Expression of Serum Exosomal miRNA 122 and Lipoprotein Levels in Dogs Naturally Infected by Leishmania infantum: A Preliminary Study

 ,
,  , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Samples Collection and Hemato-Biochemical Analysis
2.3. Exosomes Isolation and Mirna Detection
2.4. Statistical Analysis
3. Results
3.1. Clinical Examination and Blood Tests
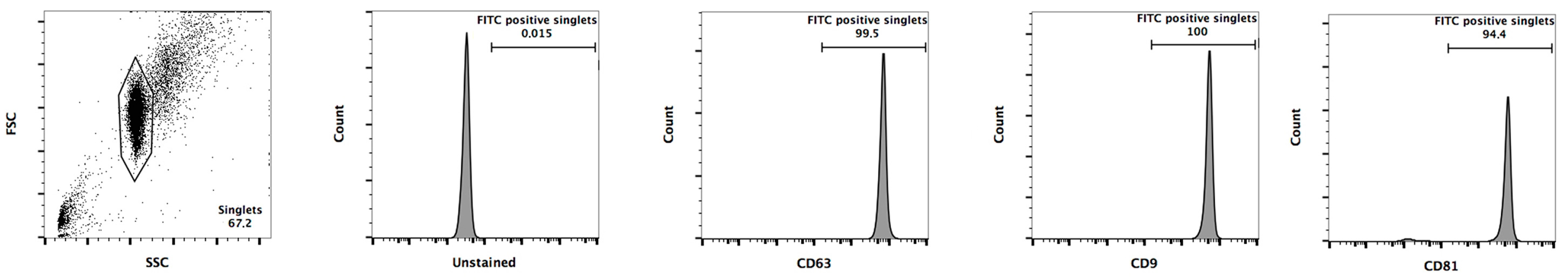
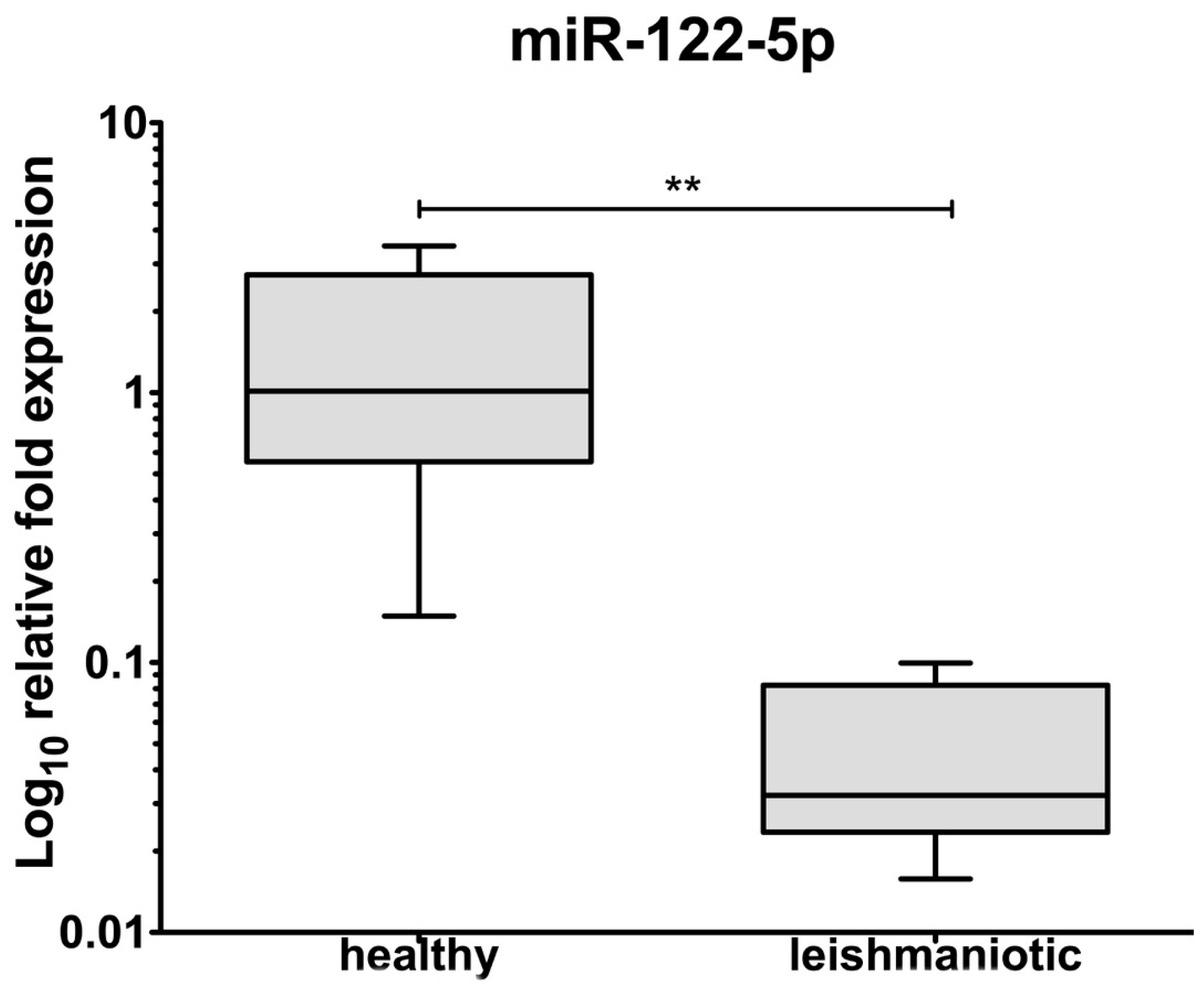
3.2. Exosomes Isolation and miRNA Detection
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A
Appendix A.1. Ethical and Regulatory Approval
Appendix A.2. Exosomes Isolation
Appendix A.3. Characterization
Appendix A.4. Analytical Methods
Appendix A.5. Exosomal Protein Isolation and Quantification
Appendix A.6. miRNAs Isolation and miR-122-5p and RNU6 Analysis
References
- Beattie, L.; Peltan, A.; Maroof, A.; Kirby, A.; Brown, N.; Coles, M.; Smith, D.F.; Kaye, P.M. Dynamic imaging of experimental Leishmania donovani-induced hepatic granulomas detects Kupffer cell-restricted antigen presentation to antigen-specific CD8 T cells. PLoS Pathog. 2010, 6, e1000805. [Google Scholar] [CrossRef] [Green Version]
- Rabhi, I.; Rabhi, S.; Ben-Othman, R.; Rasche, A.; Daskalaki, A.; Trentin, B.; Guizani-Tabbane, L. Transcriptomic signature of Leishmania infected mice macrophages: A metabolic point of view. PLoS Negl. Trop. Dis. 2012, 6, e1763. [Google Scholar] [CrossRef]
- Rodrigues, M.A.V.; Santos-Mateus, D.; Alexandre-Pires, G.; Valerio-Bolas, A.; Rafael-Fernandes, M.; Pereira, M.A. Kupffer cells and blood derived macrophages respond differently to L. Infantum parasites. In Proceedings of the 6th World Congress on Leishmaniasis, Toledo, Spain, 16–20 May 2017. [Google Scholar]
- Osorio y Fortea, J.; de La Llave, E.; Regnault, B.; Coppee, J.Y.; Milon, G.; Lang, T.; Prina, E. Transcriptional signatures of BALB/c mouse macrophages housing multiplying Leishmania amazonensis amastigotes. BMC Genom. 2009, 10, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, C.W.; McLeod, R.; Rice, D.W.; Ginger, M.; Chance, M.L.; Goad, L.J. Fatty acid and sterol metabolism: Potential antimicrobial targets in apicomplexan and trypanosomatid parasitic protozoa. Mol. Biochem. Parasitol. 2003, 126, 129–142. [Google Scholar] [CrossRef]
- Lee, M.G.S.; Yen, F.T.; Zhang, Y.; Bihain, B. Acquisition of lipoproteins in the procyclic form of Trypanosoma brucei. Mol. Biochem. Parasitol. 1999, 100, 153–162. [Google Scholar]
- Andrade-Neto, V.V.; Cicco, N.N.; Cunha-Junior, E.F.; Canto-Cavalheiro, M.M.; Atella, G.C.; Torres-Santos, E.C. The pharmacological inhibition of sterol biosynthesis in Leishmania is counteracted by enhancement of LDL endocytosis. Acta Trop. 2011, 119, 194–198. [Google Scholar] [CrossRef]
- De Cicco, N.N.; Pereira, M.G.; Correa, J.R.; Andrade-Neto, V.V.; Saraiva, F.B.; Chagas-Lima, A.C.; Atella, G.C. LDL uptake by Leishmania amazonensis: Involvement of membrane lipid microdomains. Exp. Parasitol. 2012, 130, 330–340. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, J.; Das, S.; Guha, R.; Ghosh, D.; Naskar, K.; Das, A.; Roy, S. Hyperlipidemia offers protection against Leishmania donovani infection: Role of membrane cholesterol. J. Lipid Res. 2012, 53, 25602572. [Google Scholar] [CrossRef] [Green Version]
- Nieto, C.G.; Barrera, R.; Habela, M.A.; Navarrete, I.; Molina, C.; Jimenez, A.; Serrera, J.L. Changes in the plasma concentrations of lipids and lipoprotein fractions in dogs infected with Leishmania infantum. Vet. Parasitol. 1992, 44, 175–182. [Google Scholar] [CrossRef]
- Durgut, R.; Dalkilinç, D.; Güzeln, M. Evaluation of the serum lipid profiles in dogs with symptomatic visceral leishmaniasis. Kafkas Univ. Vet. Fak. Derg. 2012, 18, 585–588. [Google Scholar]
- Gültekin, M.; Paşa, S.; Ural, K.; Balıkçı, C.; Ekren Aşıcı, G.S.; Gültekin, G. Oxidative status and lipid profile among dogs at different stages of visceral leishmaniasis. Turk. Parazitol. Derg. 2017, 41, 183–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilczynska, A.; Bushell, M. The complexity of miRNA-mediated repression. Cell Death Differ. 2015, 22, 22–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, S.; Willenbrock, S.; Nolte, I.; Escobar, H.M. Comparison of non-coding RNAs in human andcanine cancer. Front. Genet. 2013, 4, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koenig, E.M.; Fisher, C.; Bernard, H.; Wolenski, F.S.; Gerrein, J.; Carsillo, M.; Gallacher, M.; Tse, A.; Peters, R.; Smith, A.; et al. The beagle dog MicroRNA tissue atlas: Identifying translatable biomarkers of organ toxicity. BMC Genom. 2016, 17, 649. [Google Scholar] [CrossRef] [Green Version]
- Thind, A.; Wilson, C. Exosomal miRNAs as cancer biomarkers and therapeutic targets. J. Extracell. Vesicles 2016, 5, 31292. [Google Scholar] [CrossRef]
- Iaccino, E.; Mimmi, S.; Dattilo, V.; Marino, F.; Candeloro, P.; Di Loria, A.; Marimpietri, D.; Pisano, A.; Albano, F.; Vecchio, E.; et al. Monitoring multiple myeloma by idiotype-specific peptide binders of tumor-derived exosomes. Mol. Cancer 2017, 16, 159. [Google Scholar] [CrossRef]
- Manna, I.; Iaccino, E.; Dattilo, V.; Barone, S.; Vecchio, E.; Mimmi, S.; Filippelli, E.; Demonte, G.; Polidoro, S.; Granata, A.; et al. Exosome-associated miRNA profile as a prognostic tool for therapy response monitoring in multiple sclerosis patients. FASEB J. 2018, 32, 4241–4246. [Google Scholar] [CrossRef] [Green Version]
- Vlassov, A.V.; Magdaleno, S.; Setterquist, R.; Conrad, R. Exosomes: Current knowledge of their composition, biological functions, and diagnostic and therapeutic potentials. Biochim. Biophys. Acta 2012, 1820, 940–948. [Google Scholar] [CrossRef]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, J.; Das, S.; Bose, M.; Roy, S.; Battacharyya, S.N. Leishmania donovani targets Dicer1 to downregulate miR-122, lower serum cholesterol and facilitate murine liver infection. Cell Host Microbe 2013, 13, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Solano-Gallego, L.; Koutinas, A.; Miró, G.; Cardoso, L.; Pennisi, M.G.; Ferrer, L.; Bourdeau, P.; Oliva, G.; Baneth, G. Directions for the diagnosis, clinical staging, treatment and prevention of canine leishmaniosis. Vet. Parasitol. 2009, 165, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Solano-Gallego, L.; Villanueva-Saz, S.; Carbonell, M.; Trotta, M.; Furlanello, T.; Natale, A. Serological diagnosis of canine leishmaniosis: Comparison of three commercial ELISA tests (Leiscan®, ID Screen® and Leishmania 96®), a rapid test (Speed Leish K®) and an in-house IFAT. Parasites Vectors 2014, 7, 111. [Google Scholar] [CrossRef] [Green Version]
- Fridewald, W.T.; Lavy, R.; Fredricson, D.S. Estimation of low density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar]
- Rizzi, T.E.; Meinkoth, J.H.; Clinkenbeard, K.D. Shalms Veterinary Hematology, 6th ed.; Weiss, D.J., Wardrop, K.J., Eds.; Wiley-Blackwell: San Diego, CA, USA, 2010; p. 802. [Google Scholar]
- Kaneko, J.J.; Harvey, J.W.; Bruss, M.L. (Eds.) Clinical Biochemistry of Domestic Animals, 6th ed.; Academic Press: San Diego, CA, USA, 2008; pp. 889–895. [Google Scholar]
- Coppo, N.B.; Coppo, J.A.; Lazarte, M.A. Intervalos de confianza para colesterol ligado a lipoproteínas de alta y baja densidad en suero de bovinos, equinos, porcinos y caninos. Rev. Vet. 2003, 14, 1. [Google Scholar]
- Carvalho, M.D.; Alonso, D.P.; Vendrame, C.M.; Costa, D.L.; Costa, C.H.; Werneck, G.L.; Ribolla, P.E.; Goto, H. Lipoprotein lipase and PPAR alpha gene polymorphisms, increased very-low-density lipoprotein levels, and decreased high-density lipoprotein levels as risk markers for the development of visceral leishmaniasis by Leishmania infantum. Med. Inflamm. 2014, 230129. [Google Scholar] [CrossRef] [Green Version]
- Gordon, B.R.; Parker, T.S.; Levine, D.M.; Saal, S.D.; Wang, J.C.; Sloan, B.J.; Barie, P.S.; Rubin, A.L. Relationship of hypolipidemia to cytokine concentrations and outcomes in critically ill surgical patients. Crit. Care Med. 2001, 29, 1563–1568. [Google Scholar] [CrossRef]
- Jin, W.; Sun, G.S.; Marchadier, D.; Octtaviani, E.; Glick, J.M.; Rader, D.J. Endothelial cells secrete triglyceride lipase and phospholipase activities in response to cytokines as a result of endothelial lipase. Circ. Res. 2003, 92, 644–650. [Google Scholar] [CrossRef] [Green Version]
- Bekaert, E.D.; Dole, E.; Dubois, D.Y.; Bouma, M.E.; Lontie, J.F.; Kallel, R.; Malmendier, C.L.; Ayrault-Jarrier, M. Alterations in lipoprotein density classes in infantile visceral Leishmaniasis: Presence of apolipoprotein SAA. Eur. J. Clin. Investig. 1992, 22, 190–199. [Google Scholar] [CrossRef]
- Besler, C.; Luscher, T.F.; Landmesser, U. Molecular mechanisms of vascular effects of high-density lipoprotein: Alterations in cardiovascular disease. EMBO Mol. Med. 2012, 4, 251–268. [Google Scholar] [CrossRef] [Green Version]
- Hafiane, A.; Genest, J. HDL, atherosclerosis, and emerging therapies. Cholesterol 2013, 891403. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Wu, Z.; Riwanto, M.; Gao, S.; Levison, B.S.; Gu, X.; Fu, X.; Wagner, M.A.; Besler, C.; Gerstenecker, G. Myeloperoxidase, paraoxonase-1, and HDL form a functional ternary complex. J. Clin. Investig. 2013, 123, 3815–3828. [Google Scholar] [CrossRef] [Green Version]
- Riwanto, M.; Landmesser, U. High density lipoproteins and endothelial functions: Mechanistic insights and alterations in cardiovascular disease. J. Lipid Res. 2013, 54, 3227–3243. [Google Scholar] [CrossRef] [Green Version]
- Brandonisio, O.; Panaro, M.A.; Sisto, M.; Acquafredda, A.; Fumarola, L.; Leogrande, D.; Mitolo, V. Nitric oxide production by Leishmania-infected macrophages and modulation by cytokines and prostaglandins. Parassitologia 2001, 43 (Suppl. 1), 1–6. [Google Scholar]
- Rodrigues Santos, A.M.; Ozaki, C.Y.; Goto, H. Effect of plasma lipoproteins on the parasitism of thp-1 cells infected with Leishmania (Leishmania) infantum. In Proceedings of the 6th World Congress on Leishmaniasis, Toledo, Spain, 16–20 May 2017. [Google Scholar]
- Sanchez, M.A.; Diaz, N.L.; Zerpa, O.; Negron, E.; Convit, J.; Tapia, F.J. Organ-specific immunity in canine visceral leishmaniasis: Analysis of symptomatic and asymptomatic dogs naturally infected with Leishmania chagasi. Am. J. Trop. Med. Hyg. 2004, 70, 618–624. [Google Scholar] [CrossRef]
- Li, J.; Liu, K.; Liu, Y.; Xu, Y.; Zhang, F.; Yang, H.; Liu, J.; Pan, T.; Chen, J.; Wu, M.; et al. Exosomes mediate the cell-to-cell transmission of IFN-α-induced antiviral activity. Nat. Immunol. 2013, 14, 793–803. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Healthy | Leishmaniotic | Reference Range [25,26,27] | p-Value |
|---|---|---|---|---|
| RBC (×106/dL) | 6.5 ± 0.6 | 5.5 ± 1.6 | 5.5–8.5 | 0.01 |
| Hb (g/dL) | 14.9 ± 1.5 | 10.7 ± 3.1 | 12–18 | 0.0001 |
| Hct (%) | 44.3 ± 3.2 | 34.4 ± 9.6 | 37–55 | 0.0009 |
| MCV (fL) | 69.2 ± 3.4 | 63.4 ± 4.8 | 60–77 | 0.008 |
| MCH (pg) | 21.1 ± 1.4 | 19.6 ± 1.4 | 20.5–24.2 | <0.0001 |
| MCHC (%) | 33.8 ± 1.1 | 30.9 ± 0.9 | 32–36 | <0.0001 |
| PLT (×103/mm3) | 295.4 ± 89.5 | 279.3 ± 74.4 | 200–500 | 0.7 |
| WBC (×103/mm3) | 10.3 ± 1.9 | 10.9 ± 1.5 | 6–17 | 0.5 |
| Azotemia (mg/dL) | 42.2 ± 6.0 | 48.0 ± 17.6 | 21–59 | 0.7 |
| Crea (mg/dL) | 0.8 ± 0.2 | 1.4 ± 1.0 | 0.5–1.5 | 0.7 |
| ALT (U/L) | 41.9 ± 11.8 | 35.1 ± 8.3 | 21–102 | 0.1 |
| AST (U/L) | 34.6 ± 10.2 | 34.3 ± 12.9 | 23–66 | 0.9 |
| ALP (U/L) | 111.8 ± 28.2 | 110.3 ± 32.1 | 20–156 | 0.9 |
| BIL (mg/dL) | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.1–0.5 | 1 |
| GGT (U/L) | 3.0 ± 1.4 | 2.4 ± 1.2 | 1.2–6.4 | 0.4 |
| GLU (mg/dL) | 73.8 ± 12.0 | 74.1 ± 13.5 | 65–118 | 0.9 |
| TRIG (mg/dL) | 80.0 ± 23.0 | 86.1 ± 22.3 | 20–112 | 0.5 |
| CHOL (mg/dL) | 193.6 ± 49.3 | 223.7 ± 63.8 | 135–270 | 0.3 |
| HDL (mg/dL) | 89.7 ± 19.1 | 47.4 ± 26 | 49–165 | <0.0001 |
| LDL (mg/dL) | 87.9 ± 49.5 | 159.1 ± 51.1 | 5–86 | 0.01 |
| TP (g/dL) | 6.2 ± 0.5 | 9.1 ± 1.7 | 5.4–7.1 | 0.0003 |
| Alb | 3.4 ± 0.4 | 2.9 ± 0.5 | 2.6–3.3 g/dL | 0.02 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loria, A.D.; Dattilo, V.; Santoro, D.; Guccione, J.; De Luca, A.; Ciaramella, P.; Pirozzi, M.; Iaccino, E. Expression of Serum Exosomal miRNA 122 and Lipoprotein Levels in Dogs Naturally Infected by Leishmania infantum: A Preliminary Study. Animals 2020, 10, 100. https://doi.org/10.3390/ani10010100
Loria AD, Dattilo V, Santoro D, Guccione J, De Luca A, Ciaramella P, Pirozzi M, Iaccino E. Expression of Serum Exosomal miRNA 122 and Lipoprotein Levels in Dogs Naturally Infected by Leishmania infantum: A Preliminary Study. Animals. 2020; 10(1):100. https://doi.org/10.3390/ani10010100
Chicago/Turabian StyleLoria, Antonio Di, Vincenzo Dattilo, Domenico Santoro, Jacopo Guccione, Adriana De Luca, Paolo Ciaramella, Marinella Pirozzi, and Enrico Iaccino. 2020. "Expression of Serum Exosomal miRNA 122 and Lipoprotein Levels in Dogs Naturally Infected by Leishmania infantum: A Preliminary Study" Animals 10, no. 1: 100. https://doi.org/10.3390/ani10010100
APA StyleLoria, A. D., Dattilo, V., Santoro, D., Guccione, J., De Luca, A., Ciaramella, P., Pirozzi, M., & Iaccino, E. (2020). Expression of Serum Exosomal miRNA 122 and Lipoprotein Levels in Dogs Naturally Infected by Leishmania infantum: A Preliminary Study. Animals, 10(1), 100. https://doi.org/10.3390/ani10010100