B. divergens | 45.2 ± 5.9 | 0.43 ± 0.05 | 0.04 ± 0.00 | 13.8 ± 4.3 |
| B. caballi | 46.6 ± 2.5 | 0.02 ± 0.00 | 0.10 ± 0.01 | 7.95 ± 1.8 |
| T. equi | 71.3 ± 2.7 | 0.71 ± 0.05 | 0.09 ± 0.06 | 2.88 ± 0.9 |
| Parasite | ATR | Negative Control | ||
|---|---|---|---|---|
| Concentration (µM) a | Viability | |||
| B. bovis | A1 | 24.6 | + | |
| A2 | 49.2 | + | ||
| A3 | 98.4 | + | + | |
| A4 | 196.8 | – | ||
| A5 | 393.6 | – | ||
| A6 | 590.4 | – | ||
| B. bigemina | A1 | 16.1 | + | |
| A2 | 32.25 | + | ||
| A3 | 64.5 | + | + | |
| A4 | 129 | – | ||
| A5 | 258 | – | ||
| A6 | 387 | – | ||
| B. divergens | A1 | 11.3 | + | |
| A2 | 22.6 | + | ||
| A3 | 45.2 | + | + | |
| A4 | 90.4 | + | ||
| A5 | 180.8 | – | ||
| A6 | 271.2 | – | ||
| B. caballi | A1 | 11.65 | + | |
| A2 | 23.3 | + | ||
| A3 | 46.6 | + | + | |
| A4 | 93.2 | – | ||
| A5 | 186.4 | – | ||
| A6 | 279.6 | – | ||
| T. equi | A1 | 17.8 | + | |
| A2 | 35.6 | + | ||
| A3 | 71.3 | + | + | |
| A4 | 142.6 | + | ||
| A5 | 285.2 | + | ||
| A6 | 427.8 | – | ||
| Parasite | Drug Combination a | CI Values | Weighted Average CI Values b | Degree of Association c | |||
|---|---|---|---|---|---|---|---|
| IC50 | IC75 | IC90 | IC95 | ||||
| B. bovis | ATR + DA | 1.008 | 0.973 | 0.702 | 0.778 | 0.817 | Synergism |
| ATR + AV | 0.700 | 0.625 | 0.882 | 0.826 | 0.79 | Synergism | |
| ATR + CF | 0.783 | 0.398 | 0.892 | 0.919 | 0.793 | Synergism | |
| B. bigemina | ATR + DA | 0.806 | 0.994 | 0.933 | 0.938 | 0.937 | Synergism |
| ATR + AV | 0.903 | 1.007 | 1.003 | 1.91 | 1.356 | Additive | |
| ATR + CF | 0.719 | 0.382 | 0.181 | 0.393 | 0.359 | Synergism | |
| B. divergens | ATR + DA | 0.774 | 0.957 | 0.879 | 0.807 | 0.855 | Synergism |
| ATR + AV | 0.822 | 0.972 | 0.938 | 0.632 | 0.81 | Synergism | |
| ATR + CF | 0.922 | 0.791 | 0.772 | 0.883 | 0.835 | Synergism | |
| B. caballi | ATR + DA | 1.627 | 1.098 | 0.795 | 0.968 | 1.008 | Additive |
| ATR + AV | 0.482 | 0.052 | 0.372 | 0.572 | 0.399 | Synergism | |
| ATR + CF | 0.793 | 0.892 | 0.793 | 0.872 | 0.844 | Synergism | |
| T. equi | ATR + DA | 0.881 | 0.692 | 0.999 | 0.952 | 0.907 | Synergism |
| ATR + AV | 1.036 | 1.023 | 1.002 | 1.013 | 1.012 | Additive | |
| ATR + CF | 0.830 | 0.896 | 0.981 | 0.893 | 0.913 | Synergism | |
| Drug | Parasite | EC50 (µM) a | Selective Index b | ||||
|---|---|---|---|---|---|---|---|
| MDBK | NIH/3T3 | HFFs | MDBK | NIH/3T3 | HFFs | ||
| ATR | B. bovis | 775 ± 16 | 637.5 ± 12 | 750 ± 15 | 7.9 | 6.5 | 7.6 |
| B. bigemina | 775 ± 16 | 637.5 ± 12 | 750 ± 15 | 12.0 | 9.9 | 11.6 | |
| B. divergens | 775 ± 16 | 637.5 ± 12 | 750 ± 15 | 17.1 | 14.0 | 16.6 | |
| B. caballi | 775 ± 16 | 637.5 ± 12 | 750 ± 15 | 16.6 | 13.7 | 16.1 | |
| T. equi | 775 ± 16 | 637.5 ± 12 | 750 ± 15 | 10.9 | 9 | 10.6 | |
| DA | B. bovis | ˃100 | ˃100 | ˃100 | ˃285.7 | ˃285.7 | ˃285.7 |
| B. bigemina | ˃100 | ˃100 | ˃100 | ˃147.1 | ˃147.1 | ˃147.1 | |
| B. divergens | ˃100 | ˃100 | ˃100 | ˃232.5 | ˃232.5 | ˃232.5 | |
| B. caballi | ˃100 | ˃100 | ˃100 | ˃5000 | ˃5000 | ˃5000 | |
| T. equi | ˃100 | ˃100 | ˃100 | ˃140.8 | ˃140.8 | ˃140.8 | |
| AV | B. bovis | ˃100 | ˃100 | ˃100 | ˃2564 | ˃2564 | ˃2564 |
| B. bigemina | ˃100 | ˃100 | ˃100 | ˃142.7 | ˃142.7 | ˃142.7 | |
| B. divergens | ˃100 | ˃100 | ˃100 | ˃2631 | ˃2631 | ˃2631 | |
| B. caballi | ˃100 | ˃100 | ˃100 | ˃980.4 | ˃980.4 | ˃980.4 | |
| T. equi | ˃100 | ˃100 | ˃100 | ˃1052 | ˃1052 | ˃1052 | |
| CF | B. bovis | 34.7 ± 3.4 | ˃100 | ˃100 | 4.2 | ˃12.1 | ˃12.1 |
| B. bigemina | 34.7 ± 3.4 | ˃100 | ˃100 | 6.1 | ˃17.5 | ˃17.5 | |
| B. divergens | 34.7 ± 3.4 | ˃100 | ˃100 | 2.5 | ˃7.2 | ˃7.2 | |
| B. caballi | 34.7 ± 3.4 | ˃100 | ˃100 | 4.4 | ˃12.6 | ˃12.6 | |
| T. equi | 34.7 ± 3.4 | ˃100 | ˃100 | 12.1 | ˃34.7 | ˃34.7 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beshbishy, A.M.; Batiha, G.E.-S.; Alkazmi, L.; Nadwa, E.; Rashwan, E.; Abdeen, A.; Yokoyama, N.; Igarashi, I. Therapeutic Effects of Atranorin towards the Proliferation of Babesia and Theileria Parasites. Pathogens 2020, 9, 127. https://doi.org/10.3390/pathogens9020127
Beshbishy AM, Batiha GE-S, Alkazmi L, Nadwa E, Rashwan E, Abdeen A, Yokoyama N, Igarashi I. Therapeutic Effects of Atranorin towards the Proliferation of Babesia and Theileria Parasites. Pathogens. 2020; 9(2):127. https://doi.org/10.3390/pathogens9020127
Chicago/Turabian StyleBeshbishy, Amany Magdy, Gaber El-Saber Batiha, Luay Alkazmi, Eman Nadwa, Eman Rashwan, Ahmed Abdeen, Naoaki Yokoyama, and Ikuo Igarashi. 2020. "Therapeutic Effects of Atranorin towards the Proliferation of Babesia and Theileria Parasites" Pathogens 9, no. 2: 127. https://doi.org/10.3390/pathogens9020127