miRNAs: EBV Mechanism for Escaping Host’s Immune Response and Supporting Tumorigenesis


Abstract
:1. Introduction
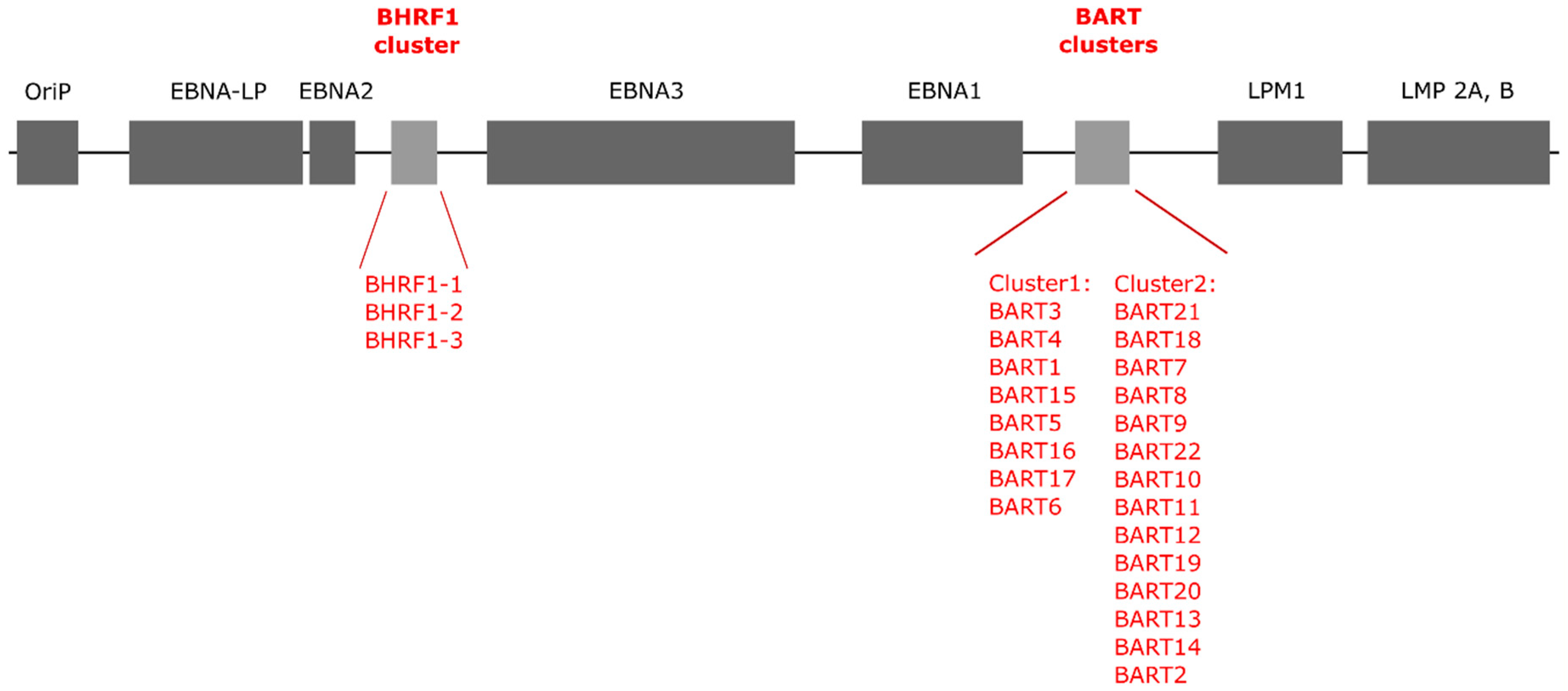
2. EBV miRNAs
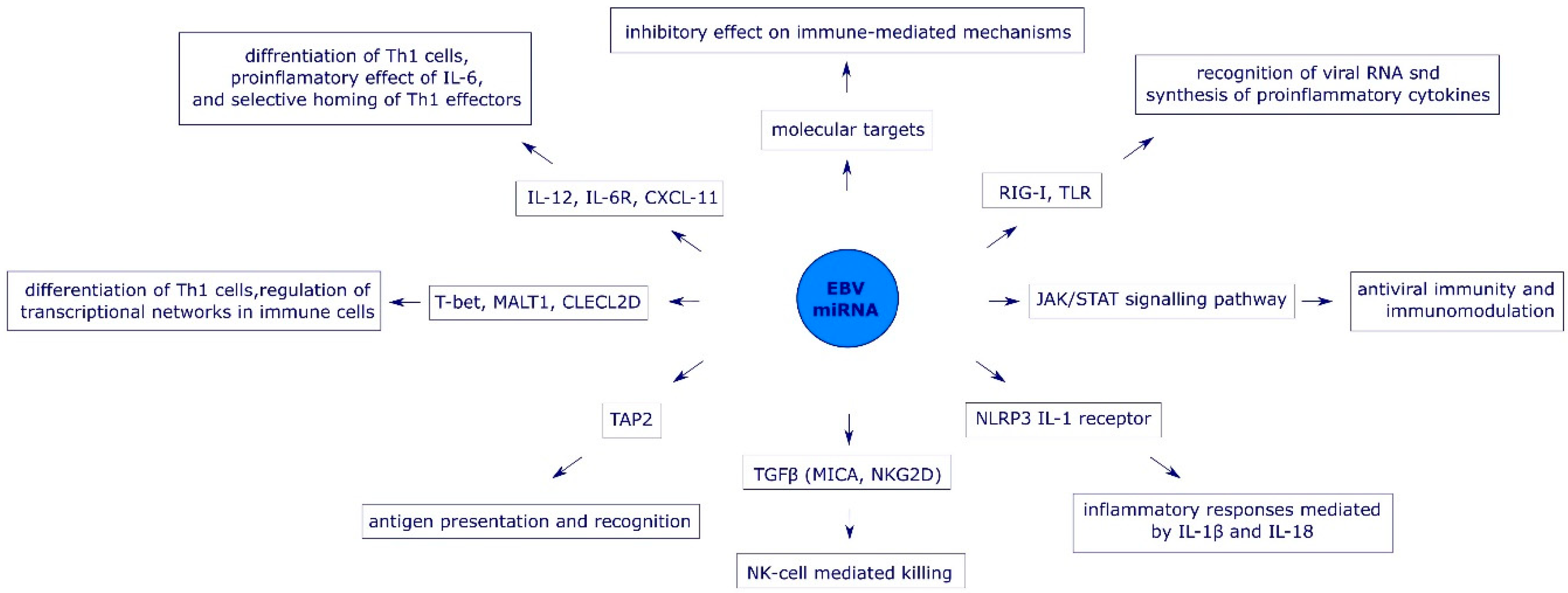
3. The Role of EBV miRNAs in Immune Evasion
3.1. Pattern-Recognition Receptor-Mediated Signaling Pathways and Interferons
3.2. Natural Killer (NK) Cells
3.3. Inflammasome
3.4. Cytokines and Chemokines
3.5. Antigen Presentation
3.6. Specific Cellular Immunity
4. The Role of EBV miRNAs in Tumorigenesis
4.1. EBV miRNAs in Lymphoma
4.2. EBV miRNAs in Carcinoma
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- International Committee on Taxonomy of Viruses ICTV (2019): ICTV Taxonomy History: Human Gammaherpesvirus 4. Available online: https://talk.ictvonline.org/taxonomy/p/taxonomy-history?taxnode_id=20171486 (accessed on 1 October 2019).
- Epstein, M.A.; Achong, B.G.; Barr, Y.M. Virus particles in cultured lymphoblasts from Burkitt’s lymphoma. Lancet 1964, 1, 702–703. [Google Scholar] [CrossRef]
- Pope, J.H. Establishment of Cell Lines from Peripheral Leucocytes in Infectious Mononucleosis. Nature 1967, 216, 810–811. [Google Scholar] [CrossRef]
- Henle, W.; Diehl, V.; Kohn, G.; Hausen, H.Z.; Henle, G. Herpes-type virus and chromosome marker in normal leukocytes after growth with irradiated Burkitt cells. Science 1967, 157, 1064–1065. [Google Scholar] [CrossRef]
- Young, L.S.; Yap, F.L.; Murray, P.G. Epstein-Barr virus: More than 50 years old and still providing surprises. Nat. Rev. Cancer 2016, 16, 789–802. [Google Scholar] [CrossRef]
- Thorley-Lawson, D.A. EBV Persistence-Introducing the Virus. Curr. Top. Microbiol. Immunol. 2015, 390 Pt 1, 151–209. [Google Scholar]
- Saha, A.; Robertson, E.S. Mechanisms of B-Cell Oncogenesis Induced by Epstein-Barr Virus. J. Virol. 2019, 93, e00238–e00319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsueh, C.Y.; Yang, C.F.; Gau, J.P.; Kuan, E.C.; Ho, C.Y.; Chiou, T.J.; Hsiao, L.T.; Lin, T.A.; Lan, M.Y. Nasopharyngeal lymphoma: A 22-year review of 35 cases. J. Clin. Med. 2019, 8, 1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, R.; Kumar, A.; Ingle, H.; Kumar, H. The Interplay Between Viral-Derived miRNAs and Host Immunity During Infection. Front. Immunol. 2020, 10, 3079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigeyoshi, F.; Nakamura, H. Animal Models for Gammaherpesvirus Infections: Recent Development in the Analysis of Virus-Induced Pathogenesis. Pathogens 2020, 9, 116. [Google Scholar]
- Gallo, A.; Miceli, V.; Bulati, M.; Iannolo, G.; Contino, F.; Conaldi, P.G. Viral miRNAs as Active Players and Participants in Tumorigenesis. Rev. Cancers 2020, 12, 358. [Google Scholar] [CrossRef] [Green Version]
- Pfeffer, S.; Zavolan, M.; Grässer, F.A.; Chien, M.; Russo, J.J.; Ju, J.; John, B.; Enright, A.J.; Marks, D.; Sander, C.; et al. Identification of virus-encoded microRNAs. Science 2004, 304, 734–736. [Google Scholar] [CrossRef] [PubMed]
- Mui, U.N.; Haley, C.T.; Tyring, S.K. Viral Oncology: Molecular Biology and Pathogenesis. J. Clin. Med. 2017, 6, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, S.; Endo, K.; Wakisaka, N.; Aga, M.; Kano, M.; Seishima, N.; Imoto, T.; Kobayashi, E.; Moriyama-Kita, M.; Nakanishi, Y.; et al. Expression of interferon regulatory factor 7 correlates with the expression of Epstein-Barr Virus latent membrane protein 1 and cervical lymph node metastasis in nasopharyngeal cancer. Pathol. Int. 2017, 67, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Chen, J.N.; Qiu, X.M.; Pan, Y.H.; Zhang, Z.G.; Shao, C.K. Comparative analysis of 22 Epstein-Barr virus genomes from diseased and healthy individuals. J. Gen. Virol. 2017, 98, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Baer, R.; Bankier, A.T.; Biggin, M.D.; Deininger, P.L.; Farrell, P.J.; Gibson, T.J.; Hatfull, G.; Hudson, G.S.; Satchwell, S.C.; Séguin, C.; et al. DNA sequence and expression of the B95-8 Epstein-Barr virus genome. Nature 1984, 310, 207–211. [Google Scholar] [CrossRef]
- Human Gammaherpesvirus 4, Complete Genome. Available online: https://www.ncbi.nlm.nih.gov/nuccore/NC_007605 (accessed on 1 October 2019).
- Barth, S.; Meister, G.; Grasser, F.A. EBV-encoded miRNAs. Biochim. Biophys. Acta 2011, 1809, 631–640. [Google Scholar] [CrossRef]
- Cai, X.; Schäfer, A.; Lu, S.; Bilello, J.P.; Desrosiers, R.C.; Edwards, R.; Raab-Traub, N.; Cullen, B.R. Epstein-Barr virus microRNAs are evolutionarily conserved and differentially expressed. PLoS Pathog. 2006, 2, e23. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Yu, F.; Wu, W.; Wang, Y.; Ding, H.; Qian, L. Epstein-Barr virus-encoded microRNAs as regulators in host immune responses. Int. J. Biol. Sci. 2018, 14, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Zanella, L.; Riquelme, I.; Buchegger, K.; Abanto, M.; Ili, C.; Brebi, P. A reliable Epstein-Barr Virus classification based on phylogenomic and population analyses. Sci. Rep. 2019, 9, 9829. [Google Scholar] [CrossRef] [Green Version]
- Pratt, Z.L.; Kuzembayeva, M.; Sengupta, S.; Sugden, B. The microRNAs of Epstein-Barr Virus are expressed at dramatically differing levels among cell lines. Virology 2009, 386, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Navari, M.; Etebari, M.; Ibrahimi, M.; Leoncini, L.; Piccaluga, P.P. Pathobiologic Roles of Epstein–Barr Virus-Encoded MicroRNAs in Human Lymphomas. Int. J. Mol. Sci. 2018, 19, 1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, H.; Lin, X.; Shumilov, A.; Bernhardt, K.; Feederle, R.; Poirey, R.; Kopp-Schneider, A.; Pereira, B.; Almeida, R.; Delecluse, H.J. The biological properties of different Epstein-Barr virus strains explain their association with various types of cancers. Oncotarget 2017, 8, 10238–10254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correia, S.; Palser, A.; Elgueta Karstegl, C.; Middeldorp, J.M.; Ramayanti, O.; Cohen, J.I.; Hildesheim, A.; Fellner, M.D.; Wiels, J.; White, R.E.; et al. Natural Variation of Epstein-Barr Virus Genes, Proteins, and Primary MicroRNA. J. Virol. 2017, 91, e00375–e00417. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.N.; Zhou, L.; Qiu, X.M.; Yang, R.H.; Liang, J.; Pan, Y.H.; Li, H.F.; Peng, G.R.; Shao, C.K. Determination and genome-wide analysis of Epstein-Barr virus (EBV) sequences in EBV-associated gastric carcinoma from Guangdong, an endemic area of nasopharyngeal carcinoma. J. Med. Microbiol. 2018, 67, 1614–1627. [Google Scholar] [CrossRef]
- Sunagawa, K.; Hishima, T.; Fukumoto, H.; Hasegawa, H.; Katano, H. Conserved sequences of bart and bhrf regions encoding viral microRNAs in epstein–barr virus-associated lymphoma. BMC Res. Notes 2017, 10, 279. [Google Scholar] [CrossRef] [Green Version]
- Zuo, L.; Yue, W.; Du, S.; Xin, S.; Zhang, J.; Liu, L.; Li, G.; Lu, J. An update: Epstein-Barr virus and immune evasion via microRNA regulation. Virol. Sin. 2017, 32, 175–187. [Google Scholar] [CrossRef]
- Jangra, S.; Yuen, K.S.; Botelho, M.G.; Jin, D.Y. Epstein–Barr Virus and Innate Immunity: Friends or Foes? Microorganisms 2019, 7, 183. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Qin, Z.; Wang, J.; Zheng, X.; Lu, J.; Zhang, X.; Wei, L.; Peng, Q.; Zheng, Y.; Ou, C.; et al. Epstein-Barr virus miR-BART6-3p inhibits the RIG-I pathway. J. Innate Immun. 2017, 9, 574–586. [Google Scholar] [CrossRef]
- Owen, K.L.; Brockwell, N.K.; Parker, B.S. JAK-STAT Signaling: A Double-Edged Sword of Immune Regulation and Cancer Progression. Cancers 2019, 11, 2002. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.T.; Lin, C.W. EBV-encoded miR-BART20-5p and miR-BART8 inhibit the IFN-gamma-STAT1 pathway associated with disease progression in nasal NK-cell lymphoma. Am. J. Pathol. 2014, 184, 1185–1197. [Google Scholar] [CrossRef]
- Wong, T.S.; Chen, S.; Zhang, M.J.; Chan, J.Y.; Gao, W. Epstein-Barr virus-encoded microRNA BART7 downregulates major histocompatibility complex class I chain-related peptide A and reduces the cytotoxicity of natural killer cells to nasopharyngeal carcinoma. Oncol. Lett. 2018, 16, 2887–2892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skinner, C.M.; Ivanov, N.S.; Barr, S.A.; Chen, Y.; Skalsky, R.L. An Epstein-Barr Virus MicroRNA Blocks Interleukin-1 (IL-1) Signaling by Targeting IL-1 Receptor 1. J. Virol. 2017, 91, e00530–e00617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haneklaus, M.; Gerlic, M.; Kurowska-Stolarska, M.; Rainey, A.A.; Pich, D.; McInnes, I.B.; Hammerschmidt, W.; O’Neill, L.A.; Masters, S.L. Cutting edge: miR-223 and EBV miR-BART15 regulate the NLRP3 inflammasome and IL-1 production. J. Immunol. 2012, 189, 3795–3799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Cao, W.; Zhu, Y. Immunoregulatory Functions of the IL-12 Family of Cytokines in Antiviral Systems. Viruses 2019, 11, 772. [Google Scholar] [CrossRef] [Green Version]
- Albanese, M.; Tagawa, T.; Bouvet, M.; Maliqi, L.; Lutter, D.; Hoser, J.; Hastreiter, M.; Hayes, M.; Sugden, B.; Martin, L.; et al. Epstein-Barr virus microRNAs reduce immune surveillance by virus-specific CD8+ T cells. Proc. Natl. Acad. Sci. USA 2016, 113, e6467–e6475. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.M.; Yu, Y.; Zhao, H.P. EBVBART63p and cellular microRNA197 compromise the immune defense of host cells in EBV-positive Burkitt lymphoma. Mol. Med. Rep. 2017, 15, 1877–1883. [Google Scholar] [CrossRef]
- Xia, T.; O’Hara, A.; Araujo, I.; Barreto, J.; Carvalho, E.; Sapucaia, J.B.; Ramos, J.C.; Luz, E.; Pedroso, C.; Manrique, M.; et al. EBV microRNAs in primary lymphomas and targeting of CXCL-11 by ebv-mir-BHRF1-3. Cancer Res. 2008, 68, 1436–1442. [Google Scholar] [CrossRef] [Green Version]
- Tagawa, T.; Albanese, M.; Bouvet, M.; Moosmann, A.; Mautner, J.; Heissmeyer, V.; Zielinkski, C. Epstein-Barr viral miRNAs inhibit antiviral CD4+ T cell responses targeting IL-12 and peptide processing. J. Exp. Med. 2016, 213, 2065–2080. [Google Scholar] [CrossRef]
- Skalsky, R.L.; Corcoran, D.L.; Gottwein, E.; Frank, C.L.; Kang, D.; Hafner, M.; Nusbaum, J.D. The viral and cellular microRNA targetome in lymphoblastoid cell lines. PLoS Pathog. 2012, 8, e1002484. [Google Scholar] [CrossRef]
- Gurer, C.; Strowig, T.; Brilot, F.; Pack, M.; Trumpfheller, C.; Arrey, F.; Park, C.G. Targeting the nuclear antigen 1 of Epstein-Barr virus to the human endocytic receptor DEC-205 stimulates protective T-cell responses. Blood 2008, 112, 1231–1239. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.C.; Liu, T.Y.; Hsu, S.M.; Lin, C.W. Epstein-Barr virus-encoded miR-BART20-5p inhibits T-bet translation with secondary suppression of p53 in invasive nasal NK/T-cell lymphoma. Am. J. Pathol. 2013, 182, 1865–1875. [Google Scholar] [CrossRef] [PubMed]
- Lazarevic, V.; Szabo, S.; Glimcher, L.H. T-bet runs INTERFERence. Immunity 2017, 46, 968–970. [Google Scholar] [CrossRef] [PubMed]
- Lazarević, V.; Glimcher, L.H.; Lord, G.M. T-bet: A bridge between innate and adaptive immunity. Nat. Rev. Immunol. 2013, 13, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Fink, S.E.K.; Gandhi, M.K.; Nourse, J.P.; Keane, C.; Jones, K.; Crooks, P.; Jöhrens, K.; Korfel, A.; Schmidt, H.; Neumann, S.; et al. A Comprehensive Analysis of the Cellular and EBV-Specific MicroRNAome in Primary CNS PTLD Identifies Different Patterns Among EBV-Associated Tumors. Am. J. Transplant. 2014, 14, 2577–2587. [Google Scholar] [CrossRef] [PubMed]
- Imig, J.; Motsch, N.; Zhu, J.Y.; Barth, S.; Okoniewski, M.; Reineke, T.; Tinguely, M.; Faggioni, A.; Trivedi, P.; Meister, G.; et al. microRNA profiling in Epstein–Barr virus-associated B-cell lymphoma. Nucleic Acids Res. 2011, 39, 1880–1893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambrosio, M.R.; Navari, M.; Di Lisio, L.; Leon, E.A.; Onnis, A.; Gazaneo, S.; Mundo, L.; Ulivieri, C.; Gomez, G.; Lazzi, S.; et al. The epstein–barr-encoded BART-6-3p microrna affects regulation of cell growth and immuno response in burkitt lymphoma. Infect. Agent Cancer 2014, 9, 12. [Google Scholar] [CrossRef] [Green Version]
- Motsch, N.; Alles, J.; Imig, J.; Zhu, J.; Barth, S.; Reineke, T.; Tinguely, M.; Cogliatti, S.; Dueck, A.; Meister, G.; et al. MicroRNA Profiling of Epstein-Barr Virus-Associated NK/T-Cell Lymphomas by Deep Sequencing. PLoS ONE 2012, 7, e42193. [Google Scholar] [CrossRef]
- Zhu, J.Y.; Pfuhl, T.; Motsch, N.; Barth, S.; Nicholls, J.; Grässer, F.; Meister, G. Identification of Novel Epstein-Barr Virus MicroRNA Genes from Nasopharyngeal Carcinomas. J. Virol. 2009, 83, 3333–3341. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Bu, Y.; Liang, Y.; Zhang, F.; Zhang, H.; Li, S. Epstein-Barr Virus (EBV)-BamHI-A Rightward Transcript (BART)-6 and Cellular MicroRNA-142 Synergistically Compromise Immune Defense of Host Cells in EBV-Positive Burkitt Lymphoma. Med. Sci. Monit. 2016, 22, 4114–4120. [Google Scholar] [CrossRef] [Green Version]
- Gilligan, K.; Sato, H.; Rajadurai, P.; Busson, P.; Young, L.; Rickinson, A.; Tursz, T.; Raab-Traub, N. Novel transcription from the Epstein-Barr virus terminal EcoRI fragment, DIJhet, in a nasopharyngeal carcinoma. J. Virol. 1990, 64, 4948–4956. [Google Scholar] [CrossRef] [Green Version]
- Hitt, M.M.; Allday, M.J.; Hara, T.; Karran, L.; Jones, M.D.; Busson, P.; Tursz, T.; Ernberg, I.; Griffin, B.E. EBV gene expression in an NPC-related tumour. EMBO J. 1989, 8, 2639–2651. [Google Scholar] [CrossRef] [PubMed]
- Gilligan, K.J.; Rajadurai, P.; Lin, J.C.; Busson, P.; Abdel-Hamid, M.; Prasad, U.; Tursz, T.; Raab-Traub, N. Expression of the Epstein-Barr virus BamHI A fragment in nasopharyngeal carcinoma: Evidence for a viral protein expressed in vivo. J. Virol. 1991, 65, 6252–6259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Guo, Z.; Shu, Y.; Zhou, H.; Wang, H.; Zhang, W. BART miRNAs: An unimaginable force in the development of nasopharyngeal carcinoma. Eur. J. Cancer Prev. 2017, 26, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.X.; Yi, H.; Qu, J.Q.; He, Q.Y.; Xiao, Z.Q. Integrated analysis of the differential cellular and EBV miRNA expression profiles in microdissected nasopharyngeal carcinoma and non-cancerous nasopharyngeal tissues. Oncol. Rep. 2015, 34, 2585–2601. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Huang, H.; Huang, L.; Sun, M.; Yan, Q.; Song, Y.; Wei, F.; Bo, H.; Gong, Z.; Zeng, Y.; et al. Regulation network and expression profiles of Epstein-Barr virus-encoded microRNAs and their potential target host genes in nasopharyngeal carcinomas. Sci. China Life Sci. 2014, 57, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Wong, A.M.; Kong, K.L.; Tsang, J.W.; Kwong, D.L.; Guan, X.Y. Profiling of Epstein-Barr virus-encoded microRNAs in nasopharyngeal carcinoma reveals potential biomarkers and oncomirs. Cancer 2012, 118, 698–710. [Google Scholar] [CrossRef]
- Cai, L.; Ye, Y.; Jiang, Q.; Chen, Y.; Lyu, X.; Li, J.; Wang, S.; Liu, T.; Cai, H.; Yao, K.; et al. Epstein-Barr virus-encoded microRNA BART1 induces tumour metastasis by regulating PTEN-dependent pathways in nasopharyngeal carcinoma. Nat. Commun. 2015, 6, 7353. [Google Scholar] [CrossRef] [Green Version]
- Cai, L.M.; Lyu, X.M.; Luo, W.R.; Cui, X.F.; Ye, Y.F.; Yuan, C.C.; Peng, Q.X.; Wu, D.H.; Liu, T.F.; Wang, E.; et al. EBV-miR-BART7-3p promotes the EMT and metastasis of nasopharyngeal carcinoma cells by suppressing the tumor suppressor PTEN. Oncogene 2015, 34, 2156–2166. [Google Scholar] [CrossRef] [Green Version]
- Lyu, X.; Wang, J.; Guo, X.; Wu, G.; Jiao, Y.; Faleti, O.D.; Liu, P.; Liu, T.; Long, Y.; Chong, T.; et al. EBV-miR-BART1-5P activates AMPK/mTOR/HIF1 pathway via a PTEN independent manner to promote glycolysis and angiogenesis in nasopharyngeal carcinoma. PLoS Pathog. 2018, 14, e1007484. [Google Scholar] [CrossRef] [Green Version]
- Lo, A.K.; To, K.F.; Lo, K.W.; Lung, R.W.; Hui, J.W.; Liao, G.; Hayward, S.D. Modulation of LMP1 protein expression by EBV-encoded microRNAs. Proc. Natl. Acad. Sci. USA 2007, 104, 16164–16169. [Google Scholar] [CrossRef] [Green Version]
- Kang, D.; Skalsky, R.L.; Cullen, B.R. EBV BART MicroRNAs Target Multiple Pro-apoptotic Cellular Genes to Promote Epithelial Cell Survival. PLoS Pathog. 2015, 11, e1004979. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Yuen, K.S.; Xu, R.; Tsao, S.W.; Chen, H.; Li, M.; Kok, K.H.; Jin, D.Y. Targeting of DICE1 tumor suppressor by Epstein-Barr virus-encoded miR-BART3* microRNA in nasopharyngeal carcinoma. Int. J. Cancer 2013, 133, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki-Ushiku, A.; Kunita, A.; Isogai, M.; Hibiya, T.; Ushiku, T.; Takada, K.; Fukayama, M. Profiling of Virus-Encoded MicroRNAs in Epstein-Barr Virus-Associated Gastric Carcinoma and Their Roles in Gastric Carcinogenesis. J. Virol. 2015, 89, 5581–5591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choy, E.Y.; Siu, K.L.; Kok, K.H.; Lung, R.W.; Tsang, C.M.; To, K.F.; Kwong, D.L.; Tsao, S.W.; Jin, D.Y. An Epstein-Barr virus-encoded microRNA targets PUMA to promote host cell survival. J. Exp. Med. 2008, 205, 2551–2560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Wang, J.; Wei, L.; Peng, Q.; Gao, Y.; Fu, Y.; Lu, Y.; Qin, Z.; Zhang, X.; Lu, J.; et al. Epstein-Barr Virus MicroRNA miR-BART5-3p Inhibits p53 Expression. J. Virol. 2018, 92, e01022-18. [Google Scholar] [CrossRef] [Green Version]
- Lung, R.W.; Hau, P.M.; Yu, K.H.; Yip, K.Y.; Tong, J.H.; Chak, W.P.; Chan, A.W.; Lam, K.H.; Lo, A.K.; Tin, E.K.; et al. EBV-encoded miRNAs target ATM-mediated response in nasopharyngeal carcinoma. J. Pathol. 2018, 244, 394–407. [Google Scholar] [CrossRef]
- He, B.; Li, W.; Wu, Y.; Wei, F.; Gong, Z.; Bo, H.; Wang, Y.; Li, X.; Xiang, B.; Guo, C.; et al. Epstein-Barr virus-encoded miR-BART6-3p inhibits cancer cell metastasis and invasion by targeting long non-coding RNA LOC553103. Cell Death Dis. 2016, 7, e2353. [Google Scholar] [CrossRef]
- Iizasa, H.; Wulff, B.E.; Alla, N.R.; Maragkakis, M.; Megraw, M.; Hatzigeorgiou, A.; Iwakiri, D.; Takada, K.; Wiedmer, A.; Showe, L.; et al. Editing of Epstein-Barr virus-encoded BART6 microRNAs controls their dicer targeting and consequently affects viral latency. J. Biol. Chem. 2010, 285, 33358–33370. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Zong, J.; Lin, W.; Wang, M.; Xu, Y.; Zhou, R.; Lin, S.; Guo, Q.; Chen, H.; Ye, Y.; et al. EBV-miR-BART8-3p induces epithelial-mesenchymal transition and promotes metastasis of nasopharyngeal carcinoma cells through activating NF-κB and Erk1/2 pathways. J. Exp. Clin. Cancer Res. 2018, 37, 283. [Google Scholar] [CrossRef]
- Hsu, C.Y.; Yi, Y.H.; Chang, K.P.; Chang, Y.S.; Chen, S.J.; Chen, H.C. The Epstein-Barr virus-encoded microRNA MiR-BART9 promotes tumor metastasis by targeting E-cadherin in nasopharyngeal carcinoma. PLoS Pathog. 2014, 10, e1003974. [Google Scholar] [CrossRef]
- Yan, Q.; Zeng, Z.; Gong, Z.; Zhang, W.; Li, X.; He, B.; Song, Y.; Li, Q.; Zeng, Y.; Liao, Q.; et al. EBV-miR-BART10-3p facilitates epithelial-mesenchymal transition and promotes metastasis of nasopharyngeal carcinoma by targeting BTRC. Oncotarget 2015, 6, 41766–41782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, K.; Lee, S.K. EBV miR-BART10-3p Promotes Cell Proliferation and Migration by Targeting DKK1. Int. J. Biol. Sci. 2019, 15, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Li, X.; Zeng, Z.; Li, Q.; Gong, Z.; Liao, Q.; Li, X.; Chen, P.; Xiang, B.; Zhang, W.; et al. Epstein-Barr virus encoded miR-BART11 promotes inflammation-induced carcinogenesis by targeting FOXP1. Oncotarget 2016, 7, 36783–36799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.J.; Zhou, R.; Zong, J.F.; Lin, W.S.; Tong, S.; Guo, Q.J.; Lin, C.; Lin, S.J.; Chen, Y.X.; Chen, M.R.; et al. Epstein-Barr virus-coded miR-BART13 promotes nasopharyngeal carcinoma cell growth and metastasis via targeting of the NKIRAS2/NF-κB pathway. Cancer Lett. 2019, 447, 33–40. [Google Scholar] [CrossRef]
- Choy, H.; Lee, H.; Kim, S.R.; Gho, Y.S.; Lee, S.K. Epstein-Barr virus-encoded microRNA BART15-3p promotes cell apoptosis partially by targeting BRUCE. J. Virol. 2013, 87, 8135–8144. [Google Scholar] [CrossRef] [Green Version]
- Kanda, T.; Miyata, M.; Kano, M.; Kondo, S.; Yoshizaki, T.; Iizasa, H. Clustered microRNAs of the Epstein-Barr virus cooperatively downregulate an epithelial cell-specific metastasis suppressor. J. Virol. 2015, 89, 2684–2697. [Google Scholar] [CrossRef] [Green Version]
- Babu, S.G.; Ponia, S.S.; Kumar, D.; Saxena, S. Cellular oncomiR orthologue in EBV oncogenesis. Comput. Biol. Med. 2011, 41, 891–898. [Google Scholar] [CrossRef]
- Skalsky, R.L.; Cullen, B.R. EBV Noncoding RNAs. Curr. Top. Microbiol. Immunol. 2015, 391, 181–217. [Google Scholar]



{kind=link}
{kind=link}
| BART | BART Target | Affected Process | Study | Method |
|---|---|---|---|---|
| 1 | PTEN | migration, signaling | Cai et al. 2015a, Cai et al. 2015b [59,60] | reporter assay |
| 1-5p | α1subunit AMPK | metabolism | Lyu et al. 2018 [61] | reporter assay, biopsies, cell culture |
| 1-5p | LMP1 | viral latency | Lo et al. 2007 [62] | reporter assay |
| 3 | DICE1 | apoptosis | Kang et al. 2015, Lei et al. 2013 [63,64] | reporter assay, PAR-CLIP |
| 3 | CASZ1a | apoptosis | Kang et al. 2015 [63] | PAR-CLIP, reporter assay |
| 4-5p | BID | apoptosis | Shinozaki-Ushiku et al. 2015 [65] | reporter assay, biopsies |
| 5 | PUMA | apoptosis | Choy et al. 2008 [66] | reporter assay |
| 5-3p | p53 | apoptosis | Zheng et al. 2018 [67] | reporter assay |
| 5-5p | ATM | DNA repair | Lung et al. 2018 [68] | reporter assay |
| 6-3p | LOC353103 RNA | anti-migration | He et al. 2016 [69] | cell culture, reporter assay |
| 6 | OCT1 | apoptosis | Kang et al. 2015 [63] | PAR-CLIP, reporter assay |
| 6 | Dicer | apoptosis | Kang et al. 2015, Iizasa et al. 2010 [63,70] | PAR-CLIP, reporter assay |
| 7-3p | PTEN | migration, signaling | Cai et al. 2015a, Cai et al. 2015b [59,60] | reporter assay |
| 7-3p | ATM | DNA repair | Lung et al. 2018 [68] | reporter assay |
| 8-3p | RNF38 | signaling, migration | Lin et al. 2018 [71] | biopsy sequencing, reporter assay |
| 8 | ARID2 | unknown | Kang et al. 2015 [63] | PAR-CLIP, reporter assay |
| 9 | E CAD | migration | Tsai et al. 2017, Hsu et al. 2014 [24,72] | reporter assay, biopsies |
| 9-3p | ATM | DNA repair | Lung et al. 2018 [68] | reporter assay |
| 10 | BTRC | signaling, migration | Zeng et al. 2014, Yan et al. 2015 [57,73] | reporter assay, biopsies |
| 10-3p | DKK1 | signaling, migration | Min et al. 2019 [74] | reporter assay |
| 11 | trFOXP1 | immune evasion, differentiation | Song et al. 2016 [75] | reporter assay, biopsies |
| 13 | NKIRAS2 | signaling | Xu et al. 2019 [76] | biopsies, cell culture |
| 14-3p | ATM | DNA repair | Lung et al. 2018 [68] | reporter assay |
| 15-3p | BRUCE | anti-apoptosis | Choy et al. 2013 [77] | reporter assay, WB |
| 16 | CRBBP | apoptosis | Kang et al. 2015 [63] | PAR-CLIP, reporter assay |
| 16 | SH2B3 | apoptosis | Kang et al. 2015 [63] | PAR-CLIP, reporter assay |
| 16 | TOMM22 | apoptosis | Kang et al. 2015 [63] | PAR-CLIP, reporter assay |
| 16 | LMP1 | viral latency | Lo et al. 2007 [62] | reporter assay |
| 17-5p | LMP1 | viral latency | Lo et al. 2007 [62] | reporter assay |
| 22 | PAK2 | apoptosis | Kang et al. 2015 [63] | PAR-CLIP, reporter assay |
| 22 | TP53INP1 | apoptosis | Kang et al. 2015 [63] | PAR-CLIP, reporter assay |
| 22 and cluster II | NDRG1 | metastasis, differentiation | Kanda et al. 2015 [78] | microarray reporter assay |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Židovec Lepej, S.; Matulić, M.; Gršković, P.; Pavlica, M.; Radmanić, L.; Korać, P. miRNAs: EBV Mechanism for Escaping Host’s Immune Response and Supporting Tumorigenesis. Pathogens 2020, 9, 353. https://doi.org/10.3390/pathogens9050353
Židovec Lepej S, Matulić M, Gršković P, Pavlica M, Radmanić L, Korać P. miRNAs: EBV Mechanism for Escaping Host’s Immune Response and Supporting Tumorigenesis. Pathogens. 2020; 9(5):353. https://doi.org/10.3390/pathogens9050353
Chicago/Turabian StyleŽidovec Lepej, Snježana, Maja Matulić, Paula Gršković, Mirjana Pavlica, Leona Radmanić, and Petra Korać. 2020. "miRNAs: EBV Mechanism for Escaping Host’s Immune Response and Supporting Tumorigenesis" Pathogens 9, no. 5: 353. https://doi.org/10.3390/pathogens9050353
APA StyleŽidovec Lepej, S., Matulić, M., Gršković, P., Pavlica, M., Radmanić, L., & Korać, P. (2020). miRNAs: EBV Mechanism for Escaping Host’s Immune Response and Supporting Tumorigenesis. Pathogens, 9(5), 353. https://doi.org/10.3390/pathogens9050353