Isolation and Characterization of Root-Associated Bacterial Endophytes and Their Biocontrol Potential against Major Fungal Phytopathogens of Rice (Oryza sativa L.)

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Isolation and Identification of Root-Associated Bacterial Endophytes
2.2. Molecular Classification and 16S rRNA Gene Sequences Based Phylogeny Association of Root-Associated Bacterial Endophytes
2.3. Biochemical and Physiological Characterization of Root-Associated Bacterial Endophytes
2.4. In-vitro Antagonistic Activities of Root-Associated Bacterial Endophytes
2.5. Molecular Detection of Antibiotic Related Genes Based on PCR Amplification of Root-Associated Bacterial Endophytes
3. Discussion
4. Materials and Methods
4.1. Plant Material and Sampling Location
4.2. Surface Sterilization of Rice Root Tissues
4.3. Isolation of Root-Associated Bacterial Endophytes
4.4. Biochemical and Physiological Characterization of Root-Associated Bacterial Endophytes
4.5. Genomic DNA Extraction of Root-Associated Bacterial Endophytes
4.6. PCR Amplification and Classification of Root-Associated Bacterial Endophytes
4.7. 16S rRNA Gene Sequences Based Phylogeny Analysis of Root-Associated Bacterial Endophytes
4.8. Rice Fungal Phytopathogens and Culture Condition
4.9. Molecular Detection of Antibiotic Related Genes Based on PCR Amplification of Root-Associated Bacterial Endophytes
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PCR | = | Polymer Chain Reaction |
| C-1 | = | Cultivar One (Xiushui-48) |
| C-2 | = | Cultivar Two (Y-003) |
| C-3 | = | Cultivar Three (CO-39) |
| CNRRI | = | China National Rice Research Institute |
| LB | = | Luria Bertani |
| PDA | = | Potato Dextrose Agar |
| RPM | = | Rounds Per Mint |
| DGH | = | Disease Growth Habit |
| TAE | = | Tris-acetate EDTA |
| GR | = | Gram Reaction |
| CA | = | Catalase Activity |
| EA | = | Endospore Activity |
| OA | = | Oxidase Activity |
| MP | = | Motility Performance |
| CA | = | Citrate Activity |
| GP | = | Gelatinase Production |
| SHA | = | Starch Hydrolyses Activity |
| NRA | = | Nitrate Reduction Activity |
| VPP | = | Voges Proskauer’s Performance |
| MRA | = | Methyl Red Activity |
| GA | = | Glucose Activity |
| MA | = | Mannitol Activity |
| LA | = | Lactose Activity |
| MA | = | Maltose Activity |
| PLT | = | Pyoluteorin |
| PRN | = | Pyrrolnitrin |
| PKSI | = | Polyketide Synthase |
| NRPS | = | Non-ribosomal Peptide Synthetases |
| HCN | = | Hydrogen Cyanide |
| Sfp | = | Surfactin Biosynthesis |
| SrfC | = | Surfactin Synthase |
| ItuD | = | Iturin A Biosynthesis |
| FenD | = | Fengycin Biosynthesis |
| BamC | = | Bacillomycin D |
| 2,4-DAPG | = | 2, 4-diacetyphloroglucinol |
References
- Slaton, N.A.; Cartwright, R.D.; Meng, J.; Gbur, E.E.; Norman, R.J. Sheath blight severity and rice yield as affected by nitrogen fertilizer rate, application method, and fungicide. Agron. J. 2003, 95, 1489–1496. [Google Scholar] [CrossRef]
- Ben Hassen, M.; Monaco, F.; Facchi, A.; Romani, M.; Valè, G.; Sali, G. Economic Performance of Traditional and Modern Rice Varieties under Different Water Management Systems. Sustainability 2017, 9, 347. [Google Scholar] [CrossRef] [Green Version]
- Rejeb, K.B.; Abdelly, C.; Savouré, A. How reactive oxygen species and proline face stress together. Plant Physiol. Biochem. 2014, 80, 278–284. [Google Scholar] [CrossRef]
- Sang, M.K.; Kim, K.D. Biocontrol activity and primed systemic resistance by compost water extracts against anthracnoses of pepper and cucumber. Phytopathology 2011, 101, 732–740. [Google Scholar] [CrossRef]
- Sadeghi, A.; Hessan, A.; Askari, H.; Aghighi, S.; Shahidi Bonjar, G. Biological control potential of two Streptomyces isolates on Rhizoctonia solani, the causal agent of damping-off of sugar beet. Pak. J. Biol. Sci. 2006, 9, 904–910. [Google Scholar]
- Kim, S.-Y.; Park, D.-S.; Lee, I.-J.; Park, Y.-G.; Seo, J.-H.; Bae, H.-K.; Hwang, C.-D.; Ko, J.-M. Disease incidence, growth, and physiological characterization of rice cultivars with different susceptibility to bakanae disease. Phili. J. Crop. Sci. PJCS 2018, 43, 38–46. [Google Scholar]
- Chung, E.J.; Hossain, M.T.; Khan, A.; Kim, K.H.; Jeon, C.O.; Chung, Y.R. Bacillus oryzicola sp. nov., an endophytic bacterium isolated from the roots of rice with antimicrobial, plant growth promoting, and systemic resistance inducing activities in rice. Plant. Pathol. J. 2015, 31, 152. [Google Scholar] [CrossRef]
- Goswami, R.S.; Kistler, H.C. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef]
- Park, J.W.; Choi, S.-Y.; Hwang, H.-J.; Kim, Y.-B. Fungal mycoflora and mycotoxins in Korean polished rice destined for humans. Int. J. Food Microbiol. 2005, 103, 305–314. [Google Scholar] [CrossRef]
- Mew, T.W.; Cottyn, B.; Pamplona, R.; Barrios, H.; Xiangmin, L.; Zhiyi, C.; Fan, L.; Nil-panit, N.; Arunyanart, P.; Van Kim, P.; et al. Applying Rice Seed-Associated Antagonistic Bacteria to Manage Rice Sheath Blight in Developing Countries. Plant Dis. 2004, 88, 557–564. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.-H.; Ndoye, M.; Zhang, J.-B.; Li, H.-P.; Liao, Y.-C. Population structure and genetic diversity of the Fusarium graminearum species complex. Toxins 2011, 3, 1020–1037. [Google Scholar] [CrossRef]
- Mosquera, G.; Giraldo, M.C.; Khang, C.H.; Coughlan, S.; Valent, B. Interaction transcriptome analysis identifies Magnaporthe oryzae BAS1-4 as biotrophy-associated secreted proteins in rice blast disease. Plant Cell. 2009, 21, 1273–1290. [Google Scholar] [CrossRef] [Green Version]
- Mokiou, S.; Magan, N. Physiological manipulation and formulation of the biocontrol yeast Pichia anomala for control of Penicillium verrucosum and ochratoxin a contamination of moist grain. Biocontrol. Sci. Technol. 2008, 18, 1063–1073. [Google Scholar] [CrossRef] [Green Version]
- Chenniappan, C.; Narayanasamy, M.; Daniel, G.; Ramaraj, G.; Ponnusamy, P.; Sekar, J.; Ramalingam, P.V. Biocontrol efficiency of native plant growth promoting rhizobacteria against rhizome rot disease of turmeric. Biol. Control. 2019, 129, 55–64. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Camele, I.; Racioppi, R.; Scrano, L.; Lacobellis, N.S.; Bufo, S.A. In vitro antifungal activity of Burkholderia gladioli pv. Agaricicola against some phytopathogenic fungi. Int. J. Mol. Sci. 2012, 13, 16291–16302. [Google Scholar] [CrossRef] [Green Version]
- Spence, C.; Alff, E.; Johnson, C.; Ramos, C.; Donofrio, N.; Sundaresan, V.; Bais, H. Natural rice rhizospheric microbes suppress rice blast infections. BMC Plant Biol. 2014, 14, 130. [Google Scholar] [CrossRef] [Green Version]
- Spence, C.A.; Raman, V.; Donofrio, N.M.; Bais, H.P. Global gene expression in rice blast pathogen Magnaporthe oryzae treated with a natural rice soil isolate. Planta 2014, 239, 171–185. [Google Scholar] [CrossRef]
- Spence, C.A.; Lakshmanan, V.; Donofrio, N.; Bais, H.P. Crucial roles of abscisic acid biogenesis in virulence of rice blast fungus Magnaporthe oryzae. Front. Plant Sci. 2015, 6, 1082. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Xie, G.; Li, B.; Luo, Y.; Zhao, L.; Wang, X.; Liu, B.; Li, W. Gram-positive bacteria associated with rice in China and their antagonists against the pathogens of sheath blight and bakanae disease in rice. Rice Sci. 2005, 12, 213–218. [Google Scholar]
- Kazempour, M.N. Biological control of Rhizoctonia solani, the causal agent of rice sheath blight by antagonistics bacteria in greenhouse and field conditions. Plant Pathol. J. 2004, 3, 88–96. [Google Scholar]
- Mehnaz, S.; Mirza, M.S.; Haurat, J.; Bally, R.; Normand, P.; Bano, A.; Malik, K.A. Isolation and 16S rRNA sequence analysis of the beneficial bacteria from the rhizosphere of rice. Can. J. Microbiol. 2001, 47, 110–117. [Google Scholar] [CrossRef]
- Engelhard, M.; Hurek, T.; Reinhold-Hurek, B. Preferential occurrence of diazotrophic endophytes, Azoarcus spp., in wild rice species and land races of (Oryza sativa L.) in comparison with modern races. Environ. Microbiol. 2000, 2, 131–141. [Google Scholar] [CrossRef]
- Yasmin, S.; Rahman Bakar, M.A.; Malik, K.A.; Hafeez, F.Y. Isolation, characterization and beneficial effects of rice associated plant growth promoting bacteria from Zanzibar soils. J. Basic Microbiol. 2004, 44, 241–252. [Google Scholar] [CrossRef]
- Kushwaha, P.; Kashyap, P.L.; Srivastava, A.K.; Tiwari, R.K. Plant growth promoting and antifungal activity in endophytic Bacillus strains from pearl millet (Pennisetum glaucum). Braz. J. Microbiol. 2019. [Google Scholar] [CrossRef]
- Xu, W.; Wang, F.; Zhang, M.; Ou, T.; Wang, R.; Strobel, G.; Xiang, Z.; Zhou, Z.; Xie, J. Diversity of cultivable endophytic bacteria in mulberry and their potential for antimicrobial and plant growth-promoting activities. Microbiol. Res. 2019, 229, 126328. [Google Scholar] [CrossRef]
- Thomas, P.; Sekhar, A.C. Live cell imaging reveals extensive intracellular cytoplasmic colonization of banana by normally non-cultivable endophytic bacteria. AoB Plants 2014, 6. [Google Scholar] [CrossRef] [Green Version]
- Warrior, P. Living systems as natural crop-protection agents. Pest Manag. Sci. 2000, 56, 681–687. [Google Scholar] [CrossRef]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [Green Version]
- Bloemberg, G.V.; Lugtenberg, B.J. Molecular basis of plant growth promotion and biocontrol by rhizobacteria. Curr. Opin. Plant Biol. 2001, 4, 343–350. [Google Scholar] [CrossRef]
- Adhikari, T.B.; Joseph, C.; Yang, G.; Phillips, D.A.; Nelson, L.M. Evaluation of bacteria isolated from rice for plant growth promotion and biological control of seedling disease of rice. Can. J. Microbiol. 2001, 47, 916–924. [Google Scholar] [CrossRef]
- Berg, G.; Kurze, S.; Buchner, A.; Wellington, E.M.; Smalla, K. Successful strategy for the selection of new strawberry-associated rhizobacteria antagonistic to Verticillium wilt. Can. J. Microbiol. 2000, 46, 1128–1137. [Google Scholar] [CrossRef]
- Passari, A.K.; Mishra, V.K.; Singh, G.; Singh, P.; Kumar, B.; Gupta, V.K.; Sarma, R.K.; Saikia, R.; Singh, B.P. Insights into the functionality of endophytic actinobacteria with a focus on their biosynthetic potential and secondary metabolites production. Sci. Rep. 2017, 7, 11809. [Google Scholar] [CrossRef]
- Rosenblueth, M.; Martínez-Romero, E. Bacterial endophytes and their interactions with hosts. Mol. Plant Microbe Interact. 2006, 19, 827–837. [Google Scholar] [CrossRef] [Green Version]
- Ryan, R.P.; Germaine, K.; Franks, A.; Ryan, D.J.; Dowling, D.N. Bacterial endophytes: Recent developments and applications. FEMS Microbiol. Lett. 2008, 278, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Pageni, B.B.; Lupwayi, N.Z.; Larney, F.J.; Kawchuk, L.M.; Gan, Y. Populations, diversity and identities of bacterial endophytes in potato (Solanum tuberosum L.) cropping systems. Can. J. Plant Sci. 2013, 93, 1125–1142. [Google Scholar] [CrossRef] [Green Version]
- Verma, P.; Yadav, A.N.; Kazy, S.K.; Saxena, A.K.; Suman, A. Evaluating the diversity and phylogeny of plant growth promoting bacteria associated with wheat (Triticum aestivum L.) growing in central zone of India. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 432–447. [Google Scholar]
- Mercado-Blanco, J. Life of Microbes inside the Plant. In Principles of Plant-Microbe Interactions: Microbes for Sustainable Agriculture; Lugtenberg, B., Ed.; Springer: Cham, Switzerland, 2015; pp. 25–32. [Google Scholar]
- Verma, P.; Yadav, A.N.; Khannam, K.S.; Panjiar, N.; Kumar, S.; Saxena, A.K.; Suman, A. Assessment of genetic diversity and plant growth promoting attributes of psychrotolerant bacteria allied with wheat (Triticum aestivum L.) from the northern hills zone of India. Ann. Microbiol. 2015, 65, 1885–1899. [Google Scholar] [CrossRef]
- Long, H.H.; Schmidt, D.D.; Baldwin, I.T. Native Bacterial Endophytes Promote Host Growth in a Species-Specific Manner; Phytohormones Manipulations Do Not Result in Common Growth Responses. PLoS ONE 2008, 3, e2702. [Google Scholar] [CrossRef] [Green Version]
- Aravind, R.; Kumar, A.; Eapen, S. Pre-plant bacterization: A strategy for delivery of beneficial endophytic bacteria and production of disease-free plantlets of black pepper (Piper nigrum L.). Arch. Phytopathol. Plant Protect. 2012, 45, 1115–1126. [Google Scholar] [CrossRef]
- Bhore, S.J.; Sathisha, G. Screening of endophytic colonizing bacteria for cytokinin-like compounds: Crude cell-free broth of endophytic colonizing bacteria is unsuitable in cucumber cotyledon bioassay. World J. Agric. Sci. 2010, 6, 345–352. [Google Scholar]
- Islam, A.; Kabir, M.S. Molecular Identification and Evaluation of Indigenous Bacterial Isolates for Their Plant Growth Promoting and Biological Control Activities against Fusarium Wilt Pathogen of Tomato. Plant Pathol. J. 2019, 35, 137–148. [Google Scholar] [CrossRef]
- Yasmin, S.; Zaka, A.; Imran, A.; Zahid, M.A.; Yousaf, S.; Rasul, G.; Arif, M.; Mirza, M.S. Plant growth promotion and suppression of bacterial leaf blight in rice by inoculated bacteria. PLoS ONE 2016, 11, e0160688. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.-G.; Weon, H.-Y.; Lee, S.-Y. In vitro antagonistic effects of Bacilli isolates against four soilborne plant pathogenic fungi. Plant Pathol. J. 2008, 24, 52–57. [Google Scholar] [CrossRef] [Green Version]
- Patil, H.J.; Srivastava, A.K.; Kumar, S.; Chaudhari, B.L.; Arora, D.K. Selective isolation, evaluation and characterization of antagonistic actinomycetes against Rhizoctonia solani. World J. Microbiol. Biotechnol. 2010, 26, 2163–2170. [Google Scholar] [CrossRef]
- Naureen, Z.; Price, A.H.; Hafeez, F.Y.; Roberts, M.R. Identification of rice blast disease-suppressing bacterial strains from the rhizosphere of rice grown in Pakistan. Crop. Protect. 2009, 28, 1052–1060. [Google Scholar] [CrossRef] [Green Version]
- Cazorla, F.; Romero, D.; Pérez-García, A.; Lugtenberg, B.; Vicente, A.D.; Bloemberg, G. Isolation and characterization of antagonistic Bacillus subtilis strains from the avocado rhizoplane displaying biocontrol activity. J. Appl. Microbiol. 2007, 103, 1950–1959. [Google Scholar] [CrossRef]
- Ghazanfar, M.; Waqas, W.; Sahi, S. Influence of various fungicides on the management of rice blast disease. Mycopath 2009, 7, 29–34. [Google Scholar]
- Miah, G.; Rafii, M.; Ismail, M.; Sahebi, M.; Hashemi, F.; Yusuff, O.; Usman, M. Blast disease intimidation towards rice cultivation: A review of pathogen and strategies to control. J. Anim. Plant Sci. 2017, 27, 1058–1066. [Google Scholar]
- Desai, S.; Reddy, M.S.; Kloepper, J.W. Comprehensive testing of biocontrol agents. Biol. Control Crop. Dis. 2002, 3, 387–420. [Google Scholar]
- Palazzini, J.M.; Yerkovich, N.; Alberione, E.; Chiotta, M.; Chulze, S.N. An integrated dual strategy to control Fusarium graminearum sensu stricto by the biocontrol agent Streptomyces sp. RC 87B under field conditions. Plant Gene 2017, 9, 13–18. [Google Scholar] [CrossRef]
- Wang, L.-Y.; Xie, Y.-S.; Cui, Y.-Y.; Xu, J.; He, W.; Chen, H.-G.; Guo, J.-H. Conjunctively screening of biocontrol agents (BCAs) against fusarium root rot and fusarium head blight caused by Fusarium graminearum. Microbiol. Res. 2015, 177, 34–42. [Google Scholar] [CrossRef]
- Xue, A.G.; Chen, Y.; Voldeng, H.D.; Fedak, G.; Savard, M.E.; Längle, T.; Zhang, J.; Harman, G.E. Concentration and cultivar effects on efficacy of CLO-1 biofungicide in controlling Fusarium head blight of wheat. Biol. Control 2014, 73, 2–7. [Google Scholar] [CrossRef]
- Zhang, S.; Schisler, D.A.; Boehm, M.J.; Slininger, P.J. Utilization of chemical inducers of resistance and Cryptococcus flavescens OH 182.9 to reduce Fusarium head blight under greenhouse conditions. Biol. Control 2007, 42, 308–315. [Google Scholar] [CrossRef]
- Dunlap, C.A.; Bowman, M.J.; Schisler, D.A. Genomic analysis and secondary metabolite production in Bacillus amyloliquefaciens AS 43.3: A biocontrol antagonist of Fusarium head blight. Biol. Control 2013, 64, 166–175. [Google Scholar] [CrossRef]
- Zhao, Y.; Selvaraj, J.N.; Xing, F.; Zhou, L.; Wang, Y.; Song, H.; Tan, X.; Sun, L.; Sangare, L.; Folly, Y.M.E. Antagonistic action of Bacillus subtilis strain SG6 on Fusarium graminearum. PLoS ONE 2014, 9, e92486. [Google Scholar] [CrossRef]
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307. [Google Scholar] [CrossRef]
- Shi, C.; Yan, P.; Li, J.; Wu, H.; Li, Q.; Guan, S. Biocontrol of Fusarium graminearum growth and deoxynivalenol production in wheat kernels with bacterial antagonists. Int. J. Env. Res. Public Health 2014, 11, 1094–1105. [Google Scholar] [CrossRef] [Green Version]
- Pal, K.; Tilak, K.; Saxcna, A.; Dey, R.; Singh, C. Suppression of maize root diseases caused by Macrophomina phaseolina, Fusarium moniliforme and Fusarium graminearum by plant growth promoting rhizobacteria. Microbiol. Res. 2001, 156, 209–223. [Google Scholar] [CrossRef]
- Miyake, N.; Chilton, J.; Psatha, M.; Cheng, L.; Andrews, C.; Chan, W.-M.; Law, K.; Crosier, M.; Lindsay, S.; Cheung, M. Human CHN1 mutations hyperactivate α2-chimaerin and cause Duane’s retraction syndrome. Science 2008, 321, 839–843. [Google Scholar] [CrossRef] [Green Version]
- Tateishi, U.; Yamaguchi, U.; Seki, K.; Terauchi, T.; Arai, Y.; Hasegawa, T. Glut-1 expression and enhanced glucose metabolism are associated with tumour grade in bone and soft tissue sarcomas: A prospective evaluation by [18 F] fluorodeoxyglucose positron emission tomography. Eur. J. Nucl. Med. Mol. Imag. 2006, 33, 683. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; De Bruijn, I.; Nybroe, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef] [Green Version]
- Xi, K.; Stephens, J.; Verma, P. Application of formulated rhizobacteria against root rot of field pea. Plant Pathol. 1996, 45, 1150–1158. [Google Scholar] [CrossRef]
- Ji, S.H.; Gururani, M.A.; Chun, S.-C. Isolation and characterization of plant growth promoting endophytic diazotrophic bacteria from Korean rice cultivars. Microbiol. Res. 2014, 169, 83–98. [Google Scholar] [CrossRef]
- Smitha, M.; Singh, R. Biocontrol of phytopathogenic fungi using mycolytic enzymes produced by rhizospheric bacteria of Cicer arietinum. Indian J. Agric. Biochem. 2014, 27, 215–218. [Google Scholar]
- Ahmad, Z.; Wu, J.; Chen, L.; Dong, W. Isolated Bacillus subtilis strain 330-2 and its antagonistic genes identified by the removing PCR. Sci. Rep. 2017, 7, 1777. [Google Scholar] [CrossRef] [Green Version]
- Bach, E.; dos Santos Seger, G.D.; de Carvalho Fernandes, G.; Lisboa, B.B.; Passaglia, L.M.P. Evaluation of biological control and rhizosphere competence of plant growth promoting bacteria. Appl. Soil Ecol. 2016, 99, 141–149. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, M.; Wang, J.; Lv, D.; Ma, Y.; Zhou, B.; Wang, B. Biocontrol effects of Brevibacillus laterosporus AMCC100017 on potato common scab and its impact on rhizosphere bacterial communities. Biol. Control 2017, 106, 89–98. [Google Scholar] [CrossRef]
- Kumar, P.; Dubey, R.C.; Maheshwari, D.K. Bacillus strains isolated from rhizosphere showed plant growth promoting and antagonistic activity against phytopathogens. Microbiol. Res. 2012, 167, 493–499. [Google Scholar] [CrossRef]
- Bibi, F.; Yasir, M.; Song, G.-C.; Lee, S.-Y.; Chung, Y.-R. Diversity and characterization of endophytic bacteria associated with tidal flat plants and their antagonistic effects on oomycetous plant pathogens. Plant Pathol. J. 2012, 28, 20–31. [Google Scholar] [CrossRef] [Green Version]
- Koh, M.C.; Lim, C.H.; Chua, S.B.; Chew, S.T.; Phang, S.T. Random amplified polymorphic DNA (RAPD) fingerprints for identification of red meat animal species. Meat Sci. 1998, 48, 275–285. [Google Scholar] [CrossRef]
- Versalovic, J.; Koeuth, T.; Lupski, R. Distribution of repetitive DNA sequences in eubacteria and application to fingerprinting of bacterial genomes. Nucleic Acids Res. 1991, 19, 6823–6831. [Google Scholar] [CrossRef]
- Kim, O.-S.; Cho, Y.-J.; Lee, K.; Yoon, S.-H.; Kim, M.; Na, H.; Park, S.-C.; Jeon, Y.S.; Lee, J.-H.; Yi, H. Introducing EzTaxon-e: A prokaryotic 16S rRNA gene sequence database with phylotypes that represent uncultured species. Int. J. Syst. Evol. Microbiol. 2012, 62, 716–721. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Shakeel, M.; Rais, A.; Hassan, M.N.; Hafeez, F.Y. Root associated Bacillus sp. improves growth, yield and zinc translocation for basmati rice (Oryza sativa) varieties. Front. Microbiol. 2015, 6, 1286. [Google Scholar] [CrossRef]

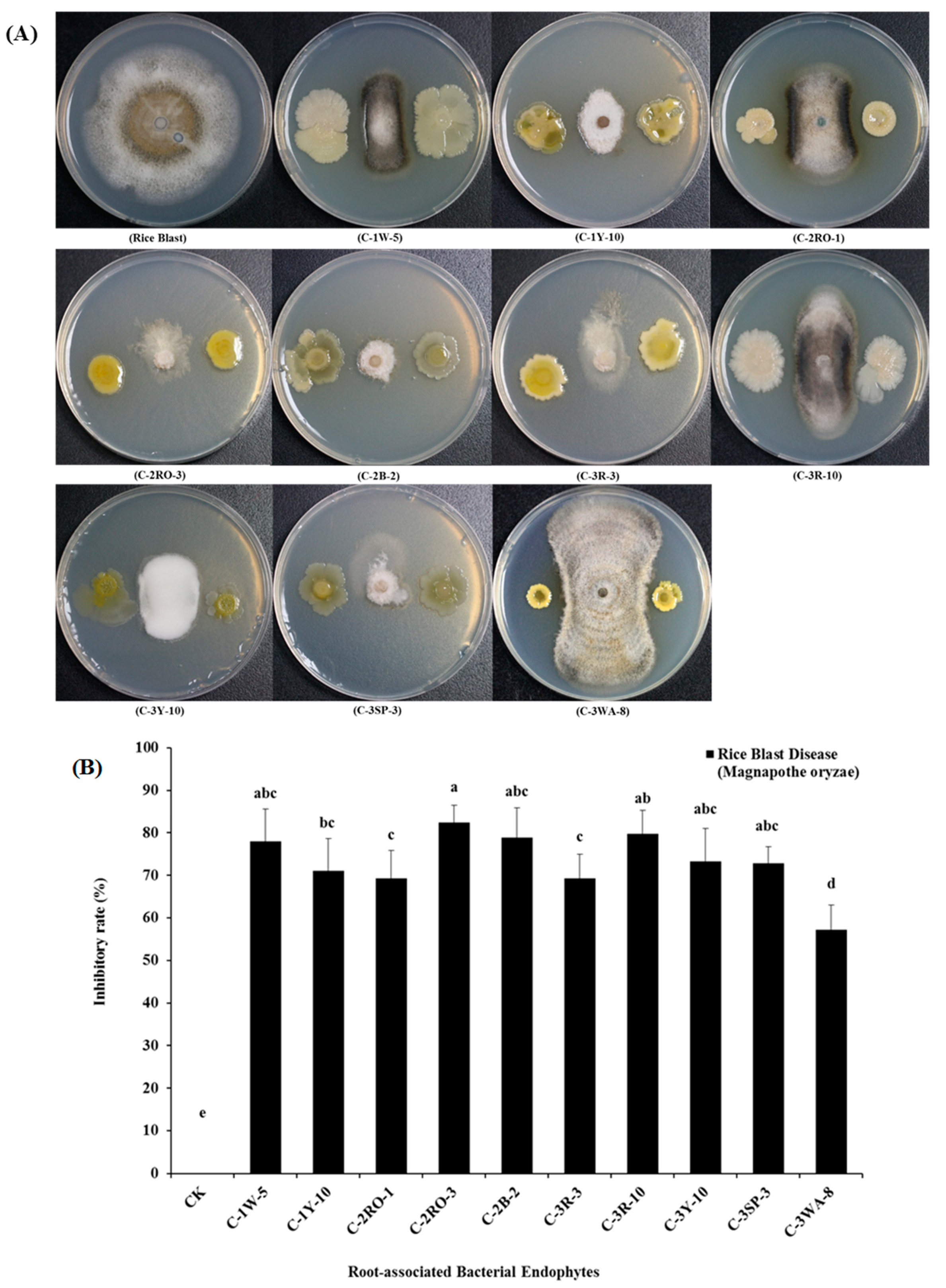


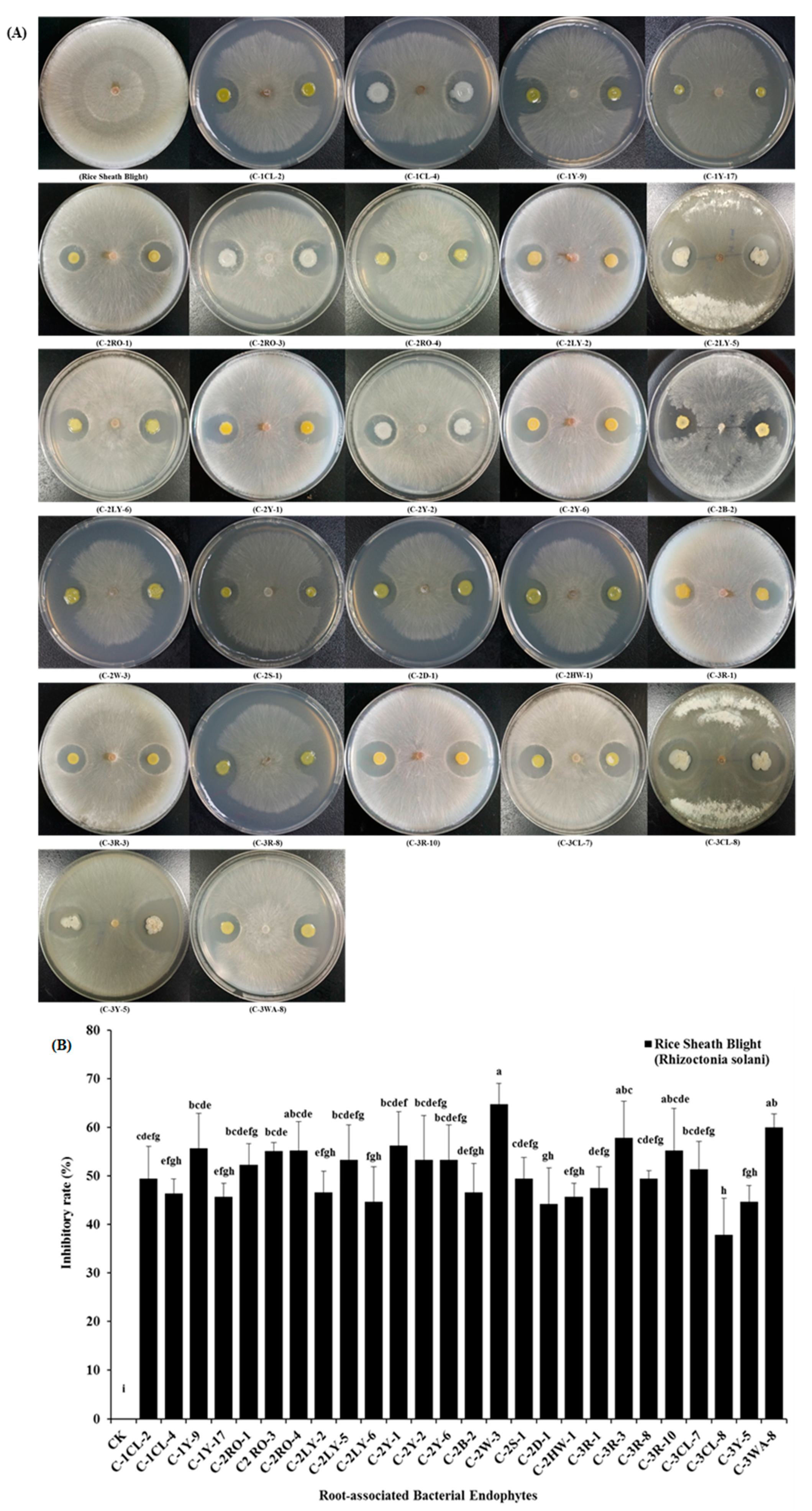

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Associated Cultivars | Endophyte Bacterial ID Code (a) | Amplified 16S rRNA Gene (bps) (b) | NCBI Accession No 16S rRNA Gene Sequences (c) | Closely Related Species, NCBI Accession No 16S rRNA Gene Sequence (d) | Similarity (%) | Completeness (%) |
|---|---|---|---|---|---|---|
| Xiushui-48 | C-1B-2 | 1418 | MN543754 | Bacillus marisflavi JCM11544(T) LGUE01000011 | 99.8 | 96.5 |
| Xiushui-48 | C-1B-3 | 1413 | MN543755 | Bacillus altitudinis 41KF2b(T)ASJC01000029 | 99.7 | 96.3 |
| Xiushui-48 | C-1B-5 | 1412 | MN543756 | Lysinibacillus fusiformis NBRC15717(T) AB271743 | 99.6 | 96.1 |
| Xiushui-48 | C-1B-6 | 1412 | MN543757 | Bacillus cereus ATCC14579(T) AE016877 | 99.9 | 96.1 |
| Xiushui-48 | C-1B-7 | 1412 | MN543758 | Bacillus cereus ATCC14579(T) AE016877 | 100 | 96.4 |
| Xiushui-48 | C-1B-9 | 1392 | MN543759 | Bacillus aryabhattai B8W22(T) EF114313 | 99.9 | 94.6 |
| Xiushui-48 | C-1CL-2 | 1392 | MN543760 | Bacillus wiedmannii FSLW8-0169(T) LOBC01000053 | 100 | 94.6 |
| Xiushui-48 | C-1CL-4 | 1372 | MN543761 | Bacillus wiedmannii FSLW8-0169(T) LOBC01000053 | 100 | 93.1 |
| Xiushui-48 | C-1F-2 | 1396 | MN543762 | Bacillus pseudomycoides DSM12442(T) ACMX01000133 | 99.8 | 95.1 |
| Xiushui-48 | C-1F-3 | 1374 | MN543763 | Bacillus wiedmannii FSLW8-0169(T) LOBC01000053 | 100 | 93.3 |
| Xiushui-48 | C-1G-1 | 1355 | MN543764 | Bacillus thioparans BMP-1(T) DQ371431 | 99.4 | 92.1 |
| Xiushui-48 | C-1G-3 | 1413 | MN543765 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.7 | 96.3 |
| Xiushui-48 | C-1S-1 | 1379 | MN543766 | Bacillus subterraneous DSM13966(T) FR733689 | 99.4 | 93.5 |
| Xiushui-48 | C-1S-2 | 1376 | MN543767 | Bacillus lehensis MLB-2(T) AY793550 | 99.9 | 93.7 |
| Xiushui-48 | C-1S-3 | 1409 | MN543768 | Fictibacillus phosphorivorans Ca7T(T) JX258924 | 99.9 | 96.1 |
| Xiushui-48 | C-1S-4 | 1379 | MN543769 | Bacillus subterraneous DSM13966(T) FR733689 | 99.4 | 93.6 |
| Xiushui-48 | C-1S-5 | 1412 | MN543770 | Bacillus acidiceler CBD119(T) DQ374637 | 99.7 | 96.2 |
| Xiushui-48 | C-1ST-4 | 1375 | MN543771 | Bacillus subterraneous DSM13966(T) FR733689 | 99.3 | 93.5 |
| Xiushui-48 | C-1ST-5 | 1355 | MN543772 | Bacillus thioparans BMP-1(T) DQ371431 | 99.8 | 92.1 |
| Xiushui-48 | C-1W-3 | 1388 | MN543773 | Lysinibacillus fusiformis NBRC15717(T) AB271743 | 99.7 | 94.2 |
| Xiushui-48 | C-1W-5 | 1393 | MN543774 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.9 | 94.7 |
| Xiushui-48 | C-1W-6 | 1393 | MN543775 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.8 | 95.0 |
| Xiushui-48 | C-1W-14 | 1391 | MN543776 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.8 | 94.6 |
| Xiushui-48 | C-1WF-1 | 1382 | MN543777 | Bacillus aryabhattai B8W22(T) EF114313 | 99.8 | 93.9 |
| Xiushui-48 | C-1WF-2 | 1413 | MN543778 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.7 | 96.3 |
| Xiushui-48 | C-1WF-3 | 1403 | MN543779 | Lysinibacillus fusiformis NBRC15717(T) AB271743 | 99.7 | 95.5 |
| Xiushui-48 | C-1WF-7 | 1401 | MN543780 | Bacillus aryabhattai B8W22(T) EF114313 | 100 | 95.3 |
| Xiushui-48 | C-1WF-9 | 1389 | MN543781 | Bacillus aryabhattai B8W22(T) EF114313 | 100 | 94.4 |
| Xiushui-48 | C-1WNF-1 | 1376 | MN543782 | Bacillus aryabhattai B8W22(T) EF114313 | 100 | 93.5 |
| Xiushui-48 | C-1WNF-2 | 1392 | MN543783 | Bacillus aryabhattai B8W22(T) EF114313 | 99.9 | 94.6 |
| Xiushui-48 | C-1WNF-7 | 1412 | MN543784 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.7 | 96.3 |
| Xiushui-48 | C-1WNF-8 | 1422 | MN543785 | Bacillus aryabhattai B8W22(T) EF114313 | 99.5 | 96.9 |
| Xiushui-48 | C-1WNF-9 | 1408 | MN543786 | Bacillus aryabhattai B8W22(T) EF114313 | 99.9 | 95.7 |
| Xiushui-48 | C-1Y-1 | 1407 | MN543787 | Fictibacillus phosphorivorans Ca7T(T) JX258924 | 99.9 | 95.9 |
| Xiushui-48 | C-1Y-2 | 1412 | MN543788 | Fictibacillus nanhaiensis JSM082006(T) GU477780 | 99.8 | 96.3 |
| Xiushui-48 | C-1Y-4 | 1388 | MN543789 | Bacillus marisflavi JCM11544(T) LGUE01000011 | 99.9 | 94.2 |
| Xiushui-48 | C-1Y-7 | 1422 | MN543790 | Fictibacillus nanhaiensis JSM082006(T) GU477780 | 99.8 | 96.3 |
| Xiushui-48 | C-1Y-8 | 1413 | MN543791 | Fictibacillus phosphorivorans Ca7T(T) JX258924 | 99.8 | 96.3 |
| Xiushui-48 | C-1Y-9 | 1371 | MN543792 | Fictibacillus phosphorivorans Ca7T(T) JX258924 | 99.9 | 93.3 |
| Xiushui-48 | C-1Y-10 | 1391 | MN543793 | Fictibacillus phosphorivorans Ca7T(T) JX258924 | 100 | 94.6 |
| Xiushui-48 | C-1Y-11 | 1418 | MN543794 | Fictibacillus nanhaiensis JSM082006(T) GU477780 | 99.9 | 96.6 |
| Xiushui-48 | C-1Y-12 | 1412 | MN543795 | Fictibacillus nanhaiensis JSM082006(T) GU477780 | 100 | 95.8 |
| Xiushui-48 | C-1Y-13 | 1390 | MN543796 | Fictibacillus phosphorivorans Ca7T(T) JX258924 | 100 | 94.6 |
| Xiushui-48 | C-1Y-16 | 1413 | MN543797 | Fictibacillus phosphorivorans Ca7T(T) JX258924 | 99.8 | 96.3 |
| Xiushui-48 | C-1Y-17 | 1404 | MN543798 | Fictibacillus phosphorivorans Ca7T(T) JX258924 | 99.9 | 95.7 |
| Y-003 | C-2B-1 | 1424 | MN543799 | Paenibacillus alvei NBRC3343(T) NR113577.1 | 99.2 | 99.0 |
| Y-003 | C-2B-2 | 1407 | MN543800 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 100 | 95.9 |
| Y-003 | C-2CL-1 | 1390 | MN543801 | Bacillus aryabhattai B8W22(T) EF114313 | 99.9 | 94.5 |
| Y-003 | C-2D-1 | 1411 | MN543802 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.5 | 96.3 |
| Y-003 | C-2GL-1 | 1428 | MN543803 | Lysinibacillus fusiformis NBRC15717(T) AB271743 | 99.3 | 97.1 |
| Y-003 | C-2GL-3 | 1425 | MN543804 | Lysinibacillus fusiformis NBRC15717(T) AB271743 | 99.7 | 96.7 |
| Y-003 | C-2HW-1 | 1390 | MN543805 | Bacillus aryabhattai B8W22(T) EF114313 | 99.9 | 100 |
| Y-003 | C-2HW-2 | 1390 | MN543806 | Bacillus cereus ATCC 14579(T) AE016877 | 99.8 | 94.5 |
| Y-003 | C-2HW-3 | 1430 | MN543807 | Lysinibacillus mangiferihumi M-GX18(T) JF731238 | 99.9 | 97.1 |
| Y-003 | C-2HW-4 | 1372 | MN543808 | Bacillus aryabhattai B8W22(T) EF114313 | 100 | 93.2 |
| Y-003 | C-2HW-5 | 1328 | MN543809 | Bacillus aryabhattai B8W22(T) EF114313 | 99.9 | 94.0 |
| Y-003 | C-2HW-6 | 1425 | MN543810 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.7 | 96.9 |
| Y-003 | C-2HW-7 | 1429 | MN543811 | Paenibacillus alvei NBRC3343(T) NR113577.1 | 99.2 | 99.0 |
| Y-003 | C-2LY-1 | 1404 | MN543812 | Bacillus aryabhattai B8W22(T) EF114313 | 100 | 95.5 |
| Y-003 | C-2LY-2 | 1394 | MN543813 | Bacillus marisflavi JCM11544(T) LGUE01000011 | 100 | 94.8 |
| Y-003 | C-2LY-4 | 1380 | MN543814 | Lysinibacillus fusiformis NBRC15717(T) AB271743 | 99.7 | 93.7 |
| Y-003 | C-2LY-5 | 1379 | MN543815 | Bacillus marisflavi JCM11544(T) LGUE01000011 | 99.9 | 93.7 |
| Y-003 | C-2LY-6 | 1388 | MN543816 | Bacillus aryabhattai B8W22(T) EF114313 | 100 | 94.4 |
| Y-003 | C-2LY-7 | 1372 | MN543817 | Bacillus aryabhattai B8W22(T) EF114313 | 100 | 93.2 |
| Y-003 | C-2R-1 | 1406 | MN543818 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 100 | 95.9 |
| Y-003 | C-2RO-1 | 1417 | MN543819 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 100 | 96.5 |
| Y-003 | C-2RO-3 | 1403 | MN543820 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 100 | 95.4 |
| Y-003 | C-2RO-4 | 1416 | MN543821 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.6 | 96.7 |
| Y-003 | C-2S-1 | 1391 | MN543822 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.7 | 94.8 |
| Y-003 | C-2SN-1 | 1393 | MN543823 | Bacillus cereus ATCC14579(T) AE016877 | 100 | 94.7 |
| Y-003 | C-2SN-2 | 1410 | MN543824 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.7 | 96.0 |
| Y-003 | C-2SN-3 | 1410 | MN543825 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.6 | 96.1 |
| Y-003 | C-2SP-1 | 1410 | MN543826 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.6 | 96.3 |
| Y-003 | C-2T-1 | 1375 | MN543827 | Paenibacillus cucumis AP-115(T) KU201962 | 99.5 | 93.0 |
| Y-003 | C-2W-1 | 1406 | MN543828 | Bacillus cereus ATCC14579(T) AE016877 | 100 | 95.4 |
| Y-003 | C-2W-2 | 1374 | MN543829 | Bacillus cereus ATCC14579(T) AE016877 | 100 | 93.3 |
| Y-003 | C-2W-3 | 1426 | MN543830 | Bacillus tequilensis KCTC13622(T) AYTO01000043 | 99.7 | 96.9 |
| Y-003 | C-2W-4 | 1421 | MN543831 | Bacillus cereus ATCC14579(T) AE016877 | 100 | 96.7 |
| Y-003 | C-2W-5 | 1420 | MN543832 | Bacillus cereus ATCC14579(T) AE016877 | 100 | 100 |
| Y-003 | C-2Y-1 | 1379 | MN543833 | Bacillus indicus LMG22858(T) JGVU01000003 | 99.9 | 94.7 |
| Y-003 | C-2Y-2 | 1403 | MN543834 | Bacillus marisflavi JCM11544(T) LGUE01000011 | 100 | 95.5 |
| Y-003 | C-2Y-3 | 1424 | MN543835 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.8 | 96.9 |
| Y-003 | C-2Y-4 | 1411 | MN543836 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.8 | 96.3 |
| Y-003 | C-2Y-5 | 1429 | MN543837 | Bacillus cereus ATCC14579(T) AE016877 | 99.7 | 97.0 |
| Y-003 | C-2Y-6 | 1394 | MN543838 | Bacillus marisflavi JCM11544(T) LGUE01000011 | 100 | 94.8 |
| CO-39 | C-3CL-1 | 1402 | MN543839 | Bacillus wiedmannii FSLW80169(T) LOBC01000053 | 100 | 95.3 |
| CO-39 | C-3CL-3 | 1394 | MN543840 | Bacillus wiedmannii FSLW80169(T) LOBC01000053 | 100 | 94.8 |
| CO-39 | C-3CL-4 | 1383 | MN543841 | Bacillus wiedmannii FSLW80169(T) LOBC01000053 | 100 | 93.9 |
| CO-39 | C-3CL-5 | 1445 | MN543842 | Bacillus cereus ATCC14579(T) AE016877 | 99.7 | 98.4 |
| CO-39 | C-3CL-6 | 1430 | MN543843 | Bacillus cereus ATCC14579(T) AE016877 | 100 | 97.1 |
| CO-39 | C-3CL-7 | 1445 | MN543844 | Bacillus tequilensis KCTC13622(T) AYTO01000043 | 99.7 | 96.9 |
| CO-39 | C-3CL-8 | 1418 | MN543845 | Bacillus wiedmannii FSLW80169(T) LOBC01000053 | 99.9 | 96.7 |
| CO-39 | C-3F-5 | 1438 | MN543846 | Lysinibacillus mangiferihumi MGX18(T) JF731238 | 99.2 | 97.6 |
| CO-39 | C-3F-7 | 1429 | MN543847 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.7 | 97.1 |
| CO-39 | C-3F-8 | 1439 | MN543848 | Lysinibacillus fusiformis NBRC15717(T) AB271743 | 99.6 | 97.9 |
| CO-39 | C-3G-1 | 1423 | MN543849 | Bacillus wiedmannii FSLW80169(T) LOBC01000053 | 99.7 | 96.9 |
| CO-39 | C-3P-3 | 1380 | MN543850 | Cupriavidus metallidurans CH34(T) CP000353 | 99.6 | 95.2 |
| CO-39 | C-3P-5 | 1408 | MN543851 | Cupriavidus metallidurans CH34(T) CP000353 | 99.2 | 97.2 |
| CO-39 | C-3R-1 | 1416 | MN543852 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 100 | 96.3 |
| CO-39 | C-3R-2 | 1428 | MN543853 | Bacillus aryabhattai B8W22(T) EF114313 | 99.3 | 96.9 |
| CO-39 | C-3R-3 | 1427 | MN543854 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.9 | 97.1 |
| CO-39 | C-3R-5 | 1407 | MN543855 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.7 | 96.0 |
| CO-39 | C-3R-6 | 1414 | MN543856 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.7 | 96.5 |
| CO-39 | C-3R-7 | 1418 | MN543857 | Lysinibacillus fusiformis NBRC15717(T) AB271743 | 99.5 | 96.5 |
| CO-39 | C-3R-8 | 1437 | MN543858 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.9 | 98.0 |
| CO-39 | C-3R-9 | 1382 | MN543859 | Bacillus cereus ATCC14579(T) AE016877 | 100 | 93.8 |
| CO-39 | C-3R-10 | 1425 | MN543860 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 100 | 96.9 |
| CO-39 | C-3SP-3 | 1399 | MN543861 | Cupriavidus metallidurans CH34(T) CP000353 | 100 | 93.3 |
| CO-39 | C-3T-1 | 1355 | MN543862 | Cupriavidus metallidurans CH34(T) CP000353 | 100 | 93.3 |
| CO-39 | C-3T-2 | 1380 | MN543863 | Cupriavidus metallidurans CH34(T) CP000353 | 99.6 | 95.2 |
| CO-39 | C-3T-3 | 1398 | MN543864 | Cupriavidus metallidurans CH34(T) CP000353 | 99.6 | 96.4 |
| CO-39 | C-3T-4 | 1413 | MN543865 | Paenibacillus uliginis N3/975(T) NR117012.1 | 96.6 | 99.0 |
| CO-39 | C-3T-5 | 1364 | MN543866 | Cupriavidus metallidurans CH34(T) CP000353 | 99.2 | 97.3 |
| CO-39 | C-3T-6 | 1407 | MN543867 | Cupriavidus metallidurans CH34(T) CP000353 | 99.5 | 94.0 |
| CO-39 | C-3T-7 | 1399 | MN543868 | Paenibacillus cucumis AP-115(T) KU201962 | 99.6 | 94.9 |
| CO-39 | C-3T-8 | 1399 | MN543869 | Cupriavidus metallidurans CH34(T) CP000353 | 99.5 | 96.6 |
| CO-39 | C-3T-9 | 1355 | MN543870 | Microbacterium laevaniformans DSM20140(T) Y17234 | 99.7 | 93.8 |
| CO-39 | C-3W-3 | 1418 | MN543871 | Lysinibacillus fusiformis NBRC15717(T) AB271743 | 99.4 | 96.6 |
| CO-39 | C-3WA-8 | 1395 | MN543872 | Bacillus altitudinis 41KF2b(T) ASJC01000029 | 99.8 | 95.0 |
| CO-39 | C-3Y-2 | 1415 | MN543873 | Bacillus marisflavi JCM11544(T) LGUE01000011 | 99.9 | 96.5 |
| CO-39 | C-3Y-5 | 1416 | MN543874 | Bacillus marisflavi JCM11544(T) LGUE01000011 | 100 | 96.6 |
| CO-39 | C-3Y-10 | 1390 | MN543875 | Fictibacillus phosphorivorans Ca7(T) JX258924 | 100 | 94.6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khaskheli, M.A.; Wu, L.; Chen, G.; Chen, L.; Hussain, S.; Song, D.; Liu, S.; Feng, G. Isolation and Characterization of Root-Associated Bacterial Endophytes and Their Biocontrol Potential against Major Fungal Phytopathogens of Rice (Oryza sativa L.). Pathogens 2020, 9, 172. https://doi.org/10.3390/pathogens9030172
Khaskheli MA, Wu L, Chen G, Chen L, Hussain S, Song D, Liu S, Feng G. Isolation and Characterization of Root-Associated Bacterial Endophytes and Their Biocontrol Potential against Major Fungal Phytopathogens of Rice (Oryza sativa L.). Pathogens. 2020; 9(3):172. https://doi.org/10.3390/pathogens9030172
Chicago/Turabian StyleKhaskheli, Maqsood Ahmed, Lijuan Wu, Guoqing Chen, Long Chen, Sajid Hussain, Dawei Song, Sihui Liu, and Guozhong Feng. 2020. "Isolation and Characterization of Root-Associated Bacterial Endophytes and Their Biocontrol Potential against Major Fungal Phytopathogens of Rice (Oryza sativa L.)" Pathogens 9, no. 3: 172. https://doi.org/10.3390/pathogens9030172