Different Cell Wall-Degradation Ability Leads to Tissue-Specificity between Xanthomonas oryzae pv. oryzae and Xanthomonas oryzae pv. oryzicola

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Rice Cultivar
2.2. Pathogen Inoculation and Filter Paper in Vitro Assay
2.3. Normal and Cryo Field-Emission Scanning Electron Microscopy
2.4. Transmission Electron Microscopy
2.5. Cellulose Synthase Gene Expression Analysis
2.6. Statistical Analysis
3. Results
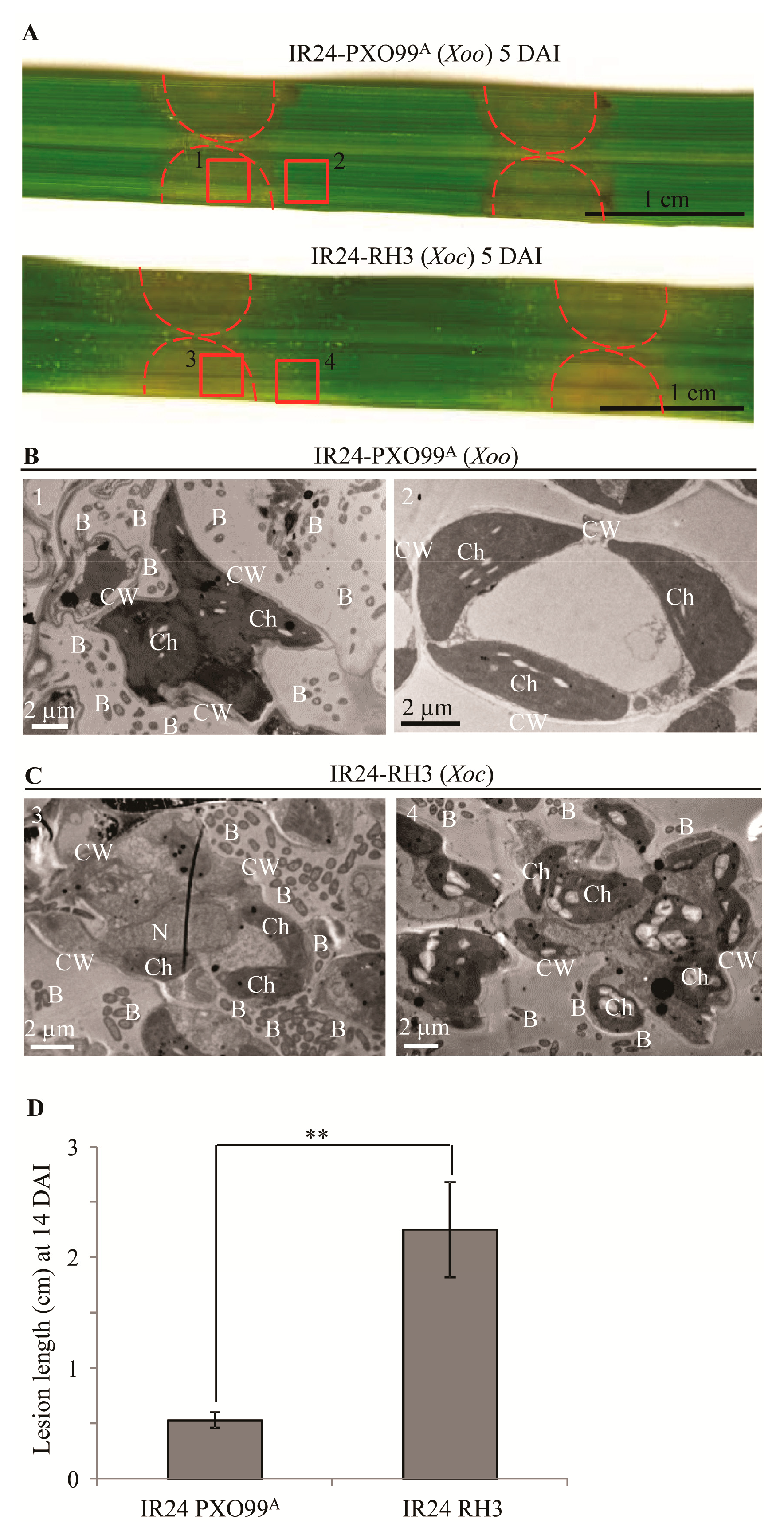
3.1. The Leaf Tissue Morphology of Susceptible Rice Infected with Xoo and Xoc
3.2. The Cell Wall Surface Structures of Mesophyll Cells in Rice Infected with Xoo and Xoc
3.3. The Cell Wall Inner Structure of Mesophyll Cells in Rice Infected with Xoo and Xoc
3.4. The Cellulose Filter Papers Less Damaged by Xoo than Xoc
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Niño-Liu, D.O.; Ronald, P.C.; Bogdanove, A.J. Xanthomonas oryzae pathovars: Model Pathogens of A Model Crop. Mol. Plant Pathol. 2006, 7, 303–324. [Google Scholar] [CrossRef]
- Ke, Y.; Deng, H.; Wang, S. Advances in Understanding Broad-Spectrum Resistance to Pathogens in Rice. Plant J. 2017, 90, 738–748. [Google Scholar] [CrossRef]
- Poulin, L.; Grygiel, P.; Magne, M.; Gagnevin, L.; Rodriguez-R, L.M.; Serna, N.F.; Zhao, S.; El Rafii, M.; Dao, S.; Tekete, C.; et al. New Multilocus Variable-Number Tandem-Repeat Analysis Tool for Surveillance and Local Epidemiology of Bacterial Leaf Blight and Bacterial Leaf Streak of Rice Caused by Xanthomonas oryzae. Appl. Environ. Microbiol. 2015, 81, 688–698. [Google Scholar] [CrossRef] [Green Version]
- Bogdanove, A.J.; Koebnik, R.; Gudlur, A.; Furutani, A.; Angiuoli, S.; Patil, P.B.; Van Sluys, M.-A.; Ryan, R.; Meyer, D.; Han, S.-W.; et al. Two New Complete Genome Sequences Offer Insight into Host and Tissue Specificity of Plant Pathogenic Xanthomonas spp. J. Bacteriol. 2011, 193, 5450–5464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.; Patil, P.; Van Sluys, M.-A.; White, F.F.; Ryan, R.; Dow, J.M.; Rabinowicz, P.; Salzberg, S.L.; Leach, J.E.; Sonti, R.; et al. Acquisition and Evolution of Plant Pathogenesis–Associated Gene Clusters and Candidate Determinants of Tissue-Specificity in Xanthomonas. PLOS ONE 2008, 3, e3828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, C.; Kirchner, O.; Egler, M.; Stuttmann, J.; Bonas, U.; Büttner, D. HpaA from Xanthomonas is A regulator of type III secretion. Mol. Microbiol. 2008, 69, 344–360. [Google Scholar] [CrossRef]
- Lindeberg, M.; Salmond, G.P.C.; Collmer, A. Complementation of Deletion Mutations in A Cloned Functional Cluster of Erwinia chrysanthemi out Genes with Erwinia carotovora out Homologues Reveals OutC and OutD as Candidate Gatekeepers of Species-Specific Secretion of Proteins Via the Type II Pathway. Mol. Microbiol. 1996, 20, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.-S.; Song, X.; Zou, L.-F.; Yuan, L.; Li, Y.-R.; Guo, W.; Che, Y.-Z.; Zhao, W.; Duan, Y.-P.; Chen, G.-Y. EcpA, An Extracellular Protease, is a Specific Virulence Factor Required by Xanthomonas oryzae pv. oryzicola but not by X. oryzae pv. oryzae in Rice. Microbiol. 2012, 158, 2372–2383. [Google Scholar] [PubMed] [Green Version]
- Debroy, S.; Dao, J.; Söderberg, M.; Rossier, O.; Cianciotto, N.P. Legionella Pneumophila Type II Secretome Reveals Unique Exoproteins and a Chitinase that Promotes Bacterial Persistence in the Lung. Proc. Natl. Acad. Sci. USA 2006, 103, 19146–19151. [Google Scholar] [CrossRef] [Green Version]
- Szurek, B.; Marois, E.; Bonas, U.; Ackerveken, G.V.D. Eukaryotic Features of the Xanthomonas Type III Effector AvrBs3: Protein Domains Involved in Transcriptional Activation and the Interaction with Nuclear Import Receptors from Pepper. Plant J. 2001, 26, 523–534. [Google Scholar] [CrossRef]
- Hummel, A.W.; Wilkins, K.E.; Wang, L.; Cernadas, R.A.; Bogdanove, A.J. A Transcription Activator-Like Effector from Xanthomonas oryzae pv. oryzicola Elicits Dose-Dependent Resistance in Rice. Mol. Plant Pathol. 2016, 18, 55–66. [Google Scholar]
- Jacques, M.-A.; Arlat, M.; Boulanger, A.; Boureau, T.; Carrere, S.; Cesbron, S.; Chen, N.; Cociancich, S.; Darrasse, A.; Denancé, N.; et al. Using Ecology, Physiology, and Genomics to Understand Host Specificity in Xanthomonas: French Network on Xanthomonads (FNX). Annu. Rev. Phytopathol. 2016, 54, 163–187. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Sonti, R.V. Role of the FeoB Protein and Siderophore in Promoting Virulence of Xanthomonas oryzae pv. oryzae on Rice. J. Bacteriol. 2010, 192, 3187–3203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, R.; Javvadi, S.G.; Chatterjee, S. Cell-cell Signalling Promotes Ferric Iron Uptake in Xanthomonas oryzae pv. oryzicola that Contribute to Its Virulence and Growth Inside Rice. Mol. Microbiol. 2015, 96, 708–727. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.; Guerinot, M.L. Facing the Challenges of Cu, Fe and Zn Homeostasis in Plants. Nat. Methods 2009, 5, 333–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jha, G.; Rajeshwari, R.; Sonti, R.V. Functional Interplay between Two Xanthomonas oryzae pv. oryzae Secretion Systems in Modulating Virulence on Rice. Mol. Plant-Microbe Interact. 2007, 20, 31–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaad, N.W.; Wang, Z.K.; Di, M.; McBeath, J.; Peterson, G.L.; Bonde, M.R. An Improved Infiltration Technique to test the Pathogenicity of Xanthomonas oryzae pv. oryzae in Rice Seedlings. Seed Sci. Technol. 1996, 24, 449–456. [Google Scholar]
- Ma, H.; Xiao, J.; Wang, S.; Chen, J.; Zhang, Z.; Yang, Z.; Li, X. MAPK Kinase 10.2 Promotes Disease Resistance and Drought Tolerance by Activating Different MAPKs in Rice. Plant J. 2017, 92, 557–570. [Google Scholar] [CrossRef] [Green Version]
- Salzberg, S.L.; Sommer, D.D.; Schatz, M.C.; Phillippy, A.; Rabinowicz, P.D.; Tsuge, S.; Furutani, A.; Ochiai, H.; Delcher, A.L.; Kelley, D.R.; et al. Genome Sequence and Rapid Evolution of the Rice Pathogen Xanthomonas oryzae pv. oryzae PXO99A. BMC Genom. 2008, 9, 204. [Google Scholar] [CrossRef] [Green Version]
- Qiu, D.; Xiao, J.; Ding, X.; Xiong, M.; Cai, M.; Cao, Y.; Li, X.; Xu, C.; Wang, S. OsWRKY13 Mediates Rice Disease Resistance by Regulating Defense-Related Genes in Salicylate- and Jasmonate-Dependent Signaling. Mol. Plant-Microbe Interactions 2007, 20, 492–499. [Google Scholar] [CrossRef] [Green Version]
- Bacete, L.; Mélida, H.; Pattathil, S.; Hahn, M.G.; Molina, A.; Miedes, E. Characterization of Plant Cell Wall Damage-Associated Molecular Patterns Regulating Immune Responses. Advanced Structural Safety Studies 2017, 1578, 13–23. [Google Scholar]
- Kim, S.G.; Kim, K.W.; Park, E.W.; Choi, I. Silicon-Induced Cell Wall Fortification of Rice Leaves: A Possible Cellular Mechanism of Enhanced Host Resistance to Blast. Phytopathol. 2002, 92, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, F.D. Ávila; Benhamou, N.; Datnoff, L.; Jones, J.B.; Bélanger, R.R. Ultrastructural and Cytochemical Aspects of Silicon-Mediated Rice Blast Resistance. Phytopathol. 2003, 93, 535–546. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Zhang, M.; Xiao, J.; Li, X.; Yuan, M.; Wang, S. Dominant and Recessive Major R Genes Lead to Different Types of Host Cell Death during Resistance to Xanthomonas oryzae in Rice. Front. Plant Sci. 2018, 9, 1711. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Zhang, M.; Zhu, M.; He, L.; Xiao, J.; Li, X.; Yuan, M. Autophagy-Like Cell Death Regulates Hydrogen Peroxide and Calcium Ion Distribution in Xa3/Xa26-Mediated Resistance to Xanthomonas oryzae pv. oryzae. Int. J. Mol. Sci. 2019, 21, 194. [Google Scholar] [CrossRef] [Green Version]
- Sage, T.L.; Sage, R.F. The Functional Anatomy of Rice Leaves: Implications for Refixation of Photorespiratory CO2 and Efforts to Engineer C4 Photosynthesis into Rice. Plant Cell Physiol. 2009, 50, 756–772. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Man, Y.; Wen, J.; Guo, Y.; Lin, J. Advances in Imaging Plant Cell Walls. Trends Plant Sci. 2019, 24, 867–878. [Google Scholar] [CrossRef]
- Hu, J.; Qian, W.; He, C. The Xanthomonas oryzae pv. oryzae eglXoB Endoglucanase Gene is Required for Virulence to Rice. FEMS Microbiol. Lett. 2007, 269, 273–279. [Google Scholar] [CrossRef]
- Ray, S.K.; Rajeshwari, R.; Sonti, R.V. Mutants of Xanthomonas oryzae pv. oryzae Deficient in General Secretary Pathway are Virulence Deficient and Unable to Secrete Xylanase. Mol. Plant-Microbe Interact. 2000, 13, 394–401. [Google Scholar] [CrossRef] [Green Version]
- Zou, H.-S.; Yuan, L.; Guo, W.; Li, Y.-R.; Che, Y.-Z.; Zou, L.-F.; Chen, G.-Y. Construction of a Tn5-Tagged Mutant Library of Xanthomonas oryzae pv. oryzicola as An Invaluable Resource for Functional Genomics. Curr. Microbiol. 2010, 62, 908–916. [Google Scholar] [CrossRef]
- Hu, K.; Cao, J.; Zhang, J.; Xia, F.; Ke, Y.; Zhang, H.; Xie, W.; Liu, H.; Cui, Y.; Cao, Y.; et al. Improvement of Multiple Agronomic Traits by A Disease Resistance Gene Via Cell Wall Reinforcement. Nat. Plants 2017, 3, 17009. [Google Scholar] [CrossRef]
- Gu, K.; Yang, B.; Tian, D.; Wu, L.; Wang, D.; Sreekala, C.; Yang, F.; Chu, Z.; Wang, G.-L.; White, F.F.; et al. R gene expression induced by A type-III effector triggers disease resistance in rice. Nat. 2005, 435, 1122–1125. [Google Scholar] [CrossRef]
- Fu, J.; Liu, H.; Li, Y.; Yu, H.; Li, X.; Xiao, J.; Wang, S. Manipulating Broad-Spectrum Disease Resistance by Suppressing Pathogen-Induced Auxin Accumulation in Rice. Plant Physiol. 2010, 155, 589–602. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Zhang, X.; Wu, T.; Yuan, B.; Ding, X.; Yao, F.; Chu, Z. The Polygalacturonase-Inhibiting Protein 4 (OsPGIP4), A Potential Component of the qBlsr5a Locus, Confers Resistance to Bacterial Leaf Streak in Rice. Planta 2016, 243, 1297–1308. [Google Scholar] [CrossRef]
- Wu, T.; Peng, C.; Li, B.; Wu, W.; Kong, L.; Li, F.; Chu, Z.; Liu, F.; Ding, X. OsPGIP1-Mediated Resistance to Bacterial Leaf Streak in Rice is Beyond Responsive to the Polygalacturonase of Xanthomonas oryzae pv. oryzicola. Rice 2019, 12, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Ju, Y.; Tian, H.; Zhang, R.; Zuo, L.; Jin, G.; Xu, Q.; Ding, X.; Li, X.; Chu, Z. Overexpression of OsHSP18.0-CI Enhances Resistance to Bacterial Leaf Streak in Rice. Rice 2017, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.-L.; Xu, M.-R.; Zhao, M.-F.; Xie, X.-W.; Zhu, L.; Fu, B.; Li, Z.-K. Genome-Wide Gene Responses in A Transgenic Rice Line Carrying the Maize Resistance Gene Rxo1 to the Rice Bacterial Streak Pathogen, Xanthomonas oryzae pv. oryzicola. BMC Genom. 2010, 11, 78. [Google Scholar] [CrossRef] [Green Version]
- Liao, Z.-X.; Ni, Z.; Wei, X.-L.; Chen, L.; Li, J.-Y.; Yu, Y.-H.; Jiang, W.; Jiang, B.-L.; He, Y.-Q.; Huang, S. Dual RNA-seq of Xanthomonas oryzae pv. oryzicola Infecting Rice Reveals Novel Insights into Bacterial-Plant Interaction. PLOS ONE 2019, 14, e0215039. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, J.; Chu, C.; Zhang, M.; He, L.; Qin, L.; Li, X.; Yuan, M. Different Cell Wall-Degradation Ability Leads to Tissue-Specificity between Xanthomonas oryzae pv. oryzae and Xanthomonas oryzae pv. oryzicola. Pathogens 2020, 9, 187. https://doi.org/10.3390/pathogens9030187
Cao J, Chu C, Zhang M, He L, Qin L, Li X, Yuan M. Different Cell Wall-Degradation Ability Leads to Tissue-Specificity between Xanthomonas oryzae pv. oryzae and Xanthomonas oryzae pv. oryzicola. Pathogens. 2020; 9(3):187. https://doi.org/10.3390/pathogens9030187
Chicago/Turabian StyleCao, Jianbo, Chuanliang Chu, Meng Zhang, Limin He, Lihong Qin, Xianghua Li, and Meng Yuan. 2020. "Different Cell Wall-Degradation Ability Leads to Tissue-Specificity between Xanthomonas oryzae pv. oryzae and Xanthomonas oryzae pv. oryzicola" Pathogens 9, no. 3: 187. https://doi.org/10.3390/pathogens9030187