Isolation, Characterization, and Application of a Bacteriophage Infecting the Fish Pathogen Aeromonas hydrophila


 and
and 
Abstract
:1. Introduction
2. Results
2.1. Isolation and Characterization of the Bacteriophage
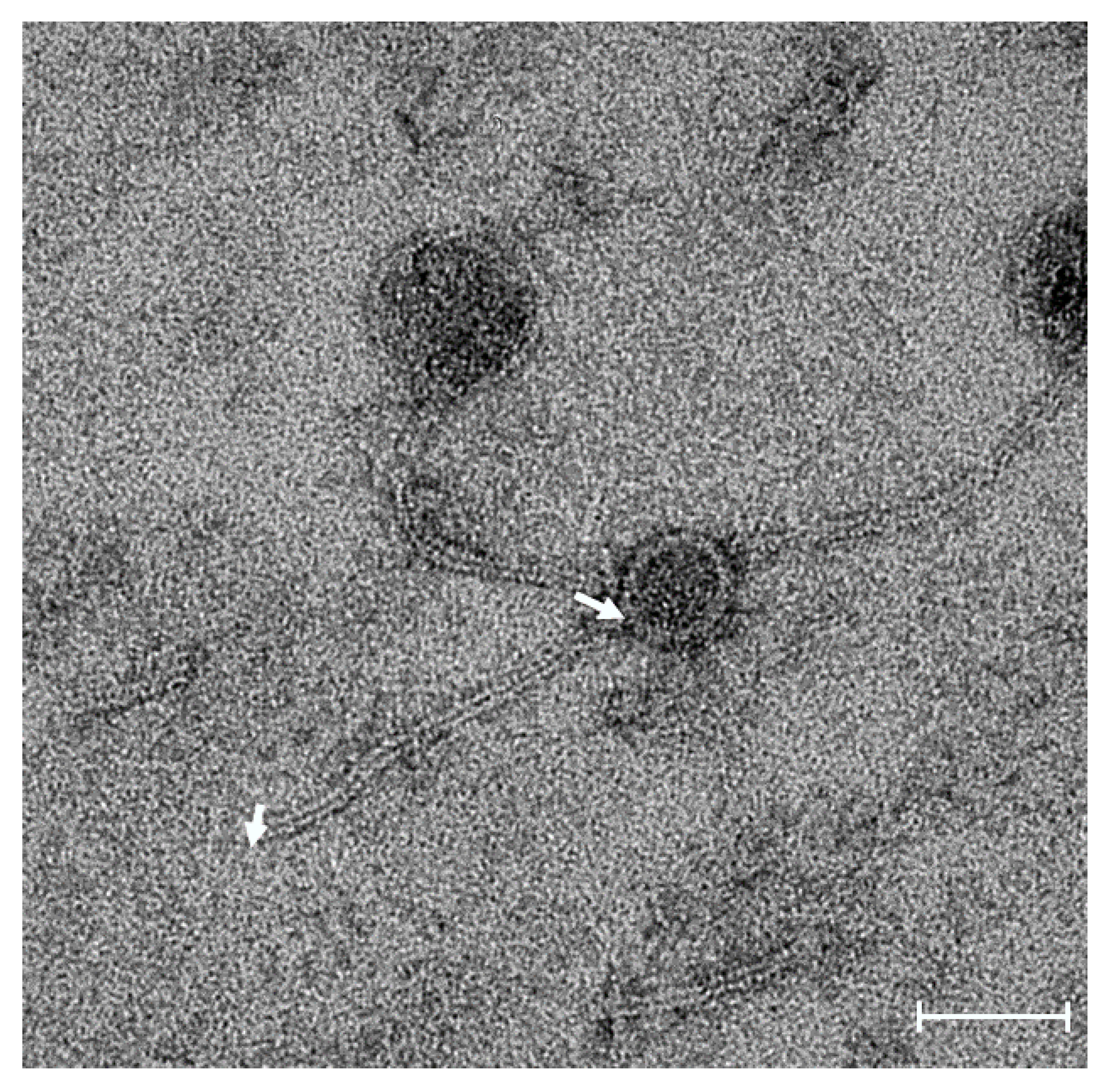
2.2. Specific Host Range and Morphology of Akh-2
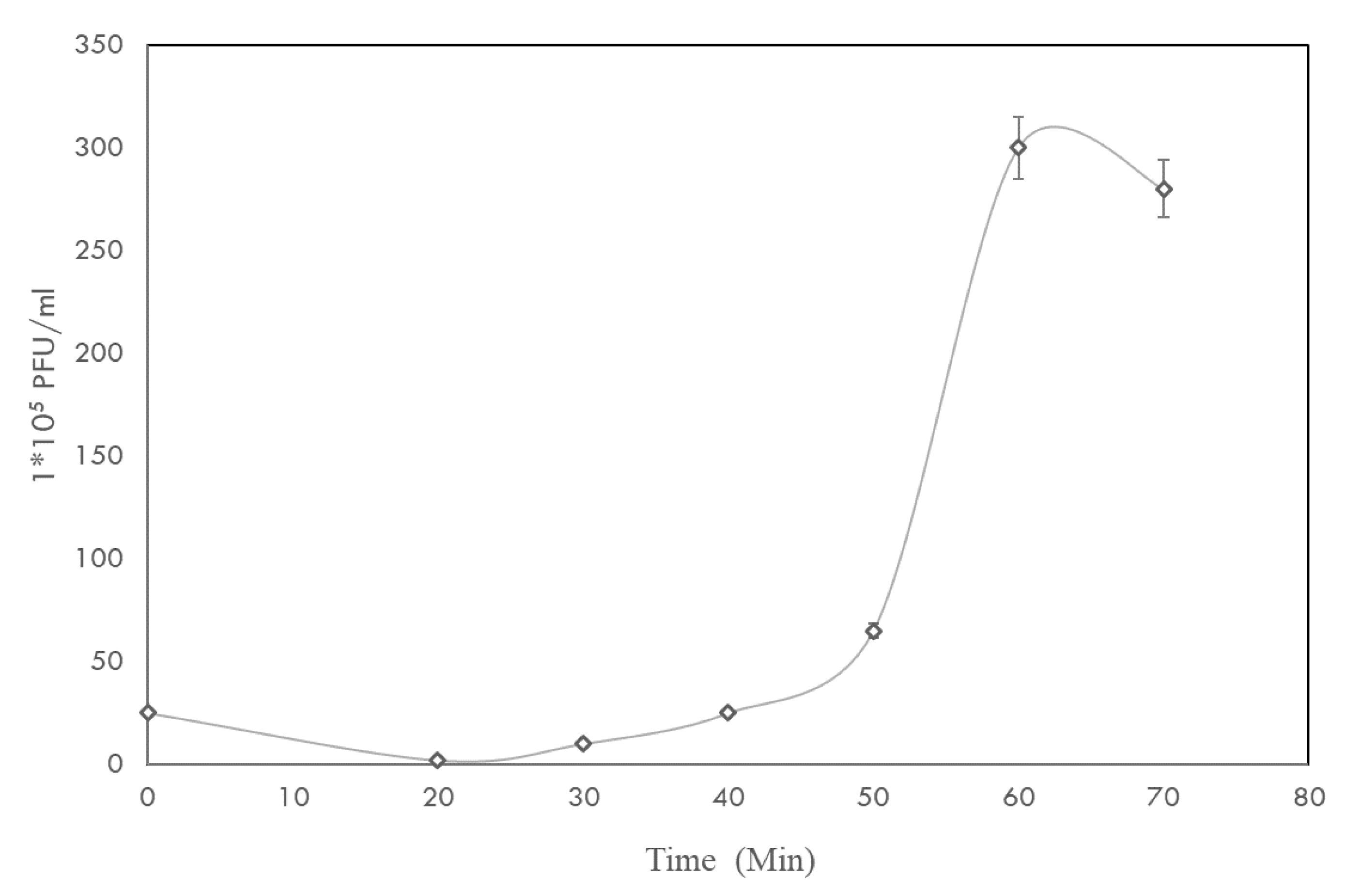
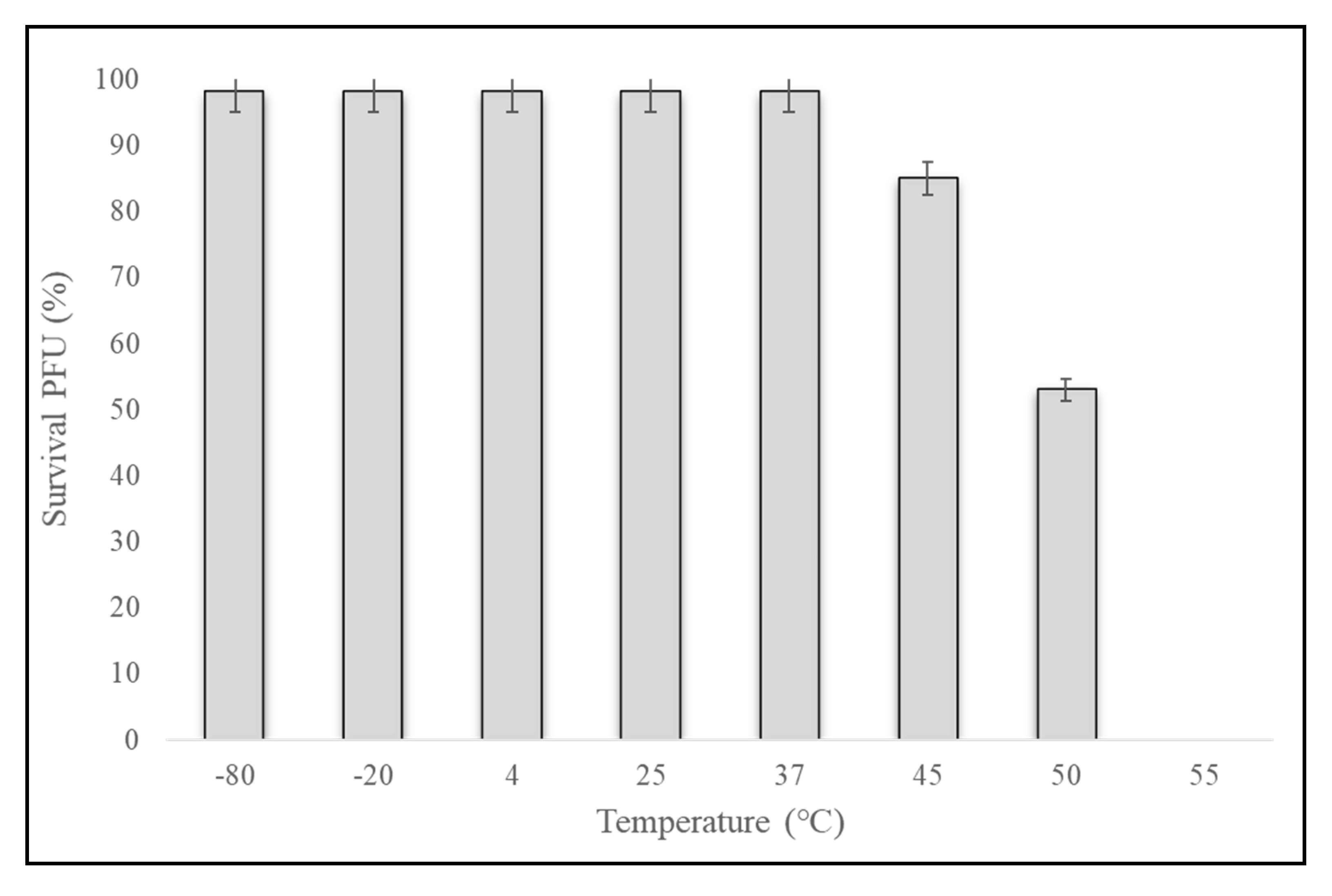
2.3. One-Step Growth Curve and Phage Stability
2.4. Genomic Analysis
2.5. Protective Effects of Akh-2
3. Discussion
4. Materials and Methods
4.1. Isolation, Culture, and Characterization of the Bacteriophage
4.2. Specific Host Range and Morphology of the Bacteriophage
4.3. One-Step Growth Curve and Stability Analyses
4.4. Genomic Analysis
4.4.1. Phage Genome Extraction
4.4.2. Whole-Genome Sequencing, Assembly, and Annotation
4.5. Protective Effects of Akh-2 in Infected Loach Fish
4.5.1. Challenge Test and Estimation of the Lethal Dose
4.5.2. Phage Treatment of Infected Fish
4.6. Statistical Analysis
4.7. Animal Experiment Approval
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- The State of World Fisheries and Aquaculture 2016; Contributing to food security and nutrition for all; FAO Fisheries and Aquaculture Department: Rome, Italy, 2016; ISBN 9789251091852.
- Luis, A.I.S.; Campos, E.V.R.; Oliveira, J.L.; Fraceto, L.F. Trends in aquaculture sciences: From now to use of nanotechnology for disease control. Rev. Aquac. 2017, 11, 119–132. [Google Scholar] [CrossRef] [Green Version]
- Jun, J.W.; Kim, J.H.; Shin, S.P.; Han, J.E.; Chai, J.Y.; Park, S.C. Protective effects of the Aeromonas phages pAh1-C and pAh6-C against mass mortality of the cyprinid loach (Misgurnus anguillicaudatus) caused by Aeromonas hydrophila. Aquaculture 2013, 416, 289–295. [Google Scholar] [CrossRef]
- Austin, B.; Austin, D.A. Aeromonadaceae Representative (Aeromonas salmonicida). In Bacterial Fish Pathogens; Springer: Dordrecht, The Netherlands, 2012; pp. 147–228. [Google Scholar]
- Lilley, J.; Hart, D.; Richards, R.H.; Roberts, R.J.; Cerenius, L.; Söderhäll, K. Pan-Asian spread of single fungal clone results in large scale fish kills. Veter Rec. 1997, 140, 653–654. [Google Scholar] [CrossRef]
- Kaskhedikar, M.; Chhabra, D. Multiple drug resistance in Aeromonas hydrophila isolates of fish. Food Microbiol. 2010, 28, 157–168. [Google Scholar]
- Altamirano, F.L.G.; Barr, J.J. Phage Therapy in the Postantibiotic Era. Clin. Microbiol. Rev. 2019, 32, e00066-18. [Google Scholar] [CrossRef] [Green Version]
- McCallin, S.; Sacher, J.C.; Zheng, J.; Chan, B.K. Current State of Compassionate Phage Therapy. Viruses 2019, 11, 343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, J.W.; Kim, J.H.; Gomez, D.; Choresca, C.H., Jr.; Han, J.E.; Shin, S.P.; Park, S.C. Occurrence of tetracycline-resistant Aeromonas hydrophila infection in Korean cyprinid loach (Misgurnus anguillicaudatus). African J. Microbiol. Res. 2010, 4, 849–855. [Google Scholar]
- Richards, G.P. Bacteriophage remediation of bacterial pathogens in aquaculture: A review of the technology. Bacteriophage 2014, 4, e975540. [Google Scholar] [CrossRef] [Green Version]
- Fauconnier, A. Phage Therapy Regulation: From Night to Dawn. Viruses 2019, 11, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazen, T.; Fliermans, C.B.; Hirsch, R.P.; Esch, G.W. Prevalence and distribution of Aeromonas hydrophila in the United States. Appl. Environ. Microbiol. 1978, 36, 731–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, M.; Cheng, Y.-H.; Sun, X.-Q.; Wang, Z.-Y.; Wang, Y.-X.; Cui, X.-L.; Xiao, W. Nine Novel Phages from a Plateau Lake in Southwest China: Insights into Aeromonas Phage Diversity. Viruses 2019, 11, 615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Easwaran, M.; Dananjaya, S.H.S.; Park, S.C.; Lee, J.; Shin, H.; De Zoysa, M. Characterization of bacteriophage pAh-1 and its protective effects on experimental infection of Aeromonas hydrophila in Zebrafish (Danio rerio). J. Fish Dis. 2016, 40, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Da, E.-A.; Th, G.E.-D.A.M.M. New Approach to Use Phage Therapy against Aeromonas hydrophila Induced Motile Aeromonas Septicemia in Nile Tilapia. J. Mar. Sci. Res. Dev. 2016, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Phumkhachorn, P.; Rattanachaikunsopon, P. A lytic podophage specific to fish pathogenic Edwardsiella tarda. Pak. J. Biotechnol. 2018, 15, 117–121. [Google Scholar]
- Vincent, A.T.; Paquet, V.E.; Bernatchez, A.; Tremblay, D.M.; Moineau, S.; Charette, S. Characterization and diversity of phages infecting Aeromonas salmonicida subsp. salmonicida. Sci. Rep. 2017, 7, 7054. [Google Scholar] [CrossRef] [Green Version]
- Haq, I.U.; Chaudhry, W.N.; Andleeb, S.; Qadri, I. Isolation and Partial Characterization of a Virulent Bacteriophage IHQ1 Specific for Aeromonas punctata from Stream Water. Microb. Ecol. 2011, 63, 954–963. [Google Scholar] [CrossRef]
- Luo, L.; Liao, G.; Liu, C.; Jiang, X.; Lin, M.; Zhao, C.; Tao, J.; Huang, Z. Characterization of bacteriophage HN48 and its protective effects in Nile tilapia Oreochromis niloticus against Streptococcus agalactiae infections. J. Fish Dis. 2018, 41, 1477–1484. [Google Scholar] [CrossRef]
- Yuan, S.; Chen, L.; Liu, Q.; Zhou, Y.; Yang, J.; Deng, D.; Li, H.; Ma, Y. Characterization and genomic analyses of Aeromonas hydrophila phages AhSzq-1 and AhSzw-1, isolates representing new species within the T5virus genus. Arch. Virol. 2018, 163, 1985–1988. [Google Scholar] [CrossRef]
- Park, S.C.; Shimamura, I.; Fukunaga, M.; Mori, K.-I.; Nakai, T. Isolation of Bacteriophages Specific to a Fish Pathogen, Pseudomonas plecoglossicida, as a Candidate for Disease Control. Appl. Environ. Microbiol. 2000, 66, 1416–1422. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Son, J.S.; Choi, Y.J.; Choresca, C.H.; Shin, S.P.; Han, J.E.; Jun, J.W.; Kang, D.H.; Oh, C.; Heo, S.J.; et al. Isolation and Characterization of a Lytic Myoviridae Bacteriophage PAS-1 with Broad Infectivity in Aeromonas salmonicida. Curr. Microbiol. 2012, 64, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Shivu, M.M.; Rajeeva, B.C.; Girisha, S.K.; Karunasagar, I.; Krohne, G.; Karunasagar, I. Molecular characterization of Vibrio harveyi bacteriophages isolated from aquaculture environments along the coast of India. Environ. Microbiol. 2007, 9, 322–331. [Google Scholar] [CrossRef]
- Weinbauer, M. Ecology of prokaryotic viruses. FEMS Microbiol. Rev. 2004, 28, 127–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jończyk, E.; Kłak, M.; Międzybrodzki, R.; Górski, A. The influence of external factors on bacteriophages—review. Folia Microbiol. 2011, 56, 191–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Choresca, C.H.; Shin, S.P.; Han, J.E.; Jun, J.W.; Park, S.C. Biological Control ofAeromonas salmonicidasubsp.salmonicidaInfection in Rainbow Trout (Oncorhynchus mykiss) UsingAeromonasPhage PAS-1. Transbound. Emerg. Dis. 2013, 62, 81–86. [Google Scholar] [CrossRef]
- Silva, Y.J.; Moreirinha, C.; Pereira, C.S.G.; Costa, L.; Rocha, R.; Cunha, A.; Gomes, N.C.M.; Calado, R.; Almeida, A. Biological control of Aeromonas salmonicida infection in juvenile Senegalese sole (Solea senegalensis) with Phage AS-A. Aquaculture 2016, 450, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Imbeault, S.; Parent, S.; Lagacé, M.; Uhland, C.F.; Blais, J.-F. Using Bacteriophages to Prevent Furunculosis Caused by Aeromonas salmonicida in Farmed Brook Trout. J. Aquat. Anim. Health 2006, 18, 203–214. [Google Scholar] [CrossRef]
- Zhang, D.; Xu, D.; Shoemaker, C. Experimental induction of motile Aeromonas septicemia in channel catfish ( Ictalurus punctatus ) by waterborne challenge with virulent Aeromonas hydrophila. Aquac. Rep. 2016, 3, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Verner–Jeffreys, D.W.; Algoet, M.; Pond, M.J.; Virdee, H.K.; Bagwell, N.J.; Roberts, E.G. Furunculosis in Atlantic salmon (Salmo salar L.) is not readily controllable by bacteriophage therapy. Aquaculture 2007, 270, 475–484. [Google Scholar]
- Horvath, P.; Barrangou, R. CRISPR/Cas, the Immune System of Bacteria and Archaea. Science 2010, 327, 167–170. [Google Scholar] [CrossRef] [Green Version]
- Jun, S.Y.; Jung, G.M.; Yoon, S.J.; Oh, M.-D.; Choi, Y.-J.; Lee, W.J.; Kong, J.-C.; Seol, J.G.; Kang, S.H. Antibacterial properties of a pre-formulated recombinant phage endolysin, SAL-1. Int. J. Antimicrob. Agents 2013, 41, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yuan, S.; Liu, Q.; Mai, G.; Yang, J.; Deng, D.; Zhang, B.; Liu, C.; Ma, Y. In Vitro Design and Evaluation of Phage Cocktails Against Aeromonas salmonicida. Front. Microbiol. 2018, 9, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, S.W.M.; Ali, S.M.; Almisherfi, M.M. Isolation and Molecular Characterization of Some Marine Aeromonas phages: Protective Effects for Nile Tilapia Infected with Aeromonas hydrophila. J. Pure Appl. Microbiol. 2018, 12, 1175–1185. [Google Scholar] [CrossRef]
- Gencay, Y.E.; Birk, T.; Sørensen, M.C.H.; Brøndsted, L. Methods for Isolation, Purification, and Propagation of Bacteriophages of Campylobacter jejuni. In Methods in Molecular Biology; Butcher, J., Stintzi, A., Eds.; Humana Press: New York, NY, USA, 2017; Volume 1512, pp. 19–28. [Google Scholar]
- Verma, V.; Harjai, K.; Chhibber, S. Characterization of a T7-Like Lytic Bacteriophage of Klebsiella pneumoniae B5055: A Potential Therapeutic Agent. Curr. Microbiol. 2009, 59, 274–281. [Google Scholar] [CrossRef]
- RAST. Available online: https://rast.theseed.org/FIG/rast.cgi. (accessed on 18 July 2018).
- Sullivan, M.J.; Petty, N.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Strain ID | Lysis by Akh-2 | Source * |
|---|---|---|---|
| Aeromonas hydrophila | KTCC 2358 | YES | 1 |
| KCCM 32586 | NO | 2 | |
| AH-A6 | YES | 3 | |
| AH-A8 | NO | 3 | |
| AH-20 | YES | 3 | |
| AH-21 | NO | 3 | |
| AH-Juwah | YES | 3 | |
| Pectobacterium spp. | 35, 48, 63, 92, E42, E44 | NO | 4 |
| Streptococcus aureus | S75, S86, S103, S106 | NO | 4 |
| Bacillus spp. | B1, B2, B3, B4, B5, B6, B8, B11, B12, B13, B87, B97 | NO | 4 |
| Escherichia coli | CJY H7 | NO | 4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akmal, M.; Rahimi-Midani, A.; Hafeez-ur-Rehman, M.; Hussain, A.; Choi, T.-J. Isolation, Characterization, and Application of a Bacteriophage Infecting the Fish Pathogen Aeromonas hydrophila. Pathogens 2020, 9, 215. https://doi.org/10.3390/pathogens9030215
Akmal M, Rahimi-Midani A, Hafeez-ur-Rehman M, Hussain A, Choi T-J. Isolation, Characterization, and Application of a Bacteriophage Infecting the Fish Pathogen Aeromonas hydrophila. Pathogens. 2020; 9(3):215. https://doi.org/10.3390/pathogens9030215
Chicago/Turabian StyleAkmal, Muhammad, Aryan Rahimi-Midani, Muhammad Hafeez-ur-Rehman, Ali Hussain, and Tae-Jin Choi. 2020. "Isolation, Characterization, and Application of a Bacteriophage Infecting the Fish Pathogen Aeromonas hydrophila" Pathogens 9, no. 3: 215. https://doi.org/10.3390/pathogens9030215