Seasonal Filarial Infections and Their Black Fly Vectors in Chiang Mai Province, Northern Thailand


 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Black Fly Species Collected and Seasonal Abundance
2.1.1. Ban Lek (BL)
2.1.2. Ban Pang Dang (BPD)
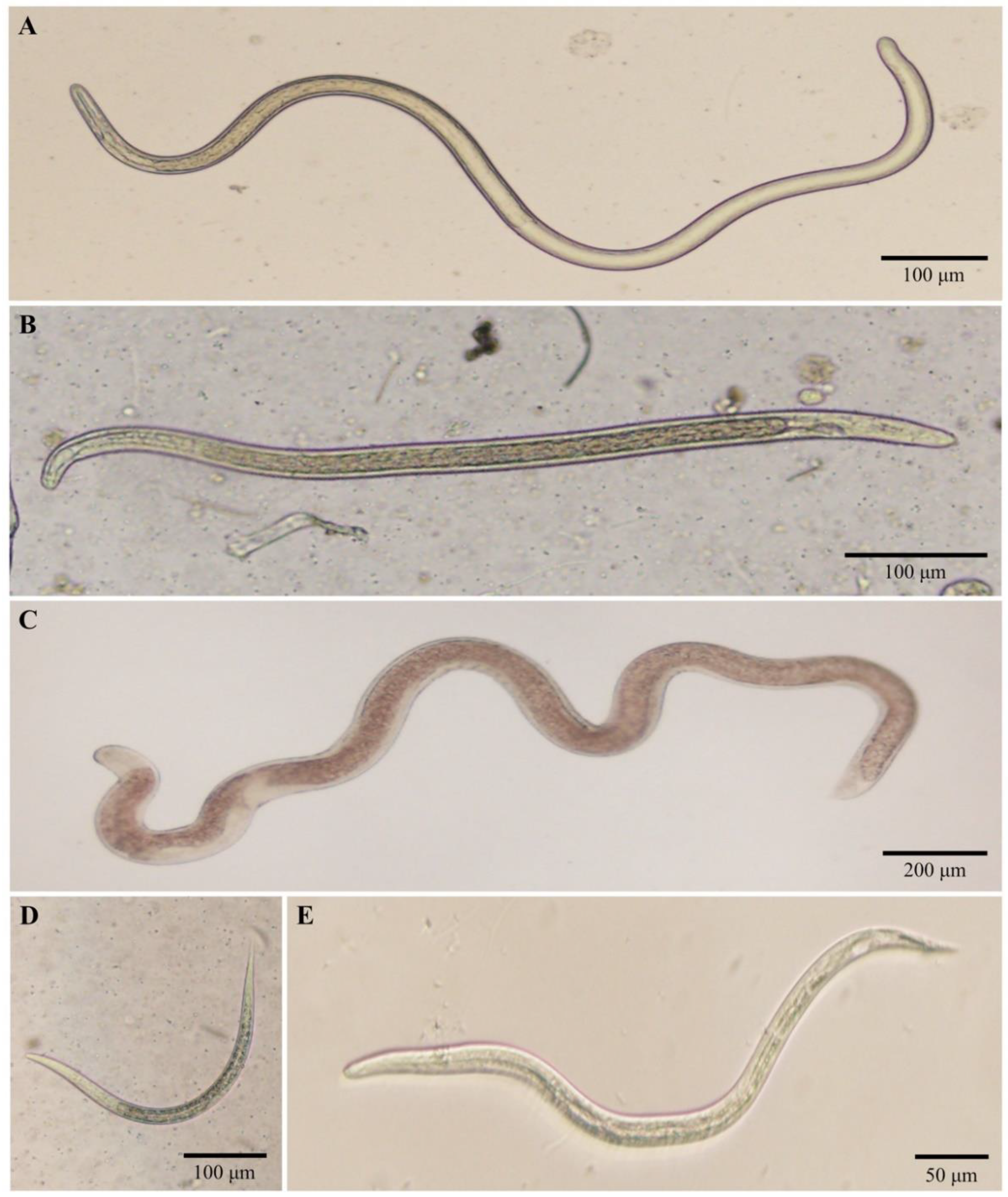
2.2. Natural Filarial Infections and Morphological Identification of Recovered Larvae
2.2.1. Ban Lek
2.2.2. Ban Pang Dang
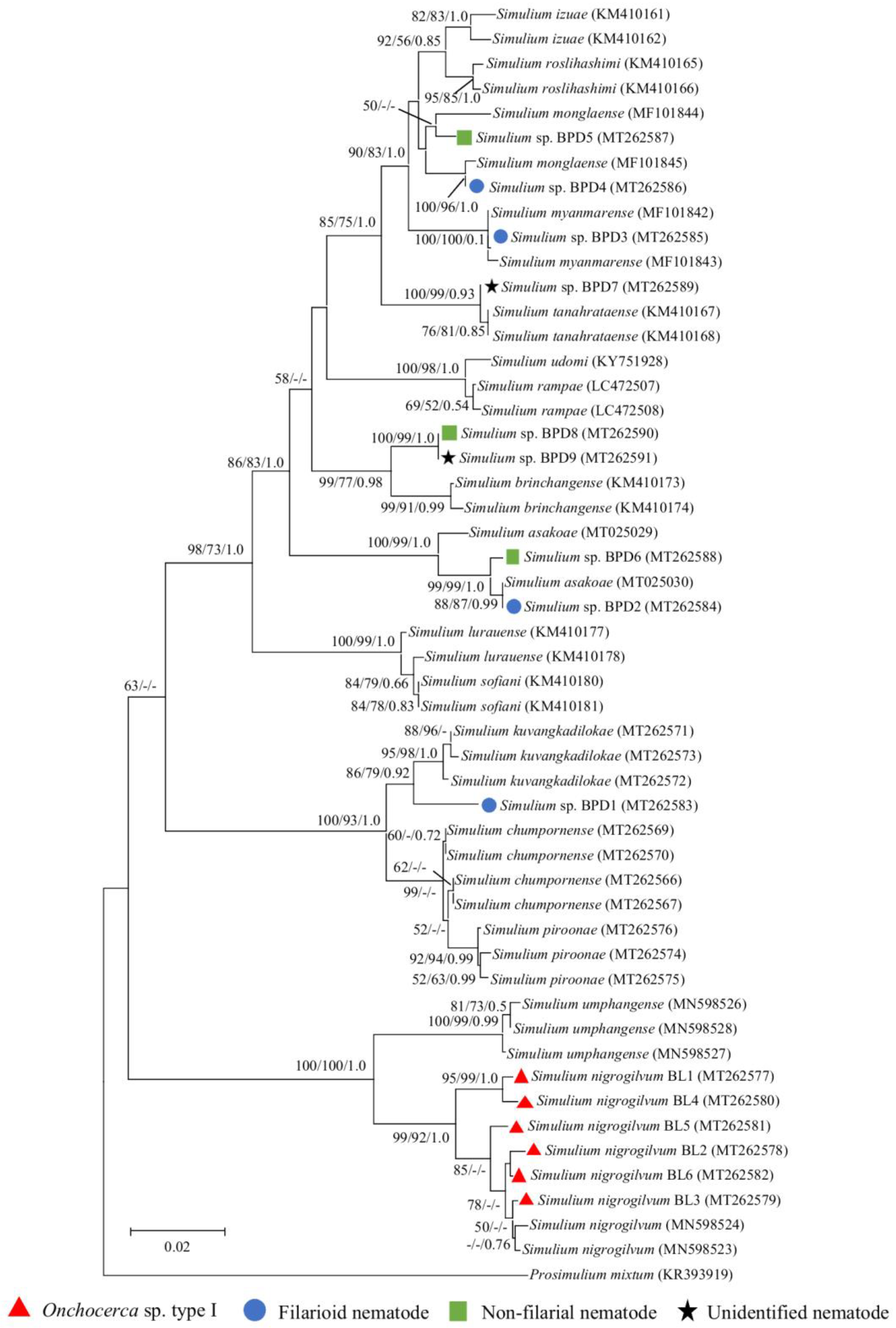
2.3. Molecular Identification of Recovered Larvae
2.3.1. Ban Lek
2.3.2. Ban Pang Dang
2.4. Molecular Identification of Infected Black Flies
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Study Areas and Adult Female Black Fly Collections
4.3. Morphological and Molecular Identification of Female Black Flies
4.4. Dissections of Black Flies and Morphological and Molecular Identification of Recovered Larvae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Crosskey, R.W. The Natural History of Blackflies; Wiley: Chichester, UK, 1990; p. 711. [Google Scholar]
- Takaoka, H.; Fukuda, M.; Otsuka, Y.; Aoki, C.; Uni, S.; Bain, O. Blackfly vectors of zoonotic onchocerciasis in Japan. Med. Vet. Entomol. 2012, 26, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M.; Uni, S.; Igari, T.; Utsumi, Y.; Otsuka, Y.; Nakatani, J.; Uga, S.; Hara, T.; Hasegawa, H.; Takaoka, H. Human case of Onchocerca dewittei japonica infection in Fukushima, Northeastern Honshu, Japan. Parasitol. Int. 2019, 72, 101943. [Google Scholar] [CrossRef] [PubMed]
- Wesolowska, M.; Zajac-Pytrus, H.; Masny, A.; Pytrus, W.; Knysz, B.; Golab, E.; Salamatin, R. Onchocerca jakutensis ocular infection in Poland: A new vector-borne human health risk? Parasit. Vectors 2020, 13, 61. [Google Scholar] [CrossRef] [Green Version]
- Lefoulon, E.; Giannelli, A.; Makepeace, B.L.; Mutafchiev, Y.; Townson, S.; Uni, S.; Verocai, G.G.; Otranto, D.; Martin, C. Whence river blindness? The domestication of mammals and host-parasite co-evolution in the nematode genus Onchocerca. Int. J. Parasitol. 2017, 47, 457–470. [Google Scholar] [CrossRef]
- Uni, S.; Mat Udin, A.S.; Agatsuma, T.; Junker, K.; Saijuntha, W.; Bunchom, N.; Fukuda, M.; Martin, C.; Lefoulon, E.; Labat, A.; et al. Description, molecular characteristics and Wolbachia endosymbionts of Onchocerca borneensis Uni, Mat Udin & Takaoka n. sp. (Nematoda: Filarioidea) from the Bornean bearded pig Sus barbatus Müller (Cetartiodactyla: Suidae) of Sarawak, Malaysia. Parasit. Vectors 2020, 13, 50. [Google Scholar] [CrossRef] [PubMed]
- Jumpato, W.; Tangkawanit, U.; Wongpakam, K.; Pramual, P. Molecular detection of Leucocytozoon (Apicomplexa: Haemosporida) in black flies (Diptera: Simuliidae) from Thailand. Acta Trop. 2019, 190, 228–234. [Google Scholar] [CrossRef]
- Thaijarern, J.; Tangkawanit, U.; Wongpakam, K.; Pramual, P. Molecular detection of Trypanosoma (Kinetoplastida: Trypanosomatidae) in black flies (Diptera: Simuliidae) from Thailand. Acta Trop. 2019, 200, 105196. [Google Scholar] [CrossRef]
- Pramual, P.; Thaijarern, J.; Tangkawanit, U.; Wongpakam, K. Molecular identification of blood meal sources in black flies (Diptera: Simuliidae) suspected as Leucocytozoon vectors. Acta Trop. 2020, 205, 105383. [Google Scholar] [CrossRef]
- Fukuda, M.; Choochote, W.; Bain, O.; Aoki, C.; Takaoka, H. Natural infections with filarial larvae in two species of black flies (Diptera: Simuliidae) in northern Thailand. Jpn. J. Trop. Med. Hyg. 2003, 31, 99–102. [Google Scholar] [CrossRef] [Green Version]
- Takaoka, H.; Choochote, W.; Aoki, C.; Fukuda, M.; Bain, O. Black flies (Diptera: Simuliidae) attracted to humans and water buffalos and natural infections with filarial larvae, probably Onchocerca sp., in northern Thailand. Parasite 2003, 10, 3–8. [Google Scholar] [CrossRef] [Green Version]
- Ishii, Y.; Choochote, W.; Bain, O.; Fukuda, M.; Otsuka, Y.; Takaoka, H. Seasonal and diurnal biting activities and zoonotic filarial infections of two Simulium species (Diptera: Simuliidae) in northern Thailand. Parasite 2008, 15, 121–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takaoka, H.; Bain, O. Infections of blackflies (Diptera: Simuliidae) with three types of zoonotic Onchocerca larvae in Oita, Japan. Jpn. J. Trop. Med. Hyg. 1990, 18, 1–10. [Google Scholar] [CrossRef]
- Fukuda, M.; Takaoka, H.; Uni, S.; Bain, O. Infective larvae of five Onchocerca species from experimentally infected Simulium species in an area of zoonotic onchocerciasis in Japan. Parasite 2008, 15, 111–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, M.; Otsuka, Y.; Uni, S.; Bain, O.; Takaoka, H. Molecular identification of infective larvae of three species of Onchocerca found in wild-caught females of Simulium bidentatum in Japan. Parasite 2010, 17, 39–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeung, A.; Srisuka, W.; Aupalee, K.; Fukuda, M.; Otsuka, Y.; Taai, K.; Maleewong, W.; Takaoka, H. Natural infections with larvae of Onchocerca species type I in the human-biting black fly, Simulium nigrogilvum (Diptera: Simuliidae), in western Thailand. Acta Trop. 2020, 204, 105344. [Google Scholar] [CrossRef] [PubMed]
- Bain, O.; Chabaud, A.G. Atlas of infective larvae of filariae. Trop. Med. Parasitol. 1986, 37, 301–340. [Google Scholar] [PubMed]
- Srisuka, W.; Takaoka, H.; Otsuka, Y.; Fukuda, M.; Thongsahuan, S.; Taai, K.; Choochote, W.; Saeung, A. Seasonal biodiversity of black flies (Diptera: Simuliidae) and evaluation of ecological factors influencing species distribution at Doi Pha Hom Pok National Park, Thailand. Acta Trop. 2015, 149, 212–219. [Google Scholar] [CrossRef]
- Ittiponpanya, N. A whole-year study of the flying activity of black fly attracted to humans in Doi Suthep-Pui National Park. MSc Thesis, Chiang Mai University, Chiang Mai, Thailand, 2006. [Google Scholar]
- Choochote, W.; Takaoka, H.; Fukuda, M.; Otsuka, Y.; Aoki, C.; Eshima, N. Seasonal abundance and daily flying activity of black flies (Diptera: Simuliidae) attracted to human baits in Doi Inthanon National Park, northern Thailand. Med. Entomol. Zool. 2005, 56, 335–348. [Google Scholar] [CrossRef] [Green Version]
- Pramual, P.; Thaijarern, J.; Wongpakam, K. DNA barcoding of human-biting black flies (Diptera: Simuliidae) in Thailand. Acta Trop. 2016, 164, 33–40. [Google Scholar] [CrossRef]
- Ferri, E.; Barbuto, M.; Bain, O.; Galimberti, A.; Uni, S.; Guerrero, R.; Ferte, H.; Bandi, C.; Martin, C.; Casiraghi, M. Integrated taxonomy: Traditional approach and DNA barcoding for the identification of filarioid worms and related parasites (Nematoda). Front. Zool. 2009, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Verocai, G.G.; Nelson, K.J.; Callahan, R.T.; Wekesa, J.W.; Hassan, H.K.; Hoberg, E.P. A cryptic species of Onchocerca (Nematoda: Onchocercidae) in blackflies (Simulium spp.) from southern California, USA. Parasites Vectors 2018, 11, 547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Low, V.L.; Srisuka, W.; Saeung, A.; Tan, T.K.; Ya’cob, Z.; Yeong, Y.S.; Takaoka, H. DNA Barcoding of Simulium asakoae (Diptera: Simuliidae) From Northern Thailand. [published online ahead of print]. J. Med. Entomol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Jomkumsing, P.; Tangkawanit, U.; Wongpakam, K.; Pramual, P. Who is biting you? DNA barcodes reveal cryptic diversity in human-biting black flies (Diptera: Simuliidae). Acta Trop. 2019, 196, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Low, V.L.; Sofian-Azirun, M.; Norma-Rashid, Y. Playing hide-and-seek with the tiny dragonfly: DNA barcoding discriminates multiple lineages of Nannophya pygmaea in Asia. J. Insect Conserv. 2016, 20, 339–343. [Google Scholar] [CrossRef]
- Takaoka, H.; Srisuka, W.; Low, V.L.; Maleewong, W.; Saeung, A. Two new species of Simulium (Gomphostilbia) (Diptera: Simuliidae) from Myanmar, and their phylogenetic relationships with related species in the S. asakoae species-group. Acta Trop. 2017, 176, 39–50. [Google Scholar] [CrossRef]
- Pramual, P.; Simwisat, K.; Martin, J. Identification and reassessment of the specific status of some tropical freshwater midges (Diptera: Chironomidae) using DNA barcode data. Zootaxa 2016, 4072, 39–60. [Google Scholar] [CrossRef]
- Pace, N.R.; Olsen, G.J.; Woese, C.R. Ribosomal RNA phylogeny and the primary lines of evolutionary descent. Cell 1986, 45, 325–326. [Google Scholar] [CrossRef]
- Wang, J.; Xu, F.; Liu, X.; Wang, G. Molecular phylogeny of entomopathogenic nematodes (Mermithidae) inferred from DNA sequences of 18S rDNA, 28S rDNA and COI genes. Acta Zool. Sin. 2007, 53, 835–844. [Google Scholar]
- Jitklang, S.; Ahantarig, A.; Kuvangkadilok, C.; Baimai, V.; Adler, P.H. Parasites of larval black flies (Diptera: Simuliidae) in Thailand. Songklanakarin J. Sci. Technol. 2012, 34, 597–599. [Google Scholar]
- Digiani, M.C.; Sutton, C.A. New reports and a redescription of Porrocaecum heteropterum (Diesing, 1851) (Ascarididae), a rare nematode parasitic in South American.threskiornithid birds1. Syst. Parasitol. 2001, 49, 1–6. [Google Scholar] [CrossRef]
- Crainey, J.L.; Wilson, M.D.; Post, R.J. An 18S ribosomal DNA barcode for the study of Isomermis lairdi, a parasite of the blackfly Simulium damnosum s.l. Med. Vet. Entomol. 2009, 23, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Takaoka, H.; Srisuka, W.; Saeung, A. Checklist and keys for the black flies (Diptera: Simuliidae) of Thailand. Med. Entomol. Zool. 2019, 70, 53–77. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Conflitti, I.M.; Shields, G.F.; Currie, D.C. A “complex” problem: Delimiting sibling species boundaries in black flies (Diptera: Simuliidae). Can. Entomol. 2012, 144, 323–336. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Zhou, S.; Fu, X.; Pei, P.; Kucka, M.; Liu, J.; Tang, L.; Zhan, T.; He, S.; Chan, Y.F.; Rodelsperger, C.; et al. Characterization of a non-sexual population of Strongyloides stercoralis with hybrid 18S rDNA haplotypes in Guangxi, Southern China. PLoS Negl. Trop. Dis. 2019, 13, e0007396. [Google Scholar] [CrossRef] [PubMed]
- Casiraghi, M.; Anderson, T.J.; Bandi, C.; Bazzocchi, C.; Genchi, C. A phylogenetic analysis of filarial nematodes: Comparison with the phylogeny of Wolbachia endosymbionts. Parasitology 2001, 122, 93–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casiraghi, M.; Bain, O.; Guerrero, R.; Martin, C.; Pocacqua, V.; Gardner, S.L.; Franceschi, A.; Bandi, C. Mapping the presence of Wolbachia pipientis on the phylogeny of filarial nematodes: Evidence for symbiont loss during evolution. Int. J. Parasitol. 2004, 34, 191–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Month | Simulium Species | No. Collected | No. Infected | No. L3 | % Infected | |
|---|---|---|---|---|---|---|---|
| Hot | March | S. chumpornense | 317 | 477 | 0 | 0 | 0 |
| S. asakoae species-group | 111 | 0 | 0 | 0 | |||
| S. nigrogilvum | 36 | 1 | 1 | 2.78 | |||
| S. striatum species-group | 13 | 0 | 0 | 0 | |||
| May | S. nigrogilvum | 151 | 153 | 2 | 3 | 1.32 | |
| S. chamlongi | 1 | 0 | 0 | 0 | |||
| S. doipuiense complex | 1 | 0 | 0 | 0 | |||
| Rainy | August | S. nigrogilvum | 34 | 48 | 0 | 0 | 0 |
| S. asakoae species-group | 13 | 0 | 0 | 0 | |||
| S. striatum species-group | 1 | 0 | 0 | 0 | |||
| October | S. nigrogilvum | 48 | 65 | 1 | 2 | 2.08 | |
| S. asakoae species-group | 14 | 0 | 0 | 0 | |||
| S. manooni | 1 | 0 | 0 | 0 | |||
| S. doipuiense complex | 1 | 0 | 0 | 0 | |||
| S. striatum species-group | 1 | 0 | 0 | 0 | |||
| Dry-cool | December | S. nigrogilvum | 40 | 59 | 0 | 0 | 0 |
| S. doipuiense complex | 10 | 0 | 0 | 0 | |||
| S. asakoae species-group | 7 | 0 | 0 | 0 | |||
| S. chamlongi | 1 | 0 | 0 | 0 | |||
| S. striatum species-group | 1 | 0 | 0 | 0 | |||
| January | S. asakoae species-group | 3 | 3 | 0 | 0 | 0 | |
| Total | 805 | 4 | 6 | - | |||
| Season | Month | Simulium Species | No. Collected | No. Infected | No. Larva | % Infected | |||
|---|---|---|---|---|---|---|---|---|---|
| FL | NFL | UID | |||||||
| Hot | March | S. chumpornense | 2249 | 2616 | 0 | 0 | 0 | 0 | 0 |
| S. asakoae species-group | 363 | 0 | 0 | 0 | 0 | 0 | |||
| S. nigrogilvum | 3 | 0 | 0 | 0 | 0 | 0 | |||
| S. sp. | 1 | 1 | 1 L1 | 0 | 0 | 100 | |||
| June | S. asakoae species-group | 157 | 157 | 1 | 0 | 0 | 1 Mf | 0.64 | |
| Rainy * | September | S. asakoae species-group | 635 | 636 | 2 | 1 L3 | 1 L3 | 0 | 0.31 |
| S. striatum species-group | 1 | 0 | 0 | 0 | 0 | 0 | |||
| Dry-cool | November | S. asakoae species-group | 368 | 368 | 3 | 2 L3 | 1 L3 | 1 L1 | 0.82 |
| January | S. asakoae species-group | 807 | 820 | 2 | 1 L3 | 1 L3 | 0 | 0.25 | |
| S. striatum species-group | 8 | 0 | 0 | 0 | 0 | 0 | |||
| S. chamlongi | 2 | 0 | 0 | 0 | 0 | 0 | |||
| S. tani | 1 | 0 | 0 | 0 | 0 | 0 | |||
| S. bullatum | 1 | 0 | 0 | 0 | 0 | 0 | |||
| S. lampangense | 1 | 0 | 0 | 0 | 0 | 0 | |||
| Total | 4597 | 9 | 5 | 3 | 2 | - | |||
| Black Fly Species (No.). | Larval Stage (No.) | Body Length × Body Width (μm) | Molecular Identification |
|---|---|---|---|
| Filarial nematodes S. nigrogilvum (4) S. asakoae species-group (3) Simulium sp. (1) | L3 (6) L3 (4) L1 (1) | (1027.1–1339.5) × (25.6–29.6) (576.1–688.8) × (26.9–28.6) 270.3 × 18.6 | 1Onchocerca sp. type I 1 Filarioid nematode 1 Filarioid nematode |
| Non-filarial nematodes S. asakoae species-group (3) | L3 (1) L3 (1) L3 (1) | 2864.1 × 102.7 446.4 × 20.3 606.2 × 26.9 | 3 Mermithid nematode 2 Rhabditida nematode 3 Ascaridoid nematode |
| Unidentified nematodes S. asakoae species-group (2) | Mf (1) L1 (1) | 336.6 × 9.6 468.9 × 16.2 | N/A |
| Region Amplified | Primer | Sequence (5′-3′) | Annealing T (°C) | Product (bp) | Ref | |
|---|---|---|---|---|---|---|
| cox1 | Fw | COIintF | TGATTGGTGGTTTTGGTAA | 48 | ~689 | [44] |
| Rev | COIintR | ATAAGTACGAGTATCAATATC | ||||
| 12S rRNA | Fw | 12SF | GTTCCAGAATAATCGGCT | 50 | ~502 | [45] |
| Rev | 12SR | ATTGACGGATGRTTTGTACC | ||||
| 18S rRNA (SSU HVR-I) | Fw | RS5401 | AAAGATTAAGCCATGCATG | 50 | ~919 | [43] |
| Rev | RS5402 | CATTCTTGGCAAATGCTTTCG | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aupalee, K.; Saeung, A.; Srisuka, W.; Fukuda, M.; Streit, A.; Takaoka, H. Seasonal Filarial Infections and Their Black Fly Vectors in Chiang Mai Province, Northern Thailand. Pathogens 2020, 9, 512. https://doi.org/10.3390/pathogens9060512
Aupalee K, Saeung A, Srisuka W, Fukuda M, Streit A, Takaoka H. Seasonal Filarial Infections and Their Black Fly Vectors in Chiang Mai Province, Northern Thailand. Pathogens. 2020; 9(6):512. https://doi.org/10.3390/pathogens9060512
Chicago/Turabian StyleAupalee, Kittipat, Atiporn Saeung, Wichai Srisuka, Masako Fukuda, Adrian Streit, and Hiroyuki Takaoka. 2020. "Seasonal Filarial Infections and Their Black Fly Vectors in Chiang Mai Province, Northern Thailand" Pathogens 9, no. 6: 512. https://doi.org/10.3390/pathogens9060512
APA StyleAupalee, K., Saeung, A., Srisuka, W., Fukuda, M., Streit, A., & Takaoka, H. (2020). Seasonal Filarial Infections and Their Black Fly Vectors in Chiang Mai Province, Northern Thailand. Pathogens, 9(6), 512. https://doi.org/10.3390/pathogens9060512