MLVA-16 Genotyping of Brucella abortus and Brucella melitensis Isolates from Different Animal Species in Egypt: Geographical Relatedness and the Mediterranean Lineage

 ,
,  , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
2.1. Biotyping and Origin of B. melitensis and B. abortus Isolates
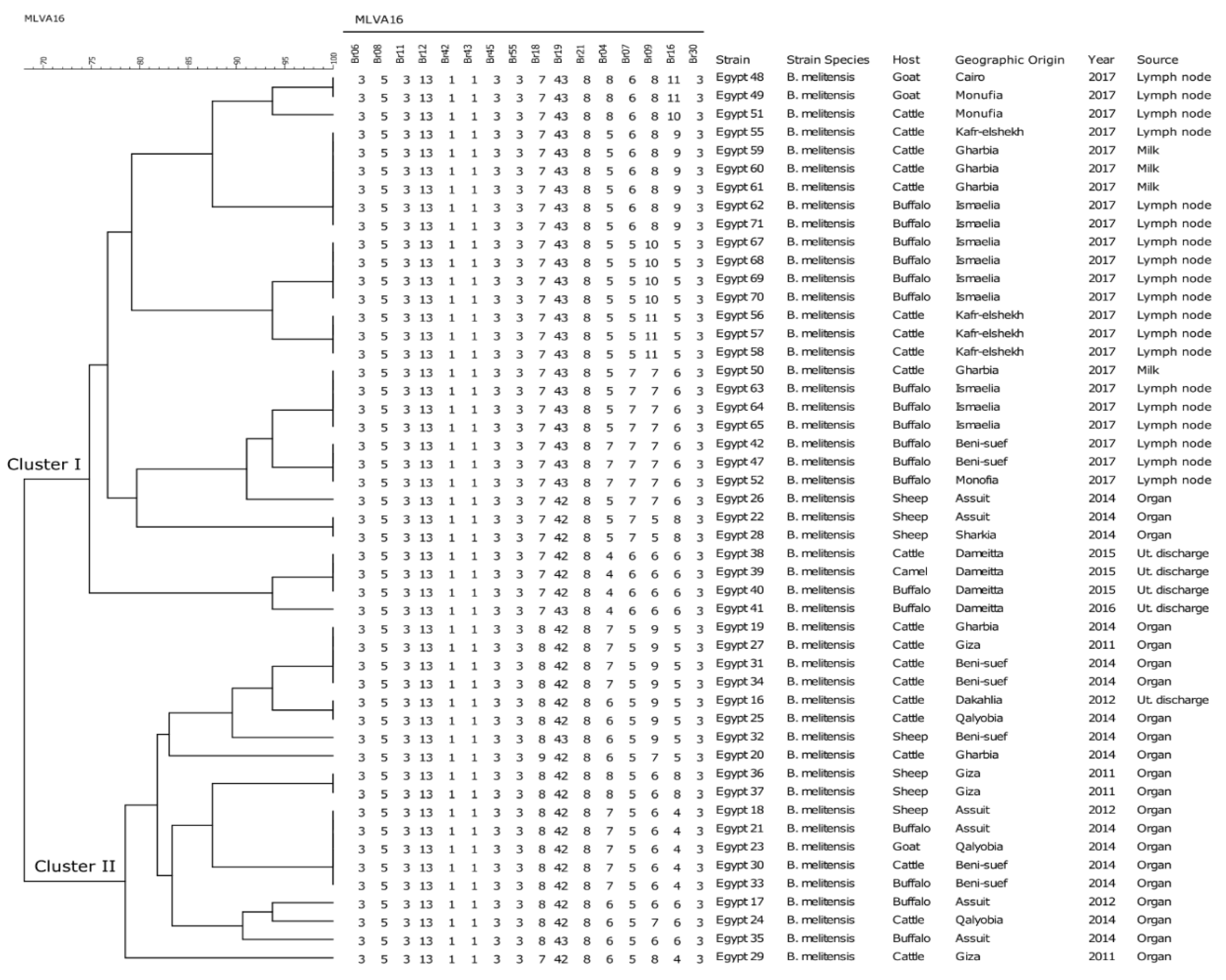
2.2. Clustering, Diversity, and Distribution of B. melitensis
2.3. Clustering, Diversity, and Distribution of B. abortus
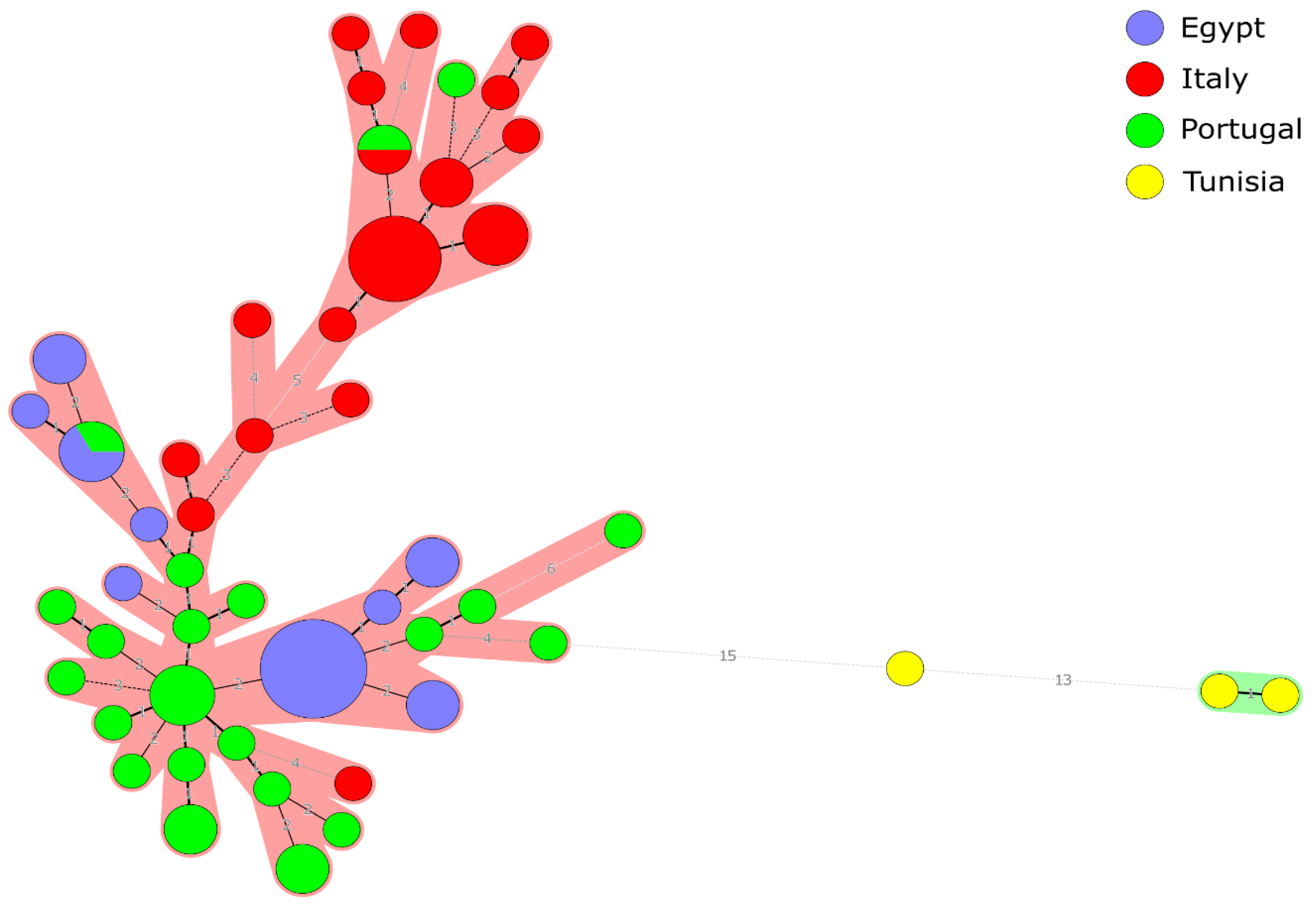
2.4. Geographical Relatedness and the “Mediterranean Lineage”
3. Discussion
4. Materials and Methods
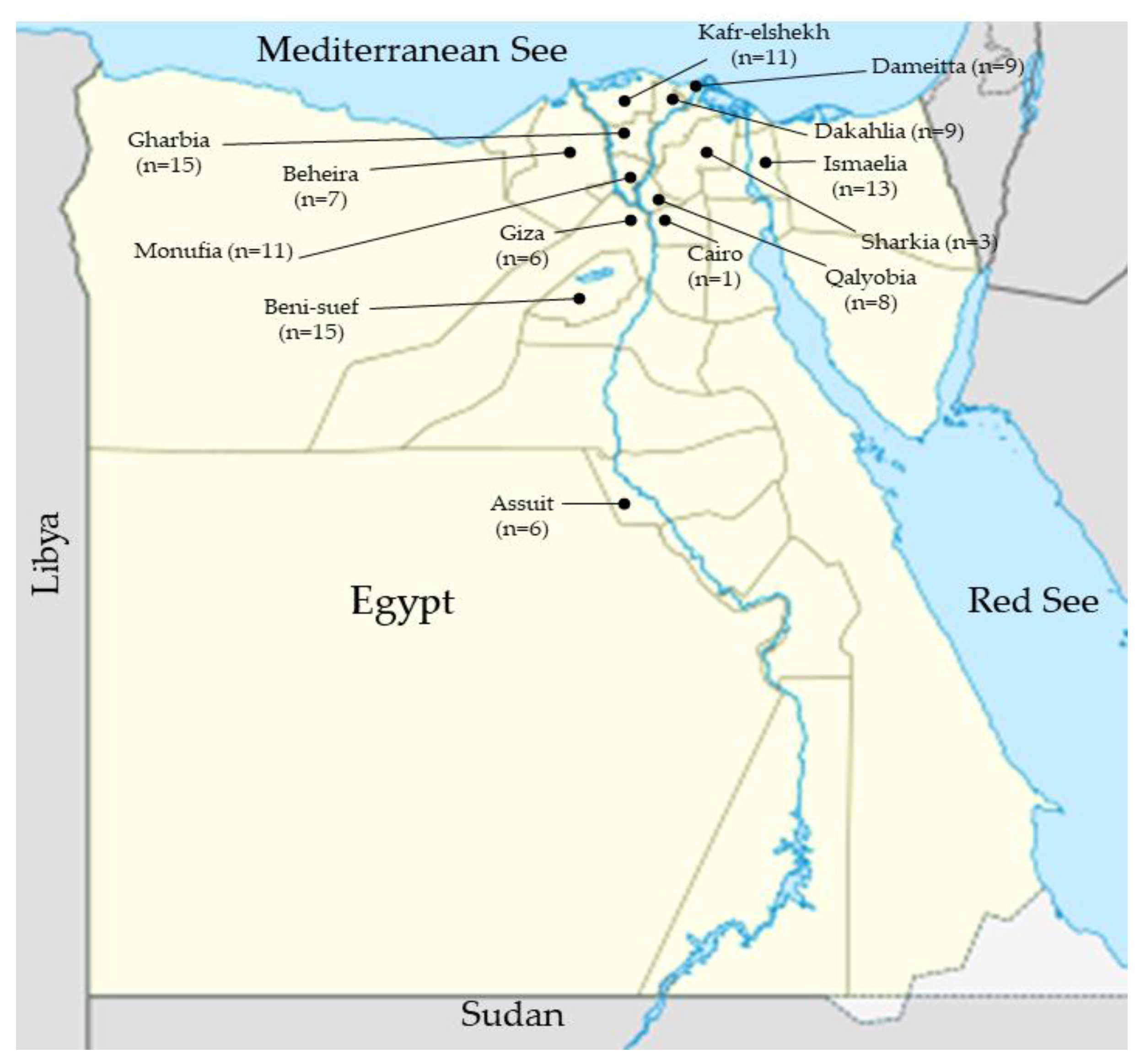
4.1. Brucella Strains
4.2. Identification and Biotyping of Isolates
4.3. Genetic Diversity of Brucella Strains Analyzed MLVA-16
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cutler, S.J.; Whatmore, A.M.; Commander, N.J. Brucellosis—New aspects of an old disease. J. Appl. Microbiol. 2005, 98, 1270–1281. [Google Scholar] [CrossRef]
- Godfroid, J.; Scholz, H.C.; Barbier, T.; Nicolas, C.; Wattiau, P.; Fretin, D.; Whatmore, A.M.; Cloeckaert, A.; Blasco, J.M.; Moriyon, I.; et al. Brucellosis at the animal/ecosystem/human interface at the beginning of the 21st century. Prev. Vet. Med. 2011, 102, 118–131. [Google Scholar] [CrossRef] [Green Version]
- Chiliveru, S.; Appari, M.; Suravajhala, P. On Brucella pathogenesis: Looking for the unified challenge in systems and synthetic biology. Syst. Synth. Biol. 2015, 9, 73–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dentinger, C.M.; Jacob, K.; Lee, L.V.; Mendez, H.A.; Chotikanatis, K.; McDonough, P.L.; Chico, D.M.; De, B.K.; Tiller, R.V.; Traxler, R.M.; et al. Human Brucella canis infection and subsequent laboratory exposures associated with a puppy, New York City, 2012. Zoonoses Public Health 2014. [Google Scholar] [CrossRef]
- Galińska, E.; Zagórski, J. Brucellosis in humans—Etiology, diagnostics, clinical forms. Ann. Agric. Environ. Med. 2013, 20, 233–238. [Google Scholar] [PubMed]
- Hashemifar, I.; Yadegar, A.; Jazi, F.M.; Amirmozafari, N. Molecular prevalence of putative virulence-associated genes in Brucella melitensis and Brucella abortus isolates from human and livestock specimens in Iran. Microb. Pathog. 2017, 105, 334–339. [Google Scholar] [CrossRef]
- De Massis, F.; Zilli, K.; Di Donato, G.; Nuvoloni, R.; Pelini, S.; Sacchini, L.; D’Alterio, N.; Di Giannatale, E. Distribution of Brucella field strains isolated from livestock, wildlife populations, and humans in Italy from 2007 to 2015. PLoS ONE 2019, 14, e0213689. [Google Scholar] [CrossRef]
- El-Tras, W.F.; Tayel, A.A.; Eltholth, M.M.; Guitian, J. Brucella infection in fresh water fish: Evidence for natural infection of Nile catfish, Clarias gariepinus, with Brucella melitensis. Vet. Microbiol. 2010, 141, 321–325. [Google Scholar] [CrossRef]
- O’Brien, M.P.; Beja-Pereira, A.; Anderson, N.; Ceballos, R.M.; Edwards, W.H.; Harris, B.; Wallen, R.L.; Costa, V. Brucellosis transmission between wildlife and livestock in the greater yellowstone ecosystem: Inferences from DNA genotyping. J. Wildl. Dis. 2017, 53, 339–343. [Google Scholar] [CrossRef]
- Wareth, G.; Melzer, F.; El-Diasty, M.; Schmoock, G.; Elbauomy, E.; Abdel-Hamid, N.; Sayour, A.; Neubauer, H. Isolation of Brucella abortus from a dog and a cat confirms their biological role in re-emergence and dissemination of bovine brucellosis on dairy farms. Transbound. Emerg. Dis. 2017, 64, e27–e30. [Google Scholar] [CrossRef]
- Pappas, G.; Papadimitriou, P.; Akritidis, N.; Christou, L.; Tsianos, E.V. The new global map of human brucellosis. Lancet Infect. Dis. 2006, 6, 91–99. [Google Scholar] [CrossRef]
- Wareth, G.; Abdeen, A.; Benyounes, A.; Ferreira, A.C.; Arla Juma, A.K.; Krt, B.; Garin-Bastuji, B.; Kornspan, D.; Laušević, D.; Melzer, F.; et al. Brucellosis in the Mediterranean Countries: History, Prevalence, Distribution, Current Situation and Attempts at Surveillance and Control; Office Int’l des Epizooties (OIE): Paris, France, 2019; Volume 12, p. 98. [Google Scholar]
- Greco, E.; El-Aguizy, O.; Ali, M.F.; Foti, S.; Cunsolo, V.; Saletti, R.; Ciliberto, E. Proteomic analyses on an ancient egyptian cheese and biomolecular evidence of brucellosis. Anal. Chem. 2018, 90, 9673–9676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capasso, L. Bacteria in two-millennia-old cheese, and related epizoonoses in Roman populations. J. Infect. 2002, 45, 122–127. [Google Scholar] [CrossRef]
- Mutolo, M.J.; Jenny, L.L.; Buszek, A.R.; Fenton, T.W.; Foran, D.R. Osteological and molecular identification of brucellosis in ancient Butrint, Albania. Am. J. Phys. Anthropol. 2012, 147, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Kay, G.L.; Sergeant, M.J.; Giuffra, V.; Bandiera, P.; Milanese, M.; Bramanti, B.; Bianucci, R.; Pallen, M.J. Recovery of a medieval Brucella melitensis genome using shotgun metagenomics. MBio 2014, 5, e01337-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Refai, M. Incidence and control of brucellosis in the Near East region. Vet. Microbiol. 2002, 90, 81–110. [Google Scholar] [CrossRef]
- Wareth, G.; Hikal, A.; Refai, M.; Melzer, F.; Roesler, U.; Neubauer, H. Animal brucellosis in Egypt. J. Infect. Dev. Ctries. 2014, 8, 1365–1373. [Google Scholar] [CrossRef] [PubMed]
- Samaha, H.; Al-Rowaily, M.; Khoudair, R.M.; Ashour, H.M. Multicenter study of brucellosis in Egypt. Emerg. Infect. Dis. 2008, 14, 1916–1918. [Google Scholar] [CrossRef]
- Hamdy, M.E.; Amin, A.S. Detection of Brucella species in the milk of infected cattle, sheep, goats and camels by PCR. Vet. J. (Lond. Engl. 1997) 2002, 163, 299–305. [Google Scholar] [CrossRef]
- Wareth, G.; Melzer, F.; Elschner, M.C.; Neubauer, H.; Roesler, U. Detection of Brucella melitensis in bovine milk and milk products from apparently healthy animals in Egypt by real-time PCR. J. Infect. Dev. Ctries. 2014, 8, 1339–1343. [Google Scholar] [CrossRef] [Green Version]
- Al Dahouk, S.; Fleche, P.L.; Nockler, K.; Jacques, I.; Grayon, M.; Scholz, H.C.; Tomaso, H.; Vergnaud, G.; Neubauer, H. Evaluation of Brucella MLVA typing for human brucellosis. J. Microbiol. Methods 2007, 69, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Maquart, M.; Le Fleche, P.; Foster, G.; Tryland, M.; Ramisse, F.; Djonne, B.; Al Dahouk, S.; Jacques, I.; Neubauer, H.; Walravens, K.; et al. MLVA-16 typing of 295 marine mammal Brucella isolates from different animal and geographic origins identifies 7 major groups within Brucella ceti and Brucella Pinnipedialis. BMC Microbiol. 2009, 9, 145. [Google Scholar] [CrossRef] [PubMed]
- Dorneles, E.M.; De Faria, A.P.; Pauletti, R.B.; Santana, J.A.; Caldeira, G.A.; Heinemann, M.B.; Titze-de-Almeida, R.; Lage, A.P. Genetic stability of Brucella abortus S19 and RB51 vaccine strains by multiple locus variable number tandem repeat analysis (MLVA16). Vaccine 2013, 31, 4856–4859. [Google Scholar] [CrossRef] [PubMed]
- Tiller, R.V.; De, B.K.; Boshra, M.; Huynh, L.Y.; Van Ert, M.N.; Wagner, D.M.; Klena, J.; Mohsen, T.S.; El-Shafie, S.S.; Keim, P.; et al. Comparison of two multiple-locus variable-number tandem-repeat analysis methods for molecular strain typing of human Brucella melitensis isolates from the Middle East. J. Clin. Microbiol. 2009, 47, 2226–2231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menshawy, A.M.; Perez-Sancho, M.; Garcia-Seco, T.; Hosein, H.I.; Garcia, N.; Martinez, I.; Sayour, A.E.; Goyache, J.; Azzam, R.A.; Dominguez, L.; et al. Assessment of genetic diversity of zoonotic Brucella spp. recovered from livestock in Egypt using multiple locus VNTR analysis. BioMed Res. Int. 2014, 2014, 353876. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Hamid, N.H.; El-Bauomy, E.M.; Ghobashy, H.M.; Shehata, A.A. Genetic variation of Brucella isolates at strain level in Egypt. Vet. Med. Sci. 2020. [Google Scholar] [CrossRef] [Green Version]
- Sayour, A.E.; Elbauomy, E.; Abdel-Hamid, N.H.; Mahrous, A.; Carychao, D.; Cooley, M.B.; Elhadidy, M. MLVA fingerprinting of Brucella melitensis circulating among livestock and cases of sporadic human illness in Egypt. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Lounes, N.; Cherfa, M.A.; Le Carrou, G.; Bouyoucef, A.; Jay, M.; Garin-Bastuji, B.; Mick, V. Human brucellosis in Maghreb: Existence of a lineage related to socio-historical connections with Europe. PLoS ONE 2014, 9, e115319. [Google Scholar] [CrossRef]
- Dadar, M.; Shahali, Y.; Wareth, G. Molecular diagnosis of acute and chronic brucellosis in humans. In Microbial Technology for the Welfare of Society. Microorganisms for Sustainability; Arora, P., Ed.; Springer: Singapore, 2019; Volume 17, p. 22. [Google Scholar]
- Ferreira, A.C.; Chambel, L.; Tenreiro, T.; Cardoso, R.; Flor, L.; Dias, I.T.; Pacheco, T.; Garin-Bastuji, B.; Le Flèche, P.; Vergnaud, G.; et al. MLVA16 typing of Portuguese human and animal Brucella melitensis and Brucella abortus isolates. PLoS ONE 2012, 7, e42514. [Google Scholar] [CrossRef] [Green Version]
- Kattar, M.M.; Jaafar, R.F.; Araj, G.F.; Le Flèche, P.; Matar, G.M.; Abi Rached, R.; Khalife, S.; Vergnaud, G. Evaluation of a multilocus variable-number tandem-repeat analysis scheme for typing human Brucella isolates in a region of brucellosis endemicity. J. Clin. Microbiol. 2008, 46, 3935–3940. [Google Scholar] [CrossRef] [Green Version]
- Sacchini, L.; Wahab, T.; Di Giannatale, E.; Zilli, K.; Abass, A.; Garofolo, G.; Janowicz, A. Whole genome sequencing for tracing geographical origin of imported cases of human brucellosis in Sweden. Microorganisms 2019, 7, 398. [Google Scholar] [CrossRef] [Green Version]
- Janowicz, A.; De Massis, F.; Ancora, M.; Cammà, C.; Patavino, C.; Battisti, A.; Prior, K.; Harmsen, D.; Scholz, H.; Zilli, K.; et al. Core genome multilocus sequence typing and single nucleotide polymorphism analysis in the epidemiology of Brucella melitensis infections. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgi, E.; Walter, M.C.; Pfalzgraf, M.T.; Northoff, B.H.; Holdt, L.M.; Scholz, H.C.; Zoeller, L.; Zange, S.; Antwerpen, M.H. Whole genome sequencing of Brucella melitensis isolated from 57 patients in Germany reveals high diversity in strains from Middle East. PLoS ONE 2017, 12, e0175425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karger, A.; Melzer, F.; Timke, M.; Bettin, B.; Kostrzewa, M.; Nockler, K.; Hohmann, A.; Tomaso, H.; Neubauer, H.; Al Dahouk, S. Interlaboratory comparison of intact-cell matrix-assisted laser desorption ionization-time of flight mass spectrometry results for identification and differentiation of Brucella spp. J. Clin. Microbiol. 2013, 51, 3123–3126. [Google Scholar] [CrossRef] [Green Version]
- Murugaiyan, J.; Walther, B.; Stamm, I.; Abou-Elnaga, Y.; Brueggemann-Schwarze, S.; Vincze, S.; Wieler, L.H.; Lubke-Becker, A.; Semmler, T.; Roesler, U. Species differentiation within the Staphylococcus intermedius group using a refined MALDI-TOF MS database. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2014, 20, 1007–1015. [Google Scholar] [CrossRef] [Green Version]
- Alton, G.G.; Jones, L.M.; Angus, R.D.; Verger, J.M. Techniques for the Brucellosis Laboratory, 17–62; Instituttional de la Recherche Agronomique: Paris, France, 1988. [Google Scholar]
- Bricker, B.J.; Halling, S.M. Differentiation of Brucella abortus bv. 1, 2, and 4, Brucella melitensis, Brucella ovis, and Brucella suis bv. 1 by PCR. J. Clin. Microbiol. 1994, 32, 2660–2666. [Google Scholar] [CrossRef] [Green Version]
- Le Fleche, P.; Jacques, I.; Grayon, M.; Al Dahouk, S.; Bouchon, P.; Denoeud, F.; Nockler, K.; Neubauer, H.; Guilloteau, L.A.; Vergnaud, G. Evaluation and selection of tandem repeat loci for a Brucella MLVA typing assay. BMC Microbiol. 2006, 6, 9. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host | Cattle | Buffalo | Sheep | Goat | Camel | Dog | Cat | Vaccine | Total |
|---|---|---|---|---|---|---|---|---|---|
| B. melitensis | 34 | 27 | 14 | 9 | 1 | 0 | 0 | 0 | 85 |
| B. abortus | 18 | 2 | 0 | 0 | 0 | 1 | 1 | 2 | 24 |
| Number of strains | 52 | 29 | 14 | 9 | 1 | 1 | 1 | 2 | 109 |
| No. | Geographical Area | Governorates | Number of Isolates |
|---|---|---|---|
| 1 | Capital cities governorates | Giza | 6 (1A, 5M) |
| Cairo | 1 (1M) | ||
| 2 | The North coastal governorates | Dakahlia | 9 (8A, 1M) |
| Dameitta | 9 (5A, 4M) | ||
| 3 | The Northeastern part of the country | Ismaelia | 13 (1A, 12M) |
| 4 | Upper Egypt governorates | Assuit | 6 (6M) |
| Beni-suef | 15 (3A, 12M) | ||
| 5 | Delta region governorates | Beheira | 7 (6A, 1M) |
| Gharbia | 15 (15M) | ||
| Monufia | 11 (11M) | ||
| Qalyobia | 8 (8M) | ||
| Sharkia | 3 (3M) | ||
| Kafr-elshekh | 6 (6M) | ||
| Total number | 13 | 109 (24A, 85M) | |
| Country | Brucella spp. | Host | Years of Isolation | No. of Isolates |
|---|---|---|---|---|
| Egypt (n = 69) | B. abortus | Cattle, buffalo, dog, cat, RB51 vaccine strain | 2011–2017 | 20 |
| B. melitensis | Cattle, buffalo, sheep, goat, camel | 2011–2017 | 49 | |
| Italy (n = 49) | B. abortus | Cattle, buffalo | 2011–2015 | 25 |
| B. melitensis | Sheep, goat, bovine, humans, ibex | 2011–2016 | 24 | |
| Portugal (n = 52) | B. abortus | Cattle | 2001–2007 | 26 |
| B. melitensis | Sheep, goat, cattle, human | 2001–2010 | 26 | |
| Greece (n = 63) | B. abortus | - | - | 0 |
| B. melitensis | Human, small ruminants | ND * | 63 | |
| Tunisia (n = 4) | B. abortus | Cattle | 2018 | 3 |
| B. melitensis | Sheep | 2017 | 1 | |
| Total | 2001–2017 | 237 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wareth, G.; El-Diasty, M.; Melzer, F.; Schmoock, G.; Moustafa, S.A.; El-Beskawy, M.; Khater, D.F.; Hamdy, M.E.R.; Zaki, H.M.; Ferreira, A.C.; et al. MLVA-16 Genotyping of Brucella abortus and Brucella melitensis Isolates from Different Animal Species in Egypt: Geographical Relatedness and the Mediterranean Lineage. Pathogens 2020, 9, 498. https://doi.org/10.3390/pathogens9060498
Wareth G, El-Diasty M, Melzer F, Schmoock G, Moustafa SA, El-Beskawy M, Khater DF, Hamdy MER, Zaki HM, Ferreira AC, et al. MLVA-16 Genotyping of Brucella abortus and Brucella melitensis Isolates from Different Animal Species in Egypt: Geographical Relatedness and the Mediterranean Lineage. Pathogens. 2020; 9(6):498. https://doi.org/10.3390/pathogens9060498
Chicago/Turabian StyleWareth, Gamal, Mohamed El-Diasty, Falk Melzer, Gernot Schmoock, Shawky A. Moustafa, Mohamed El-Beskawy, Dali F. Khater, Mahmoud E.R. Hamdy, Hoda M. Zaki, Ana Cristina Ferreira, and et al. 2020. "MLVA-16 Genotyping of Brucella abortus and Brucella melitensis Isolates from Different Animal Species in Egypt: Geographical Relatedness and the Mediterranean Lineage" Pathogens 9, no. 6: 498. https://doi.org/10.3390/pathogens9060498
APA StyleWareth, G., El-Diasty, M., Melzer, F., Schmoock, G., Moustafa, S. A., El-Beskawy, M., Khater, D. F., Hamdy, M. E. R., Zaki, H. M., Ferreira, A. C., Ekateriniadou, L. V., Boukouvala, E., Abdel-Glil, M. Y., Menshawy, A. M. S., Sancho, M. P., Sakhria, S., Pletz, M. W., & Neubauer, H. (2020). MLVA-16 Genotyping of Brucella abortus and Brucella melitensis Isolates from Different Animal Species in Egypt: Geographical Relatedness and the Mediterranean Lineage. Pathogens, 9(6), 498. https://doi.org/10.3390/pathogens9060498