PCR-RFLP Detection and Genogroup Identification of Piscirickettsia salmonis in Field Samples

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Design of a PCR-RFLP Assay to Discriminate between P. salmonis Genogroups
2.2. A T928G Transversion in the 16S rDNA of P. salmonis Enabled Differentiation between Genogroups
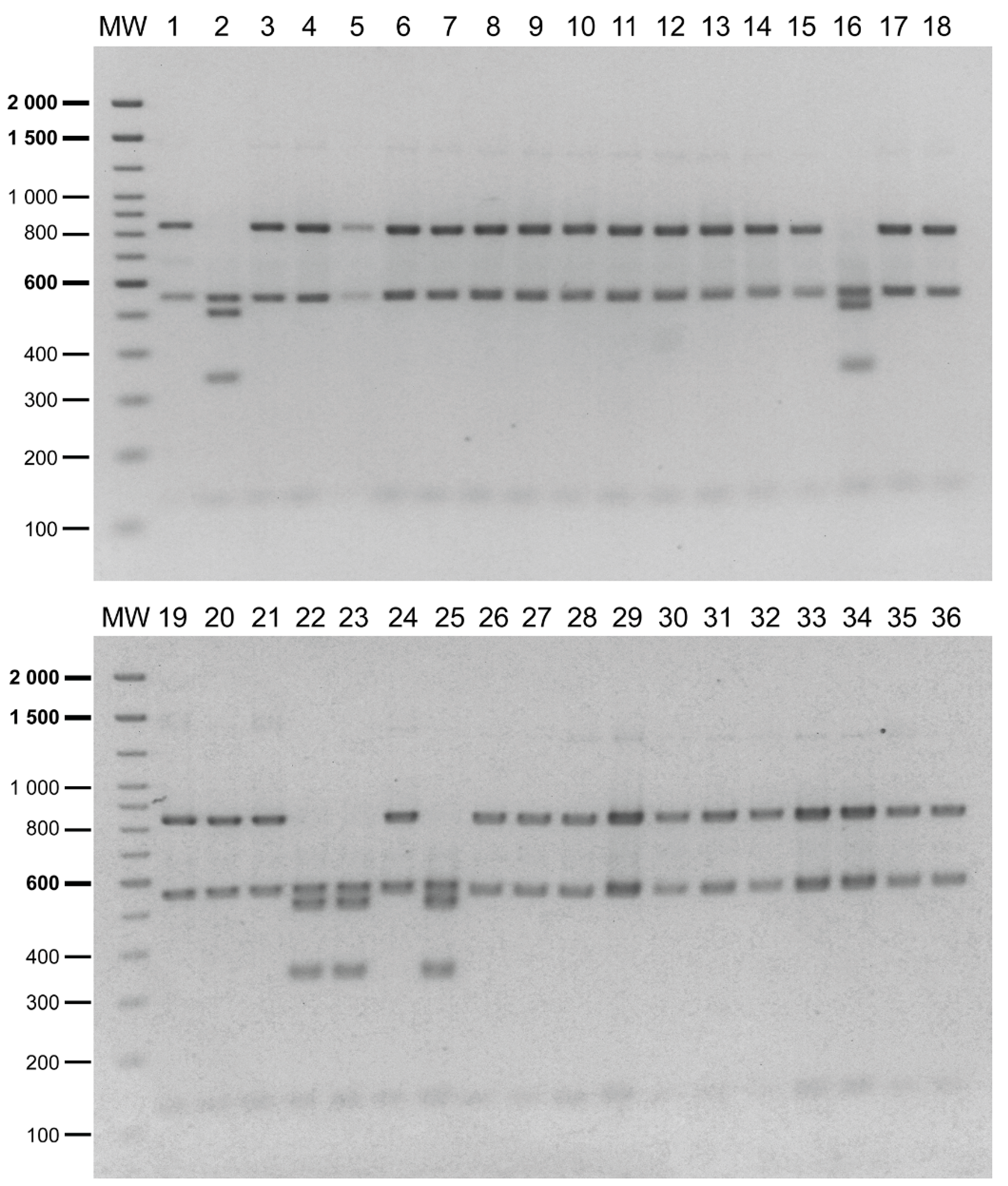
2.3. Detection of P. salmonis Genogroups in Field Samples
2.4. Phylogenetic Analysis of the Field Samples 16S rDNA Sequences
2.5. Bacterial Diversity in Field Samples Assessed by High-Throughput 16S Sequencing
3. Discussion
4. Materials and Methods
4.1. P. salmonis Strains and Growth Conditions
4.2. 16S rDNA Sequences and Restriction Enzymes Bioinformatics Selection
4.3. DNA Extraction from P. salmonis Strains
4.4. Fish Samples Collection, DNA Extraction from Fish Tissues and P. salmonis Detection
4.5. 16S rRNA Gene Amplification and Sequencing
4.6. Phylogenetic Analyses of 16S rRNA Gene
4.7. PCR-RFLP Assays of P. salmonis Strains and Field Samples
4.8. Sequencing of Bacterial 16S rDNA Amplicons
4.9. Analysis of 16S rDNA Amplicon Sequencing
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fryer, J.L.; Hedrick, R.P. Review piscirickettsia salmonis: A Gram-negative intracellular bacterial pathogen of fish. J. Fish. Dis. 2003, 26, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Groff, J.; LaPatra, S. Infectious diseases impacting the commercial culture of salmonids. J. Appl. Aquacult. 2000, 10, 17–90. [Google Scholar] [CrossRef]
- Sernapesca. Informe Sanitario De Salmonicultura En Centros Marinos Año 2018; Sernapesca: Valparaíso, Chile, 2019. [Google Scholar]
- Fryer, J.L.; Lannan, C.N.; Giovannoni, S.J.; Wood, N.D. Piscirickettsia salmonis gen. nov., sp. nov. the causative agent of an epizootic disease in salmonid fishes. Int. J. Syst. Bacteriol. 1992, 42, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Fryer, J.L.; Lannan, C.N.; Giovannoni, S.J.; Wood, N.D. Piscirickettsia salmonis gen. nov., sp. nov., the causative agent of an epizootic disease in salmonid fishes. Fish Pathol. 1990, 25, 107–114. [Google Scholar] [CrossRef]
- Cvitanich, J.D.; Garate, O.N.; Smith, C.E. The isolation of a rickettsia-like organism causing disease and mortality in Chilean salmonids and its confirmation by Koch’ s postulate. J. Fish Dis. 1991, 14, 121–145. [Google Scholar] [CrossRef]
- Branson, E.J.; Nieto Diaz-Munoz, D. Description of a new disease condition occurring in farmed coho salmon, Oncorhynchus kisuteh (Walbaum), in South America. J. Fish. Dis. 1991, 14, 147–157. [Google Scholar] [CrossRef]
- Fryer, J.L.; Lannan, C.N.; Garcés, L.H.; Larenas, J.J.; Smith, P.A. Isolation of a rickettsiales-like organism from diseased coho salmon (Oncorhynchus kisutch) in Chile. Fish. Pathol. 1990, 4, 107–114. [Google Scholar] [CrossRef]
- Brocklebank, J.R.; Speare, D.J.; Armstrong, R.D.; Evelyn, T. British Columbia. Septicemia suspected to be caused by a ricksettia-like agent in farmed Atlantic salmon. Can. Vet. J. 1992, 33, 407–408. [Google Scholar]
- Corbeil, S.; Hyatt, A.D.; Crane, M.S.J. Characterisation of an emerging rickettsia-like organism in Tasmanian farmed Atlantic salmon Salmo salar. Dis. Aquat. Organ. 2005, 64, 37–44. [Google Scholar] [CrossRef]
- Rozas, M.; Enríquez, R. Piscirickettsiosis and Piscirickettsia salmonis in fish: A review. J. Fish. Dis. 2014, 37, 163–188. [Google Scholar] [CrossRef]
- Birrell, J.; Mitchell, S.; Bruno, D.W. Piscirickettsia salmonis in farmed Atlantic salmon, Salmo solar in Scotland. Bull. Eur. Assoc. Fish. Pathol. 2003, 23, 213–218. [Google Scholar]
- Olsen, A.B.; Melby, H.P.; Speilberg, L.; Evensen, Ø.; Håstein, T. Piscirickettsia salmonis infection in Atlantic salmon Salmo salar in Norway-epidemiological, pathological and microbiological findings. Dis. Aquat. Organ. 1997, 31, 35–48. [Google Scholar] [CrossRef] [Green Version]
- Jones, S.R.M.; Markham, F.R.J.; Groman, D.B.; Cusack, R.R. Virulence and antigenic characteristics of a cultured Rickettsiales-like organism isolated from farmed Atlantic salmon Salmo salar in eastern Canada. Dis. Aquat. Organ. 1998, 33, 25–31. [Google Scholar] [CrossRef] [Green Version]
- House, M.; Bartholomew, J.; Winton, J.; Fryer, J. Relative virulence of three isolates of Piscirickettsia salmonis for coho salmon Oncorhynchus Kisutch. Dis. Aquat. Organ. 1992, 35, 107–113. [Google Scholar] [CrossRef]
- Mauel, M.J.; Giovannoni, S.J.; Fryer, J.L. Phylogenetic analysis of Piscirickettsia salmonis by 16S, internal transcribed spacer (ITS) and 23s ribosomal DNA sequencing. Dis. Aquat. Organ. 1999, 35, 115–123. [Google Scholar] [CrossRef]
- Bohle, H.; Henríquez, P.; Grothusen, H.; Navas, E.; Sandoval, A.; Bustamante, F.; Bustos, P.; Mancilla, M. Comparative genome analysis of two isolates of the fish pathogen Piscirickettsia salmonis from different hosts reveals major differences in virulence-associated secretion systems. Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Mandakovic, D.; Glasner, B.; Maldonado, J.; Aravena, P.; González, M.; Cambiazo, V.; Pulgar, R. Genomic-based restriction enzyme selection for specific detection of Piscirickettsia salmonis by 16S rDNA PCR-RFLP. Front. Microbiol. 2016, 7, 643. [Google Scholar] [CrossRef] [Green Version]
- Otterlei, A.; Øyvind, J.; Brevik, D.J.; Duesund, H.; Sommerset, I.; Frost, P.; Mendoza, J.; McKenzie, P.; Nylund, A.; Apablaza, P. Phenotypic and genetic characterization of Piscirickettsia salmonis from Chilean and Canadian salmonids. BMC Vet. Res. 2016, 12, 55. [Google Scholar] [CrossRef] [Green Version]
- Nourdin-Galindo, G.; Sánchez, P.; Molina, F.C.; Espinoza-Rojas, D.A.; Oliver, C.; Ruiz, P.; Vargas-Chacoff, L.; Cárcamo, J.G.; Figueroa, J.E.; Mancilla, M.; et al. Comparative pan-genome analysis of Piscirickettsia salmonis reveals genomic divergences within genogroups. Front. Cell. Infect. Microbiol. 2017, 7, 459. [Google Scholar] [CrossRef] [Green Version]
- Saavedra, J.; Hernandez, N.; Osses, A.; Castillo, A.; Cancino, A.; Grothusen, H.; Navas, E.; Henriquez, P.; Bohle, H.; Bustamante, F.; et al. Prevalence, geographic distribution and phenotypic differences of Piscirickettsia salmonis EM-90-like isolates. J. Fish. Dis. 2017, 2015, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Rozas-Serri, M.; Ildefonso, R.; Peña, A.; Enríquez, R.; Barrientos, S.; Maldonado, L. Comparative pathogenesis of piscirickettsiosis in Atlantic salmon (Salmo salar L.) post-smolt experimentally challenged with LF-89-like and EM-90-like Piscirickettsia salmonis isolates. J. Fish. Dis. 2017, 40, 1451–1472. [Google Scholar] [CrossRef]
- Mauel, M.J.; Ware, C.; Smith, P.A. Culture of Piscirickettsia salmonis on enriched blood agar. J. Vet. Diagn. Investig. 2008, 20, 213–214. [Google Scholar] [CrossRef] [Green Version]
- Mikalsen, J.; Skjaervik, O.; Wiik-Nielsen, J.; Wasmuth, M.A.; Colquhoun, D.J. Agar culture of Piscirickettsia salmonis, a serious pathogen of farmed salmonid and marine fish. FEMS Microbiol. Lett. 2008, 278, 43–47. [Google Scholar] [CrossRef]
- Vera, T.; Isla, A.; Cuevas, A.; Figueroa, J. A new liquid medium for the pathogen Piscirickettsia salmonis T. Arch. Med. Vet. 2012, 44, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Mauel, M.J.; Giovannoni, S.J.; Fryer, J.L. Development of polymerase chain reaction assays for detection, identification, and differentiation of Piscirickettsia salmonis. Dis. Aquat. Org. 1996, 26, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Marshall, S.; Heath, S.; Henríquez, V.; Orrego, C. Minimally invasive detection of Piscirickettsia salmonis in cultivated salmonids via the PCR. Appl. Environ. Microbiol. 1998, 64, 3066–3069. [Google Scholar] [CrossRef] [Green Version]
- Corbeil, S.; McColl, K.A.; Crane, M.S.J. Development of a TaqMan quantitative PCR assay for the identification of Piscirickettsia salmonis. Bull. Eur. Assoc. Fish. Pathol. 2003, 23, 95–101. [Google Scholar]
- Karatas, S.; Mikalsen, J.; Steinum, T.M.; Taksdal, T.; Bordevik, M.; Colquhoun, D.J. Real time PCR detection of Piscirickettsia salmonis from formalin-fixed paraffin-embedded tissues. J. Fish Dis. 2008, 31, 747–753. [Google Scholar] [CrossRef]
- Lannan, C.; Ewing, S.; Fryer, J. A fluorescent antibody test for detection of the rickettsia causing disease in chilean salmonids. J. Aquat. Anim. Health 1991, 3, 124–129. [Google Scholar] [CrossRef]
- Reiling, N.; Homolka, S.; Walter, K.; Brandenburg, J.; Niwinski, L.; Ernst, M.; Herzmann, C.; Lange, C.; Diel, R.; Ehlers, S.; et al. Clade-specific virulence patterns of mycobacterium tuberculosis complex strains in human primary macrophages and aerogenically infected mice. MBio 2013, 4, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sousa, P.S.; Silva, I.N.; Moreira, L.M.; Veríssimo, A.; Costa, J. Differences in virulence between Legionella pneumophila isolates from human and non-human sources determined in Galleria mellonella infection model. Front. Cell. Infect. Microbiol. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Isla, A.; Saldarriaga-Córdoba, M.; Fuentes, D.E.; Albornoz, R.; Haussmann, D.; Mancilla-Schulz, J.; Martínez, A.; Figueroa, J.; Avendaño-Herrera, R.; Yáñe, A. Multilocus sequence typing detects new Piscirickettsia salmonis hybrid genogroup in Chilean fish farms: Evidence for genetic diversity and population structure. J. Fish. Dis. 2019, 42, 721–737. [Google Scholar] [CrossRef] [PubMed]
- Hallin, M.; Deplano, A.; Struelens, M.J. Molecular typing of bacterial pathogens: A tool for the epidemiological study and control of infectious diseases. In New Frontiers of Molecular Epidemiology of Infectious Diseases; Springer: Dordrecht, The Netherlands, 2012; pp. 9–25. [Google Scholar]
- Karlsen, C.; Vanberg, C.; Mikkelsen, H.; Sørum, H. Co-infection of Atlantic salmon (Salmo salar), by Moritella viscosa and Aliivibrio wodanis, development of disease and host colonization. Vet. Microbiol. 2014, 171, 112–121. [Google Scholar] [CrossRef]
- Loch, T.P.; Scribner, K.; Tempelman, R.; Whelan, G.; Faisal, M. Bacterial infections of Chinook salmon, Oncorhynchus tshawytscha (Walbaum), returning to gamete collecting weirs in Michigan. J. Fish. Dis. 2012, 35, 39–50. [Google Scholar] [CrossRef]
- Zarkasi, K.Z.; Abell, G.C.J.; Taylor, R.S.; Neuman, C.; Hatje, E.; Tamplin, M.L.; Katouli, M.; Bowman, J.P. Pyrosequencing-based characterization of gastrointestinal bacteria of Atlantic salmon (Salmo salar L.) within a commercial mariculture system. J. Appl. Microbiol. 2014, 117, 18–27. [Google Scholar] [CrossRef] [Green Version]
- De Bruijn, I.; Liu, Y.; Wiegertjes, G.F.; Raaijmakers, J.M. Exploring fish microbial communities to mitigate emerging diseases in aquaculture. FEMS Microbiol. Ecol. 2018, 94, 1–12. [Google Scholar] [CrossRef]
- Larsen, A.; Tao, Z.; Bullard, S.A.; Arias, C.R. Diversity of the skin microbiota of fishes: Evidence for host species specificity. FEMS Microbiol. Ecol. 2013, 85, 483–494. [Google Scholar] [CrossRef]
- Galbraith, H.; Iwanowicz, D.; Spooner, D.; Iwanowicz, L.; Keller, D.; Zelanko, P.; Adams, C. Exposure to synthetic hydraulic fracturing waste influences the mucosal bacterial community structure of the brook trout (Salvelinus fontinalis) epidermis. AIMS Microbiol. 2018, 4, 413–427. [Google Scholar] [CrossRef]
- Navarrete, P.; Espejo, R.T.; Romero, J. Molecular analysis of microbiota along the digestive tract of juvenile atlantic salmon (Salmo salar L.). Microb. Ecol. 2009, 57, 550–561. [Google Scholar] [CrossRef]
- Zhou, Z.; Liu, Y.; Shi, P.; He, S.; Yao, B.; Ringø, E. Molecular characterization of the autochthonous microbiota in the gastrointestinal tract of adult yellow grouper (Epinephelus awoara) cultured in cages. Aquaculture 2009, 286, 184–189. [Google Scholar] [CrossRef]
- Egerton, S.; Culloty, S.; Whooley, J.; Stanton, C.; Ross, R.P. The gut microbiota of marine fish. Front. Microbiol. 2018, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Miranda, C.; Palomares, E.; Jurado, M.; Marín, A.; Vega, F.; Soriano-Vargas, E. Isolation and distribution of bacterial flora in farmed rainbow trout from Mexico. J. Aquat. Anim. Health 2010, 22, 244–247. [Google Scholar] [CrossRef]
- Sevellec, M.; Pavey, S.A.; Boutin, S.; Filteau, M.; Derome, N.; Bernatchez, L. Microbiome investigation in the ecological speciation context of lake whitefish (Coregonus clupeaformis) using next-generation sequencing. J. Evol. Biol. 2014, 27, 1029–1046. [Google Scholar] [CrossRef]
- Gomez-Gil, B.; Fajer-Avila, E.; García-Vargas, F. Vibrios of the spotted rose snapper Lutjanus guttatus Steindachner, 1869 from northwestern Mexico. J. Appl. Microbiol. 2007, 102, 1518–1526. [Google Scholar] [CrossRef]
- Meron, D.; Davidovich, N.; Ofek-Lalzar, M.; Berzak, R.; Scheinin, A.; Regev, Y.; Diga, R.; Tchernov, D.; Morick, D. Specific pathogens and microbial abundance within liver and kidney tissues of wild marine fish from the eastern Mediterranean sea. Microb. Biotechnol. 2020, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Tort, L. Stress and immune modulation in fish. Dev. Comp. Immunol. 2011, 35, 1366–1375. [Google Scholar] [CrossRef]
- Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2016, 44, D67–D72. [Google Scholar] [CrossRef] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.R.; Wang, Q.E.; Cardenas, J.; Fish, B.; Chai, R.J.; Farris, A.S.; Kulam-Syed-Mohideen, D.M.; McGarrell, T.; Marsh, G.; Garrity, M.; et al. The Ribosomal database project: Improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef] [Green Version]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Dong, H.; Zhang, G.; Yu, B.; Chapman, L.R.; Fields, M.W. Microbial diversity in water and sediment of lake Chaka, an athalassohaline lake in northwestern China. Appl. Environ. Microbiol. 2006, 72, 3832–3845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, R.J.; Vincze, T.; Posfai, J.; Macelis, D. REBASE—A database for DNA restriction and modification: Enzymes, genes and genomes. Nucleic Acids Res. 2015, 43, D298–D299. [Google Scholar] [CrossRef] [PubMed]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Sumpos. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.; Pryer, K.M.; Miao, V.P.; Palmer, J.D. Investigating deep phylogenetic relationships among cyanobacteria and plastids by small subunit rRNA sequence analysis. J. Eukaryot. Microbiol. 1999, 46, 327–338. [Google Scholar] [CrossRef]
- Mandakovic, D.; Maldonado, J.; Pulgar, R.; Cabrera, P.; Gaete, A.; Urtuvia, V.; Seeger, M.; Cambiazo, V.; González, M. Microbiome analysis and bacterial isolation from Lejía lake soil in Atacama desert. Extremophiles 2018, 22, 665–673. [Google Scholar] [CrossRef]
- Dowd, S.E.; Callaway, T.R.; Wolcott, R.D.; Sun, Y.; McKeehan, T.; Hagevoort, R.G.; Edrington, T.S. Evaluation of the bacterial diversity in the feces of cattle using 16S rDNA bacterial tag-encoded FLX amplicon pyrosequencing (bTEFAP). BMC Microbiol. 2008, 8, 125. [Google Scholar] [CrossRef] [Green Version]
- Handl, S.; Dowd, S.E.; Garcia-Mazcorro, J.F.; Steiner, J.M.; Suchodolski, J.S. Massive parallel 16S rRNA gene pyrosequencing reveals highly diverse fecal bacterial and fungal communities in healthy dogs and cats. FEMS Microbiol. Ecol. 2011, 76, 301–310. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aravena, P.; Pulgar, R.; Ortiz-Severín, J.; Maza, F.; Gaete, A.; Martínez, S.; Serón, E.; González, M.; Cambiazo, V. PCR-RFLP Detection and Genogroup Identification of Piscirickettsia salmonis in Field Samples. Pathogens 2020, 9, 358. https://doi.org/10.3390/pathogens9050358
Aravena P, Pulgar R, Ortiz-Severín J, Maza F, Gaete A, Martínez S, Serón E, González M, Cambiazo V. PCR-RFLP Detection and Genogroup Identification of Piscirickettsia salmonis in Field Samples. Pathogens. 2020; 9(5):358. https://doi.org/10.3390/pathogens9050358
Chicago/Turabian StyleAravena, Pamela, Rodrigo Pulgar, Javiera Ortiz-Severín, Felipe Maza, Alexis Gaete, Sebastián Martínez, Ervin Serón, Mauricio González, and Verónica Cambiazo. 2020. "PCR-RFLP Detection and Genogroup Identification of Piscirickettsia salmonis in Field Samples" Pathogens 9, no. 5: 358. https://doi.org/10.3390/pathogens9050358