Tools for Molecular Epidemiology of Streptococcus suis



Abstract
:1. Introduction
2. Multilocus Sequence Typing (MLST)
3. Prediction of CC Using Polymerase Chain Reaction
4. Whole-Genome Sequencing Approaches
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goyette-Desjardins, G.; Auger, J.-P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent-an update on the worldwide distribution based on serotyping and sequence typing. Emerg. Microbes. Infect. 2014, 3, e45. [Google Scholar] [CrossRef] [PubMed]
- Hlebowicz, M.; Jakubowski, P.; Smiatacz, T. Streptococcus suis Meningitis: Epidemiology, Clinical Presentation and Treatment. Vector Borne Zoonotic Dis. 2019, 19, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Huong, V.T.L.; Ha, N.; Huy, N.T.; Horby, P.; Nghia, H.D.T.; Thiem, V.D.; Zhu, X.; Hoa, N.T.; Hien, T.T.; Zamora, J.; et al. Epidemiology, Clinical manifestations, and outcomes of Streptococcus suis infection in Humans. Emerg. Infect. Dis. 2014, 20, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Dutkiewicz, J.; Sroka, J.; Zając, V.; Wasiński, B.; Cisak, E.; Sawczyn, A.; Kloc, A.; Wójcik-Fatla, A. Streptococcus suis: A re-emerging pathogen associated with occupational exposure to pigs or pork products. Part I—Epidemiology. Ann. Agric. Environ. Med. 2017, 24, 683–695. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, M.; Higgins, R.; Jacques, M.; Mittal, K.R.; Henrichsen, J. Description of 14 new capsular types of Streptococcus suis. J. Clin. Microbiol. 1989, 27, 2633–2636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottschalk, M.; Higgins, R.; Jacques, M.; Beaudoin, M.; Henrichsen, J. Characterization of six new capsular types (23 through 28) of Streptococcus suis. J. Clin. Microbiol. 1991, 29, 2590–2594. [Google Scholar] [CrossRef] [Green Version]
- Gottschalk, M.; Higgins, R.; Jacques, M.; Beaudoin, M.; Henrichsen, J. Isolation and characterization of Streptococcus suis capsular types 9–22. J. Vet. Diagn. Investig. 1991, 3, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Higgins, R.; Gottschalk, M.; Boudreau, M.; Lebrun, A.; Henrichsen, J. Description of Six New Capsular Types (29–34) of Streptococcus Suis. J. Vet. Diagn. Investig. 1995, 7, 405–406. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.E.; Gottschalk, M.; Brousseau, R.; Harel, J.; Hemmingsen, S.M.; Goh, S.H. Biochemical analysis, cpn60 and 16S rDNA sequence data indicate that Streptococcus suis serotypes 32 and 34, isolated from pigs, are Streptococcus orisratti. Vet. Microbiol. 2005, 107, 63–69. [Google Scholar] [CrossRef]
- Tien, L.H.T.; Nishibori, T.; Nishitani, Y.; Nomoto, R.; Osawa, R. Reappraisal of the taxonomy of Streptococcus suis serotypes 20, 22, 26, and 33 based on DNA–DNA homology and sodA and recN phylogenies. Vet. Microbiol. 2013, 162, 842–849. [Google Scholar] [CrossRef]
- Nomoto, R.; Maruyama, F.; Ishida, S.; Tohya, M.; Sekizaki, T.; Osawa, R. Reappraisal of the taxonomy of Streptococcus suis serotypes 20, 22 and 26: Streptococcus parasuis sp. Int. J. Syst. Evol. Microbiol. 2015, 65, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Okura, M.; Osaki, M.; Nomoto, R.; Arai, S.; Osawa, R.; Sekizaki, T.; Takamatsu, D. Current Taxonomical Situation of Streptococcus suis. Pathogens 2016, 5, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tohya, M.; Arai, S.; Tomida, J.; Watanabe, T.; Kawamura, Y.; Katsumi, M.; Ushimizu, M.; Ishida-Kuroki, K.; Yoshizumi, M.; Uzawa, Y.; et al. Defining the taxonomic status of Streptococcus suis serotype 33: The proposal for Streptococcus ruminantium sp. nov. Int. J. Syst. Evol. Microbiol. 2017, 67, 3660–3665. [Google Scholar] [CrossRef] [PubMed]
- Hatrongjit, R.; Kerdsin, A.; Gottschalk, M.; Takeuchi, D.; Hamada, S.; Oishi, K.; Akeda, Y. First human case report of sepsis due to infection with Streptococcus suis serotype 31 in Thailand. BMC Infect. Dis. 2015, 15, 392. [Google Scholar] [CrossRef] [PubMed]
- Kerdsin, A.; Oishi, K.; Sripakdee, S.; Boonkerd, N.; Polwichai, P.; Nakamura, S.; Uchida, R.; Sawanpanyalert, P.; Dejsirilert, S. Clonal dissemination of Streptococcus suis serotype 14 in Thailand. J. Med. Microbiol. 2009, 58, 1508–1513. [Google Scholar] [CrossRef] [PubMed]
- Kerdsin, A.; Dejsirilert, S.; Sawanpanyalert, P.; Boonnark, A.; Noithachang, W.; Sriyakum, D.; Simkum, S.; Chokngam, S.; Gottschalk, M.; Akeda, Y.; et al. Sepsis and spontaneous bacterial peritonitis in Thailand. Lancet 2011, 378, 960. Available online: https://doi.org/10.1016/S0140-6736(11)60923-9 (accessed on 27 January 2020). [CrossRef]
- Kerdsin, A.; Hatrongjit, R.; Gottschalk, M.; Takeuchi, D.; Hamada, S.; Akeda, Y.; Oishi, K. Emergence of Streptococcus suis serotype 9 infection in humans. J. Microbiol. Immunol. Infect. 2017, 50, 545–546. [Google Scholar] [CrossRef] [Green Version]
- Berthelot-Hérault, F.; Marois, C.; Gottschalk, M.; Kobisch, M. Genetic diversity of Streptococcus suis strains isolated from pigs and humans as revealed by pulsed-field gel electrophoresis. J. Clin. Microbiol 2002, 40, 615–619. [Google Scholar] [CrossRef] [Green Version]
- Vela, A.I.; Goyache, J.; Tarradas, C.; Luque, I.; Mateos, A.; Moreno, M.A.; Borge, C.; Perea, J.A.; Domínguez, L.; Fernandez-Garayzabal, J.F. Analysis of Genetic Diversity of Streptococcus suis Clinical Isolates from Pigs in Spain by Pulsed-Field Gel Electrophoresis. J. Clin. Microbiol. 2003, 41, 2498–2502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatellier, S.; Gottschalk, M.; Higgins, R.; Brousseau, R.; Harel, J. Relatedness of Streptococcus suisSerotype 2 Isolates from Different Geographic Origins as Evaluated by Molecular Fingerprinting and Phenotyping. J. Clin. Microbiol. 1999, 37, 362–366. [Google Scholar] [CrossRef] [Green Version]
- Cloutier, G.; D’Allaire, S.; Martinez, G.; Surprenant, C.; Lacouture, S.; Gottschalk, M. Epidemiology of Streptococcus suis serotype 5 infection in a pig herd with and without clinical disease. Vet. Microbiol. 2003, 97, 135–151. [Google Scholar] [CrossRef] [PubMed]
- Rehm, T.; Baums, C.G.; Strommenger, B.; Beyerbach, M.; Valentin-Weigand, P.; Goethe, R. Amplified fragment length polymorphism of Streptococcus suis strains correlates with their profile of virulence-associated genes and clinical background. J. Med. Microbiol. 2007, 56, 102–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marois, C.; Le Devendec, L.; Gottschalk, M.; Kobisch, M. Molecular characterization of Streptococcus suis strains by 16S–23S intergenic spacer polymerase chain reaction and restriction fragment length polymorphism analysis. Can. J. Vet. Res. 2006, 70, 94–104. [Google Scholar]
- Harel, J.; Higgins, R.; Gottschalk, M.; Bigras-Poulin, M. Genomic relatedness among reference strains of different Streptococcus suis serotypes. Can. J. Vet. Res. 1994, 58, 259–262. [Google Scholar]
- Okwumabua, O.; Staats, J.; Chengappa, M.M. Detection of genomic heterogeneity in Streptococcus suis isolates by DNA restriction fragment length polymorphisms of rRNA genes (ribotyping). J. Clin. Microbiol. 1995, 33, 968–972. [Google Scholar] [CrossRef] [Green Version]
- King, S.J.; Leigh, J.A.; Heath, P.J.; Luque, I.; Tarradas, C.; Dowson, C.G.; Whatmore, A.M. Development of a Multilocus Sequence Typing Scheme for the Pig Pathogen Streptococcus suis: Identification of Virulent Clones and Potential Capsular Serotype Exchange. J. Clin. Microbiol. 2002, 40, 3671–3680. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Ye, C.; Jing, H.; Cui, Z.; Bai, X.; Jin, D.; Zheng, H.; Zhao, A.; Xu, Y.; Gottschalk, M.; et al. Streptococcus suis outbreak investigation using multiple-locus variable tandem repeat number analysis. Microbiol. Immunol. 2010, 54, 380–388. [Google Scholar] [CrossRef]
- Zheng, H.; Lan, R.; Zheng, X.; Cui, Z.; Liu, Z.; Bai, X.; Ji, S.; Gottschalk, M.; Xu, J. Comparative Genomic Hybridization Identifies Virulence Differences in Streptococcus suis. PLoS ONE 2014, 9, e87866. [Google Scholar] [CrossRef]
- Willemse, N.; Howell, K.J.; Weinert, L.A.; Heuvelink, A.; Pannekoek, Y.; Wagenaar, J.A.; Smith, H.E.; Van Der Ende, A.; Schultsz, C. An emerging zoonotic clone in the Netherlands provides clues to virulence and zoonotic potential of Streptococcus suis. Sci. Rep. 2016, 6, 28984. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Zhang, W.; Zheng, H.; Lan, R.; Wang, H.; Du, P.; Bai, X.; Ji, S.; Meng, Q.; Jin, N.; et al. Minimum core genome sequence typing of bacterial pathogens: A unified approach for clinical and public health microbiology. J. Clin. Microbiol. 2013, 51, 2582–2591. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Ji, S.; Lan, R.; Liu, Z.; Bai, X.; Zhang, W.; Gottschalk, M.; Xu, J. Population Analysis of Streptococcus suis Isolates from Slaughtered Swine by Use of Minimum Core Genome Sequence Typing. J. Clin. Microbiol. 2014, 52, 3568–3572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wileman, T.M.; Weinert, L.A.; Howell, K.J.; Wang, J.; Peters, S.E.; Williamson, S.M.; Wells, J.M.; Langford, P.R.; Rycroft, A.N.; Wren, B.W.; et al. Pathotyping the Zoonotic Pathogen Streptococcus suis: Novel Genetic Markers to Differentiate Invasive Disease-Associated Isolates from Non-Disease-Associated Isolates from England and Wales. J. Clin. Microbiol. 2019, 57, e01712-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerdsin, A.; Akeda, Y.; Takeuchi, D.; Dejsirilert, S.; Gottschalk, M.; Oishi, K. Genotypic diversity of Streptococcus suis strains isolated from humans in Thailand. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-I.; Jeon, A.B.; Jung, B.-Y.; Byun, J.-W.; Gottschalk, M.; Kim, A.; Kim, J.W.; Kim, H.-Y. Capsular serotypes, virulence-associated genes and antimicrobial susceptibility of Streptococcus suis isolates from pigs in Korea. J. Vet. Med. Sci. 2017, 79, 780–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultsz, C.; Jansen, E.; Keijzers, W.; Rothkamp, A.; Duim, B.; Wagenaar, J.A.; Van Der Ende, A. Differences in the Population Structure of Invasive Streptococcus suis Strains Isolated from Pigs and from Humans in the Netherlands. PLoS ONE 2012, 7, e33854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groves, M.D.; Jordan, D.; Chapman, T.A.; Al Jassim, R. Multilocus sequence typing of Australian Streptococcus suis type 2 by MALDI-TOF mass spectrometry analysis of PCR amplicons. Vet. Microbiol. 2015, 177, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Tark, D.; Moon, S.-H.; Han, J.-I.; Kim, W.-I.; Cho, H.-S. Sepsis Caused by Streptococcus suis Serotype 2 in a Eurasian River Otter (Lutra lutra) in the Republic of Korea. J. Wildl. Dis. 2018, 54, 866–869. [Google Scholar] [CrossRef]
- Estrada, A.A.; Gottschalk, M.; Rossow, S.; Rendahl, A.; Gebhart, C.; Marthaler, D.G. Serotype and Genotype (Multilocus Sequence Type) of Streptococcus suis Isolates from the United States Serve as Predictors of Pathotype. J. Clin. Microbiol. 2019, 57, e00377-19. [Google Scholar] [CrossRef] [Green Version]
- Takamatsu, D.; Osaki, M.; Tharavichitkul, P.; Takai, S.; Sekizaki, T. Allelic variation and prevalence of serum opacity factor among the Streptococcus suis population. J. Med. Microbiol. 2008, 57, 488–494. [Google Scholar] [CrossRef]
- Takamatsu, D.; Nishino, H.; Ishiji, T.; Ishii, J.; Osaki, M.; Fittipaldi, N.; Gottschalk, M.; Tharavichitkul, P.; Takai, S.; Sekizaki, T. Genetic organization and preferential distribution of putative pilus gene clusters in Streptococcus suis. Vet. Microbiol. 2009, 138, 132–139. [Google Scholar] [CrossRef]
- Hatrongjit, R.; Kerdsin, A.; Gottschalk, M.; Hamada, S.; Oishi, K.; Akeda, Y. Development of a multiplex PCR assay to detect the major clonal complexes of Streptococcus suis relevant to human infection. J. Med. Microbiol. 2016, 65, 392–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maneerat, K.; Yongkiettrakul, S.; Kramomtong, I.; Tongtawe, P.; Tapchaisri, P.; Luangsuk, P.; Chaicumpa, W.; Gottschalk, M.; Srimanote, P. Virulence Genes and Genetic Diversity ofStreptococcus suisSerotype 2 Isolates from Thailand. Transbound. Emerg. Dis. 2013, 60, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Kidchana, A.; Meekhanon, N.; Hatrongjit, R.; Gottschalk, M.; Kerdsin, A. Application of random amplified polymorphism DNA and 16S-23S rDNA intergenic spacer polymerase chain reaction-restriction fragment length polymorphism to predict major Streptococcus suis clonal complexes isolated from humans and pigs. Mol. Cell. Probes 2019, 43, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Athey, T.B.T.; Teatero, S.; Lacouture, S.; Takamatsu, D.; Gottschalk, M.; Fittipaldi, N. Determining Streptococcus suis serotype from short-read whole-genome sequencing data. BMC Microbiol. 2016, 16, 162. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Wang, M.; Hao, H.; Yang, R.; Xie, J.; Su, J.; Lin, M.; Cui, Y.; Jiang, Y. Genomic epidemiological investigation of a Streptococcus suis outbreak in Guangxi, China, 2016. Infect. Genet. Evol. 2019, 68, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Zheng, H.; Zhou, J.; Lan, R.; Ye, C.; Jing, H.; Jin, D.; Cui, Z.; Bai, X.; Liang, J.; et al. Detection of Multiple Parallel Transmission Outbreak of Streptococcus suis Human Infection by Use of Genome Epidemiology, China, 2005. Emerg. Infect Dis. 2017, 23, 204–211. [Google Scholar] [CrossRef] [Green Version]
- Willemse, N.; van der Ende, A.; Schultsz, C. Reinfection with Streptococcus suis analysed by whole genome sequencing. Zoonoses Public Health 2019, 66, 179–183. [Google Scholar] [CrossRef]
- Athey, T.B.; Teatero, S.; Takamatsu, D.; Wasserscheid, J.; Dewar, K.; Gottschalk, M.; Fittipaldi, N. Population Structure and Antimicrobial Resistance Profiles of Streptococcus suis Serotype 2 Sequence Type 25 Strains. PLoS ONE 2016, 11, e0150908. [Google Scholar] [CrossRef] [Green Version]
- Athey, T.B.T.; Auger, J.-P.; Teatero, S.; Dumesnil, A.; Takamatsu, D.; Wasserscheid, J.; Dewar, K.; Gottschalk, M.; Fittipaldi, N. Complex Population Structure and Virulence Differences among Serotype 2 Streptococcus suis Strains Belonging to Sequence Type 28. PLoS ONE 2015, 10, e0137760. [Google Scholar] [CrossRef]
- Zheng, H.; Du, P.; Qiu, X.; Kerdsin, A.; Roy, D.; Bai, X.; Xu, J.; Vela, A.I.; Gottschalk, M. Genomic comparisons of Streptococcus suis serotype 9 strains recovered from diseased pigs in Spain and Canada. Vet. Res. 2018, 49, 1. [Google Scholar] [CrossRef] [Green Version]
- Ruan, Z.; Feng, Y. BacWGSTdb, a database for genotyping and source tracking bacterial pathogens. Nucleic Acids Res. 2016, 44, D682–D687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Greeff, A.; Wisselink, H.J.; De Bree, F.M.; Schultsz, C.; Baums, C.G.; Thi, H.N.; Stockhofe-Zurwieden, N.; Smith, H.E. Genetic diversity of Streptococcus suis isolates as determined by comparative genome hybridization. BMC Microbiol. 2011, 11, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinert, L.A.; Chaudhuri, R.R.; Wang, J.; Peters, S.E.; Corander, J.; Jombart, T.; Baig, A.; Howell, K.J.; Vehkala, M.; Välimäki, N.; et al. Genomic signatures of human and animal disease in the zoonotic pathogen Streptococcus suis. Nat. Commun. 2015, 6, 6740. Available online: https://doi.org/10.1038/ncomms7740 (accessed on 27 January 2020). [CrossRef] [PubMed] [Green Version]
- Gomez-Torres, J.; Nimir, A.; Cluett, J.; Aggarwal, A.; Elsayed, S.; Soares, D.; Teatero, S.; Chen, Y.; Gottschalk, M.; Fittipaldi, N. Human Case of Streptococcus suis Disease, Ontario, Canada. Emerg. Infect. Dis. 2017, 23, 2107–2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, T.; Tsukagoshi, H.; Sekizuka, T.; Ishikawa, D.; Imai, M.; Fujita, M.; Kuroda, M.; Saruki, N. Next-generation DNA sequencing analysis of two Streptococcus suis ST28 isolates associated with human infective endocarditis and meningitis in Gunma, Japan: A case report. Infect. Dis. 2018, 51, 62–66. [Google Scholar] [CrossRef]
- Tan, S.; Dvorak, C.M.T.; Estrada, A.A.; Gebhart, C.; Marthaler, D.G.; Murtaugh, M.P. MinION sequencing of Streptococcus suis allows for functional characterization of bacteria by multilocus sequence typing and antimicrobial resistance profiling. J. Microbiol. Methods 2019, 169, 105817. [Google Scholar] [CrossRef]


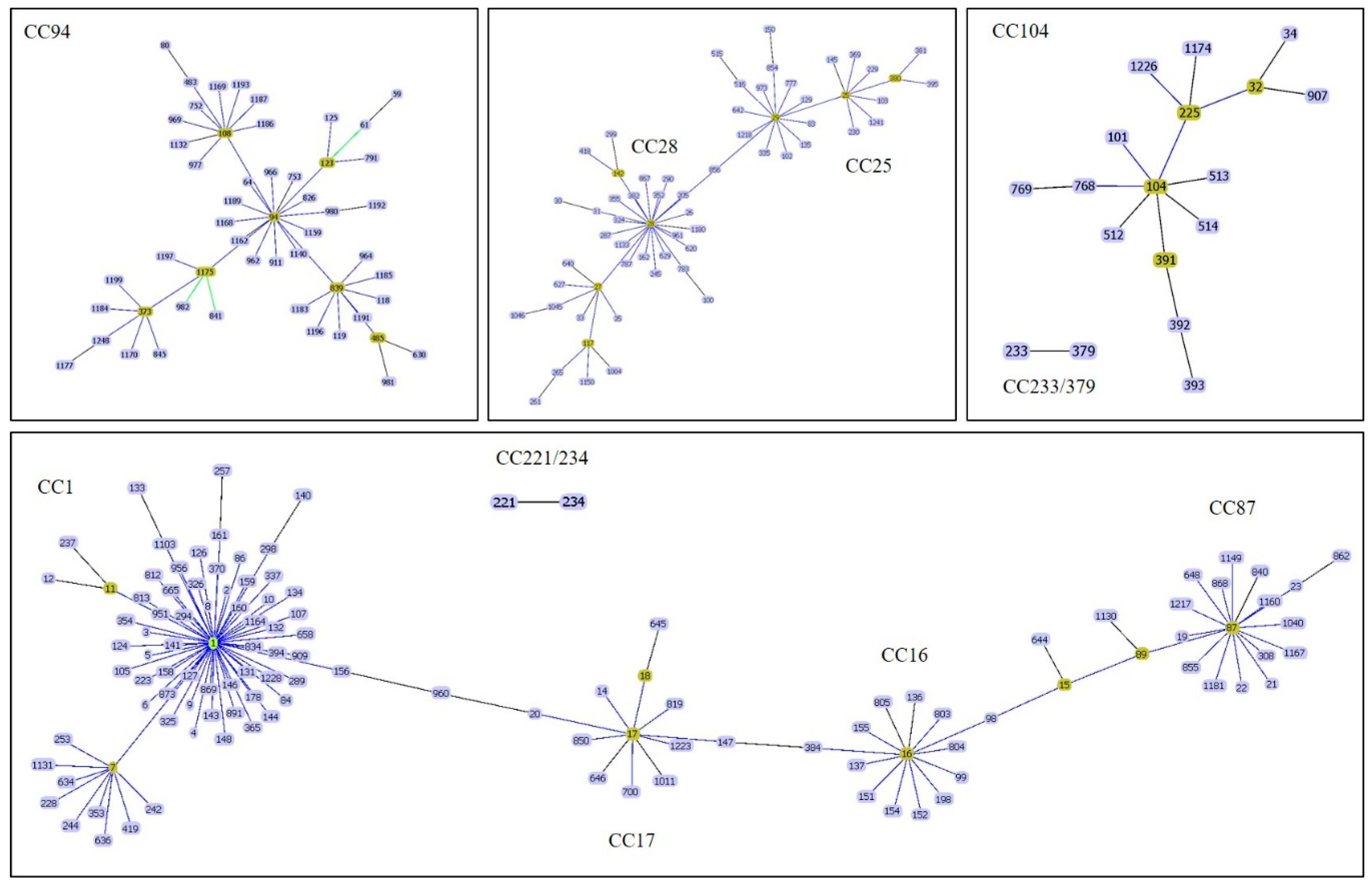{kind=link}
| Characteristic | WGS | MLST | Multiplex PCR-CC | RAPD | PCR-RFLP | MLVA | AFLP | PFGE | Ribotyping |
|---|---|---|---|---|---|---|---|---|---|
| Reproducibility | Good | Good | Good | Poor to moderate | Moderate | Good | Good | Good | Good |
| Discriminatory power | Excellent | High | Moderate | Moderate to good | Poor to moderate | Excellent | Excellent | Excellent | Good |
| Ease of use | Moderately labor-intensive | Simple to moderate labor | Simple | Simple | Simple | Simple | Moderate | Labor-intensive | Labor-intensive |
| Interpretation | Moderate to very complex | Simple to moderate | Simple | Moderate to complex | Simple | Simple | Complex | Moderate to complex | Moderate to complex |
| Cost | Very high | Moderate | Low | Low | Low | Low to moderate | Moderate | High | High |
| Universal applicability | Yes | Yes | Limit to some CCs | Yes | Yes | Yes | Yes | Yes | Yes |
| Clonal Complexes | PCR Methods | ||||
|---|---|---|---|---|---|
| Multiplex PCR | PCR of ofs Genes | PCR-Pilus-Associated Gene Profiles | RAPD | 16S-23S rDNA PCR-RFLP | |
| CC1 | 🗸 | 🗸 | 🗸 | 🗸 | 🗸 |
| CC16 | 🗶 | 🗶 | 🗶 | 🗸 | 🗸 |
| CC20 ** | 🗶 | 🗶 | 🗶 | 🗶 | 🗶 |
| CC25 | 🗸 | 🗸 * | 🗸 * | 🗸 | 🗸 * |
| CC28 | 🗸 | 🗸 * | 🗸 * | 🗸 | 🗸 * |
| CC94 | 🗶 | 🗸 * | 🗶 | 🗶 | 🗶 |
| CC104 | 🗸 | 🗸 * | 🗸 | 🗸 | 🗸 * |
| CC233/379 | 🗸 | 🗶 | 🗶 | 🗸 | 🗸 * |
| CC221/234 | 🗸 | 🗶 | 🗶 | 🗸 | 🗸 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hatrongjit, R.; Fittipaldi, N.; Gottschalk, M.; Kerdsin, A. Tools for Molecular Epidemiology of Streptococcus suis. Pathogens 2020, 9, 81. https://doi.org/10.3390/pathogens9020081
Hatrongjit R, Fittipaldi N, Gottschalk M, Kerdsin A. Tools for Molecular Epidemiology of Streptococcus suis. Pathogens. 2020; 9(2):81. https://doi.org/10.3390/pathogens9020081
Chicago/Turabian StyleHatrongjit, Rujirat, Nahuel Fittipaldi, Marcelo Gottschalk, and Anusak Kerdsin. 2020. "Tools for Molecular Epidemiology of Streptococcus suis" Pathogens 9, no. 2: 81. https://doi.org/10.3390/pathogens9020081
APA StyleHatrongjit, R., Fittipaldi, N., Gottschalk, M., & Kerdsin, A. (2020). Tools for Molecular Epidemiology of Streptococcus suis. Pathogens, 9(2), 81. https://doi.org/10.3390/pathogens9020081