Understanding the Toxin Effects of β-Zearalenol and HT-2 on Bovine Granulosa Cells Using iTRAQ-Based Proteomics


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation, Culture, and Identification of Bovine GCs
2.2. Cell Treatment and Cell Viability Analysis
2.3. Protein Extraction, Digestion, and iTRAQ Labeling
2.4. Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS) Analysis
2.5. Protein Identification and Quantitative Analysis
2.6. Biological Function Enrichment Analysis
2.7. Determination of Hormone
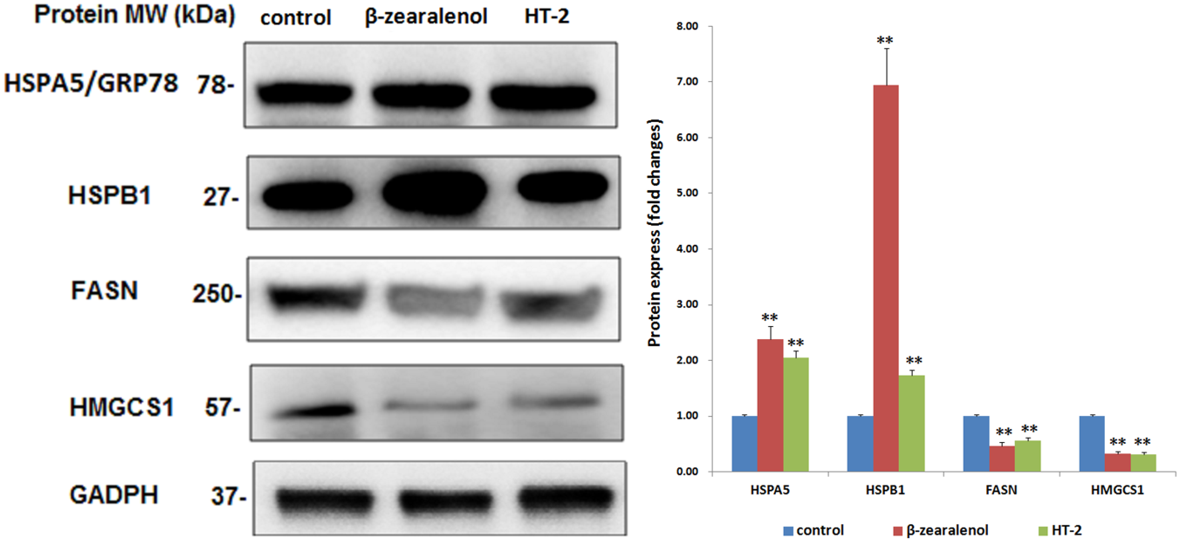
2.8. Western Blotting
2.9. Apoptosis Detection by Flow Cytometry and Caspase-3 Activity Measurement
2.10. Measurement the Accumulation of ROS
2.11. Statistical Analysis
3. Results
3.1. Overview of Quantitative Proteomic Analyses
3.2. Differentially Expressed Proteins
3.3. Analysis of GO and KEGG Pathways for Common DEPs
3.4. DEPs Analysis
3.5. Effect of β-Zearalenol or HT-2 on GC Viability and Apoptosis
3.6. Effects of β-Zearalenol or HT-2 on Estradiol (E2) and Progesterone (P4) Production in GCs
3.7. Effect of β-Zearalenol or HT-2 on GCs ROS
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and its metabolites: Effect on human health, metabolism and neutralization methods. Toxicon 2019, 162, 46–56. [Google Scholar] [CrossRef]
- Pitt, J.I.; Miller, J.D. A concise history of mycotoxin research. J. Agric. Food Chem. 2017, 65, 7021–7033. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, K.; Habrowska-Górczynska, D.E.; Piastowska-Ciesielska, A.W. Zearalenone as an endocrine disruptor in humans. Environ. Toxicol. Pharmacol. 2016, 48, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.F.; Girgin, G.; Baydar, T.; Krska, R.; Sulyok, M. Occurrence of multiple mycotoxins and other fungal metabolites in animal feed and maize samples from egypt using lc-ms/ms. J. Sci. Food Agric. 2017, 97, 4419–4428. [Google Scholar] [CrossRef]
- Abdallah, M.F.; Girgin, G.; Baydar, T. Occurrence, prevention and limitation of mycotoxins in feeds. Anim. Nutr. Feed Technol. 2015, 15, 471–490. [Google Scholar] [CrossRef]
- Wu, Q.; Huang, L.; Liu, Z.; Yao, M.; Yuan, Z. A comparison of hepatic in vitro metabolism of t-2 toxin in rats, pigs, chickens, and carp. Xenobiotica 2011, 10, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Engemann, A.; Cramer, B.; Welsch, T.; Yuan, Z.; Humpf, H.U. Intestinal metabolism of t-2 toxin in the pig cecum model. Mycotoxin Res. 2012, 28, 191–198. [Google Scholar] [CrossRef]
- Yang, L.; Tu, D.; Zhao, Z.; Cui, J. Cytotoxicity and apoptosis induced by mixed mycotoxins (t-2 and ht-2 toxin) on primary hepatocytes of broilers in vitro. Toxicon 2017, 129, 1–10. [Google Scholar] [CrossRef]
- Zhang, Y.; Jia, R.X.; Pan, M.H.; Lu, Y.J.; Sun, S.C. Ht-2 toxin affects development of porcine parthenotes by altering dna and histone methylation in oocytes matured in vitro. Theriogenology 2017, 103, 110. [Google Scholar] [CrossRef]
- Zhang, L.; Li, L.; Xu, J.; Pan, M.H.; Sun, S.C. HT-2 toxin exposure induces mitochondria dysfunction and DNA damage during mouse early embryo development. Reprod. Toxicol. 2019, 85, 104–109. [Google Scholar] [CrossRef]
- Zhang, G.L.; Sun, X.F.; Feng, Y.Z.; Li, B.; Li, Y.P.; Yang, F.; Nyachoti, C.M.; Shen, W.; Sun, S.D.; Li, L. Zearalenone exposure impairs ovarian primordial follicle formation via downregulation of Lhx8 expression in vitro. Toxicol. Appl. Pharmacol. 2017, 317, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Zwierzchowski, W.; Przybylowicz, M.; Obremski, K.; Zielonka, L.; SkorskaWyszynska, E.; Gajecka, M.; Polak, M.; Jakimiuk, E.; Jana, B.; Rybarczyk, L.; et al. Level of zearalenone in blood serum and lesions in ovarian follicles of sexually immature gilts in the course of zearalenone micotoxicosis. Pol. J. Vet. Sci. 2005, 8, 209–218. [Google Scholar] [PubMed]
- Maruniakova, N.; Kadasi, A.; Sirotkin, A.V.; Bulla, J.; Kolesarova, A. T-2 toxin and its metabolite ht-2 toxin combined with insulin-like growth factor-i modify progesterone secretion by porcine ovarian granulosa cells. J. Environ. Sci. Health Part A 2014, 49, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Eppig, J.J.; Wigglesworth, K.; Pendola, F.L. The mammalian oocyte orchestrates the rate of ovarian follicular development. Proc. Natl. Acad. Sci. USA 2002, 99, 2890. [Google Scholar] [CrossRef] [Green Version]
- Petro, E.M.; Leroy, J.L.; Van Cruchten, S.J.; Covaci, A.; Jorssen, E.P.; Bols, P.E. Endocrine disruptors and female fertility: Focus on (bovine) ovarian follicular physiology. Theriogenology 2012, 78, 1887–1900. [Google Scholar] [CrossRef]
- Pizzo, F.; Caloni, F.; Schutz, L.F.; Totty, M.L.; Spicer, L.J. Individual and combined effects of deoxynivalenol and alpha-zearalenol on cell proliferation and steroidogenesis of granulosa cells in cattle. Environ. Toxicol. Pharmacol. 2015, 40, 722–728. [Google Scholar] [CrossRef]
- Tatone, C.; Amicarelli, F.; Carbone, M.C.; Monteleone, P.; Caserta, D.; Marci, R.; Artini, P.G.; Piomboni, P.; Focarelli, R. Cellular and molecular aspects of ovarian follicle ageing. Hum. Reprod. Update 2008, 14, 131–142. [Google Scholar] [CrossRef] [Green Version]
- Lagaly, D.V.; Aad, P.Y.; Grado-Ahuir, J.A.; Hulsey, L.B.; Spicer, L.J. Role of adiponectin in regulating ovarian granulosa and theca cell function. Mol. Cell. Endocrinol. 2008, 284, 38–45. [Google Scholar] [CrossRef]
- Spicer, L.J.; Chamberlain, C.S.; Maciel, S.M. Influence of gonadotropins on insulin and insulin-like growth factor I (IGF-I) induced steroid production by bovine granulosa cells. Domest. Anim. Endocrinol. 2002, 22, 237–254. [Google Scholar] [CrossRef]
- Spicer, L.J.; Aad, P.Y. Insulin-like growth factor (IGF) 2 stimulates steroidogenesis and mitosis of bovine granulosa cells through the IGF1 receptor: Role of follicle-stimulating hormone and IGF2 receptor. Biol. Reprod. 2007, 77, 18–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Li, L.; Chen, K.; Li, C.; Wang, Y.; Wang, G. Melatonin alleviates β-zearalenol and HT-2 toxin-induced apoptosis and oxidative stress in bovine ovarian granulosa cells. Environ. Toxicol. Pharmacol. 2019, 68, 52. [Google Scholar] [CrossRef] [PubMed]
- Wenting, D.; Quanjuan, W.; Fengqi, Z.; Jianxin, L.; Hongyun, L. Understanding the regulatory mechanisms of milk production using integrative transcriptomic and proteomic analyses: Improving inefficient utilization of crop by-products as forage in dairy industry. BMC Genom. 2018, 19, 403. [Google Scholar]
- Langhout, D.J.; Spicer, L.J.; Geisert, R.D. Development of a culture system for bovine granulosa cells: effects of growth hormone, estradiol, and gonadotropins on cell proliferation, steroidogenesis, and protein synthesis. J. Anim. Sci. 2008, 69, 3321–3334. [Google Scholar] [CrossRef] [PubMed]
- Pulskamp, K.; Diabate, S.; Krug, H.F. Carbon nanotubes show no sign of acute toxicity but induce intracellular reactive oxygen species in dependence on contaminants. Toxicol. Lett. 2007, 168, 58–74. [Google Scholar] [CrossRef]
- Tiemann, U.; Tomek, W.; Schneider, F.; Vanselow, J. Effects of the mycotoxins and -Zearalenol on regulation of progesterone synthesis in cultured granulose cells from porcine ovaries. Reprod. Toxicol. 2003, 17, 673–681. [Google Scholar] [CrossRef]
- Cortinovis, C.; Caloni, F.; Schreiber, N.; Spicer, L.J. Effects of fumonisin B1 alone and combined with deoxynivalenol or zearalenone on porcine granulosa cell proliferation and steroid production. Theriogenology 2014, 81, 1042–1049. [Google Scholar] [CrossRef]
- Zhang, Y.; Han, J.; Zhu, C.C.; Tang, F.; Cui, X.S.; Kim, N.H.; Sun, S.C. Exposure to ht-2 toxin causes oxidative stress induced apoptosis/autophagy in porcine oocytes. Sci. Rep. 2016, 6, 33904. [Google Scholar] [CrossRef] [Green Version]
- Golli-Bennour, E.E.; Bacha, H. Hsp70 expression as biomarkers of oxidative stress: Mycotoxins’ exploration. Toxicology 2011, 287, 1–7. [Google Scholar] [CrossRef]
- Wu, G.; Hu, X.; Ding, J.; Yang, J. Abnormal expression of HSP70 may contribute to PCOS pathology. J. Ovarian Res. 2019, 12, 74. [Google Scholar] [CrossRef]
- Hao, Q.; Zhu, Z.; Xu, D.; Liu, W.; Lu, L.; Li, P. Proteomic characterization of bovine granulosa cells in dominant and subordinate follicles. Hereditas 2019, 156, 21. [Google Scholar] [CrossRef]
- Caloni, F.; Ranzenigo, G.; Cremonesi, F.; Spicer, L.J. Effects of a trichothecene, T-2 toxin, on proliferation and steroid production by porcine granulosa cells. Toxicon 2009, 54, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Yuan, H.; Guo, C.; Lu, Y.; Deng, S.; Yang, Y.; Wei, Q.; Wen, L.; He, Z. Zearalenone induces apoptosis and necrosis in porcine granulosa cells via a caspase-3- and caspase-9-dependent mitochondrial signaling pathway. J. Cell. Physiol. 2012, 227, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Tu, D.; Yuan, L.Y.; Yuan, H.; Wen, L.X. T-2 toxin exposure induces apoptosis in rat ovarian granulosa cells through oxidative stress. Environ. Toxicol. Pharmacol. 2013, 36, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Badinga, L.; Driancourt, M.A.; Savio, J.D.; Wolfenson, D.; Drost, M.; De La Sota, R.L.; Thatcher, W.W. Endocrine and ovarian responses associated with the first-wave dominant follicle in cattle. Biol. Reprod. 1992, 47, 871–883. [Google Scholar] [CrossRef] [Green Version]
- Pizzo, F.; Caloni, F.; Schreiber, N.B.; Cortinovis, C.; Spicer, L.J. In vitro effects of deoxynivalenol and zearalenone major metabolites alone and combined, on cell proliferation, steroid production and gene expression in bovine small-follicle granulosa cells. Toxicon 2016, 109, 70–83. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, L.; Luo, G.; Tang, X.; Ma, L.; Zheng, Y.; Liu, S.A.; Price, C.; Jiang, Z. Arachidonic Acid Regulation of Intracellular Signaling Pathways and Target Gene Expression in Bovine Ovarian Granulosa Cells. Animals 2019, 9, 374. [Google Scholar] [CrossRef] [Green Version]
- Velázquez, M.M.; Salvetti, N.R.; Amweg, A.N.; Díaz, P.U.; Matiller, V.; Ortega, H.H. Changes in the expression of heat shock proteins in ovaries from bovines with cystic ovarian disease induced by ACTH. Res. Vet. Sci. 2013, 95, 1059–1067. [Google Scholar] [CrossRef]
- Lin, P.; Yang, Y.; Li, X.; Chen, F.; Cui, C.; Hu, L.; Li, Q.; Liu, W.; Jin, Y. Endoplasmic reticulum stress is involved in granulosa cell apoptosis during follicular atresia in goat ovaries. Mol. Reprod. Dev. 2012, 79, 423–432. [Google Scholar] [CrossRef]
- Guerin, P.E.; Mouatassim, S.; Menezo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef]
- Alemu, T.W.; Pandey, H.O.; Wondim, D.S.; Gebremedhn, S.; Neuhof, C.; Tholen, E.; Holker, M.; Schellander, K.; Tesfaye, D. Oxidative and endoplasmic reticulum stress defense mechanisms of bovine granulosa cells exposed to heat stress. Theriogenology 2018, 110, 130–141. [Google Scholar] [CrossRef]
- Yanglei, Y.; Fan, Z.; Nan, W.; Huan, L.; Lijun, Y.; Aihua, W.; Yapin, J. Endoplasmic reticulum stress is involved in the t-2 toxin-induced apoptosis in goat endometrium epithelial cells. J. Appl. Toxicol. 2018, 38, 1492–1501. [Google Scholar]
- Quinones, Q.J.; de Ridder, G.G.; Pizzo, S.V. GRP78: A chaperone with diverse roles beyond the endoplasmic reticulum. Histol. Histopathol. 2008, 23, 1409–1416. [Google Scholar] [PubMed]
- Brenjian, S.; Moini, A.; Yamini, N.; Kashani, L.; Faridmojtahedi, M.; Bahramrezaie, M.; Khodarahmian, M.; Amidi, F. Resveratrol Treatment in Patients with Polycystic Ovary Syndrome Decreased Pro-inflammatory and Endoplasmic Reticulum Stress Markers. Am. J. Reprod. Immunol. 2020, 83, e13186. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lin, P.; Chen, F.; Wang, A.; Lan, X.; Song, Y.; Jin, Y. Luman recruiting factor regulates endoplasmic reticulum stress in mouse ovarian granulosa cell apoptosis. Theriogenology 2013, 79, 633–639. [Google Scholar] [CrossRef]
- Wang, H.; Liu, H.; Zheng, Z.M.; Zhang, K.B.; Wang, T.P.; Sribastav, S.S.; Liu, T. Role of death receptor, mitochondrial and endoplasmic reticulum pathways in different stages of degenerative human lumbar disc. Apoptosis 2011, 16, 990. [Google Scholar] [CrossRef]
- Shibue, T.; Taniguchi, T. BH3-only proteins: Integrated control point of apoptosis. Int. J. Cancer 2006, 119, 2036–2043. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HSP Family. | Gene ID | Protein Name | Fold Change | Difference | |
|---|---|---|---|---|---|
| β-Zearalenol | HT-2 | ||||
| HSP10 | HSPE1 | 10 kDa heat shock protein | 2.72 | 2.49 | up |
| HSP27 | HSPB1 | Heat shock 27 kDa protein | 6.79 | 1.58 | up |
| HSP60 | HSPD1 | 60 kDa heat shock protein | 2.17 | 1.83 | up |
| HSP70 | HSPA5/GRP | Endoplasmic reticulum chaperone BiP | 1.90 | 1.82 | up |
| 78 HSPA8 | Heat shock cognate 71 kDa protein | 3.42 | 1.95 | up | |
| HSPA9 | Stress-70 protein | 1.12 | 1.71 | up | |
| HSPA14 | Heat shock 70 kDa protein 14 | 0.30 | 0.63 | down | |
| HSP90 | HSP90AA1 | Heat shock protein HSP 90-alpha | 1.77 | 0.48 | up/down |
| HSP105 | HSPH1 | Heat shock protein 105 kDa | 3.69 | 1.57 | up |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Yang, M.; Li, C.; Yang, F.; Wang, G. Understanding the Toxin Effects of β-Zearalenol and HT-2 on Bovine Granulosa Cells Using iTRAQ-Based Proteomics. Animals 2020, 10, 130. https://doi.org/10.3390/ani10010130
Li L, Yang M, Li C, Yang F, Wang G. Understanding the Toxin Effects of β-Zearalenol and HT-2 on Bovine Granulosa Cells Using iTRAQ-Based Proteomics. Animals. 2020; 10(1):130. https://doi.org/10.3390/ani10010130
Chicago/Turabian StyleLi, Lian, Min Yang, Chengmin Li, Fangxiao Yang, and Genlin Wang. 2020. "Understanding the Toxin Effects of β-Zearalenol and HT-2 on Bovine Granulosa Cells Using iTRAQ-Based Proteomics" Animals 10, no. 1: 130. https://doi.org/10.3390/ani10010130