Simple Summary
Information on the role of cephalopods in the life cycle of elasmobranch tapeworms and other parasites transmitted through the food web is limited. Such knowledge is useful to gain further understanding of the biology and ecology of this group of mollusks and would inform a correct management of wild cephalopod stocks for conservation and fishery purposes. In the present work, we aimed to characterize tapeworms infecting the common octopus Octopus vulgaris, one of the most widely distributed and commercially important cephalopod species, with morphological and molecular methods. Our results suggest a possible important role of O. vulgaris in the transmission of elasmobranch tapeworms and add valuable information on the host-range and distribution of the tapeworms identified.
Abstract
Cephalopods are intermediate/paratenic hosts in the life cycle of elasmobranch tapeworms, nevertheless most records of infection in this group of mollusks are outdated and fragmentary. The present work aimed to investigate the cestode fauna of the common octopus Octopus vulgaris from the Tyrrhenian Sea (Central Mediterranean). The parasitic stages were characterized by light and Scanning Electron Microscopy (SEM) and sequencing of 28S rDNA. Three cestode taxa were identified to the genus level: the onchoproteocephalidean Acanthobothrium sp. (prevalence 28%), the “tetraphyllidean” Anthobothrium sp. (prevalence 13%) and the trypanorhynch Nybelinia sp. (prevalence 3%). The remarkable prevalence observed for gastrointestinal cestodes highlight a possible important role of O. vulgaris in the transmission of elasmobranch tapeworms, particularly Onchoproteocephalideans. Furthermore, the present work provides, for the first time, detailed morphological (SEM) and molecular support to confirm the occurrence of Anthobothrium sp. in cephalopod hosts. In order to gain higher taxonomic resolution for the identified taxa, we stress the need to collect further morphological and molecular data of adult cestodes infecting their elasmobranch definitive hosts.
Keywords:
octopus; cephalopod; parasite; cestode; life cycle; SEM; 28S rDNA; Acanthobothrium; Anthobothrium; Nybelinia 1. Introduction
Cephalopods can host a diversity of larval cestodes, which mainly infect their digestive tract [1,2,3,4] but can also occur free in the mantle cavity or encysted in the mantle musculature [5,6]. Knowledge of the role of cephalopods in the life cycle of elasmobranch tapeworms is limited, since information on the biology and ecology of this group of parasites are still scarce [7,8]. Either eggs or free-swimming coracidia (e.g., in some trypanorhynch species) are ingested by first intermediate hosts (invertebrates) in which procercoids develop; metacestodes are found in cephalopods and other invertebrates, in teleosts and in cetaceans as second intermediate/paratenic hosts and are trophically transmitted to the definitive host represented by elasmobranchs (skates, rays and sharks) in which they develop into adults [9,10].
The numerous reports of infection by elasmobranch tapeworms [11] suggest an important role of cephalopods as intermediate/paratenic hosts in the life cycle of these parasites, nevertheless most of the available information is decades/centuries-old and fragmentary [12]. Particularly, oceanic squids are important hosts in the life cycle of Trypanorhynchs (Nybelinia spp., Tentacularia coryphaenae) and Phyllobothriids (Phyllobothrium spp., Pelichnibothrium spp.) [13,14], while for octopods, records of infection with cestode parasites are scarce [11].
Members of the genus Octopus Cuvier, 1798 host cestodes belonging to the orders Trypanorhyncha, Phyllobothriidea, Onchoproteocephalidea and to the non-monophyletic order “Tetraphyllidea” [1,15,16,17,18,19,20]. With respect to the Mediterranean Sea, the phyllobothridean Phyllobothrium sp. and the trypanorhynchs Nybelinia lingualis (Cuvier, 1817) Dollfus, 1933 and Tetrarhynchus corollatus von Siebold, 1850 (synonymized with N. lingualis, see Reference [10]) have been reported from the common octopus Octopus vulgaris Cuvier, 1797, while the “tetraphyllidean” Scolex pleuronectis Müller, 1788 has been reported from both O. vulgaris and the spider octopus O. salutii Vérany, 1836.
The present work aimed to investigate cestodes infecting O. vulgaris from Tyrrhenian Sea (Southern Italy, Central Mediterranean) through morphological analysis by light microscopy (Leica DMLS, Leica Microsystem, Milan, Italy) and Scanning Electron Microscopy (SEM) (JEOL, Basiglio, Italy) and through molecular analysis of partial 28S rDNA.
2. Materials and Methods
2.1. Sample Collection and Preservation
Fresh dead adult specimens of O. vulgaris (n = 32) were obtained from different areas in the Gulf of Naples (Tyrrhenian Sea, Mediterranean Sea) from local fishermen in the period March 2015–February 2016. Octopuses were dissected and examined for the presence of helminths in the body cavity and in organs of the digestive tract. Collected worms were washed in saline (8‰) and stored in 70% ethanol for further analyses. For permanent mounts, fixed specimens were stained with Semichon’s acetocarmine, destained in 1% acid ethanol, dehydrated in a graded ethanol series and mounted in Canada balsam. Line drawings were made with the help of a camera lucida and all measurements were taken with the imaging software NIS-Elements.
The terminology for larval stages follows Chervy [21] and Palm [10]. The description of tegument surface and microtriches appearance by SEM analysis follows Chervy [22].
2.2. Scanning Electron Microscopy (SEM)
For SEM, specimens were fixed in 2.5% glutaraldehyde (0.1 M sodium cacodylate buffer, pH 7.4), washed in the same buffer and post-fixed in buffered 1% osmium tetroxide, dehydrated through a graded ethanol series, critical point dried and sputter coated with platinum. Observations were made using a JEOL JSM 6700F scanning electron microscope operating at 5.0 kV(JEOL, Basiglio, Italy).
2.3. Molecular Analysis
For molecular analysis, genomic DNA was extracted from eight larvae using the PureLink® Genomic DNA Kit (Life Technologies, Carlsbad, CA, USA) and following the manufacturer’s instructions. The amplification of the D1-D3 region of 28S rDNA was performed with primers U178_f (5′-GCACCCGCTGAAYTTAAG-3′) and L1642_r (5′-CCAGCGCCATCCATTTTCA-3′) [23]. The thermal cycler program (Tpersonal, Biometra) was 40 cycles of 30 sec at 94 °C, 30 sec at 52 °C and 2 min at 72 °C, preceded by a denaturation step at 94 °C for 2 min and followed by an extended elongation step at 72 °C for 10 min. The PCR products were electrophoresed on 1% agarose gel stained with SYBR® Safe DNA Gel Stain (Thermo Fisher Scientific, Carlsbad, CA, USA) in 0.5X TBE. Amplicons were purified by NucleoSpin Gel and PCR Cleanup (Mackerey-Nagel, Düren, Germany, CA, USA) and sequenced with the internal primers 900F (5′-CCGTCTTGAAACACGGACCAAG-3′) and EDC2 (5′-CCTTGGTCCGTGTTTCAAGACGGG-3′) of Lockyer et al. [23], with an ABI 3730 DNA analyzer (StarSEQ, Mainz, Germany). The DNA trace files were assembled with ContigExpress (VectorNTI Advance 11 software, Invitrogen, Carlsbad, California) and the consensus sequences were compared with previously published data by BLAST tools (https://blast.ncbi.nlm.nih.gov/Blast.cgi). Multiple sequences alignments were constructed using BioEdit 7.2.5 [24]. Pairwise distance, using a Kimura 2-parameter model and maximum likelihood (ML) tree (GTR + G + I, bootstrap of 1000 replicates) were obtained by MEGA version X [25]. The sequences generated in this study were deposited in GenBank under the accession numbers MN660283-89 (Acanthobothrium sp.) and MN660290 (Anthobothrium sp.).
3. Results
Cestodes larvae were recovered in 44% of examined octopuses (14 specimens), with intensity values ranging from 1 to 3. Particularly, two larval forms were found in the digestive tract and were assigned to two distinct orders on the basis of their general morphology: 19 Onchoproteocephalidea larvae were recovered from the caecum and intestine of 9 octopuses (prevalence 28%, mean intensity 2.11) and 5 “Tetraphyllidea” larvae were found in the caecum and intestine of 4 octopuses (prevalence 13%, mean intensity 1.25). Furthermore, a single specimen (prevalence 3%), assigned to the Trypanorhyncha, was recovered from the mantle cavity of a single octopus, adhering to the external wall of the crop.
Coinfection of different larval forms was not observed. All the collected larvae were alive, moving vigorously by rapid contractions and extensions of their body. Descriptions and morphometric features in micrometers (µm; mean followed by the range in parentheses) are provided below.
3.1. Morphological Description
3.1.1. Acanthobothrium sp. (Onchoproteocephalidea: Onchobothriidae) plerocercoids
(Measurements based on six specimens). Body elongated, 2053.5 (1146.7–3271) total length × 384.4 (214–652) maximum width, tapering posteriorly (Figure 1a). Posterior to the scolex, a pair of bright red pigmented spots are visible in living specimens (Figure 2a,b). Scolex 351.6 (227–484) long × 445.8 (307.3–773) wide, with large apical sucker and four elongated acetabula in form of bothridia; apical sucker 127.7 (88–172) long × 145 (109–195) wide; bothridia, 277.8 (183.9–363) long × 134.7 (86–178) wide. Bothridia sessile anteriorly, free posteriorly, divided into apical pad and posterior loculus (Figure 3a). Posterior loculus subdivided into three loculi. Hooks not yet developed. Parasite tegument characterized by acicular filitriches and gladiate spinitriches over bothridial surface and neck region (Figure 3b); capilliform filitriches covering entire larval body including caudal region (Figure 3c,d).
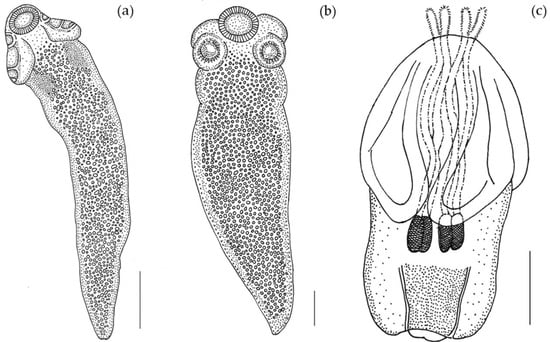
Figure 1.
Line drawing of larvae from O. vulgaris. (a) larva of Acanthobothrium (Scale bar: 300 µm); (b) larva of Anthobothrium (Scale bar: 140 µm); (c) larva of Nybelinia (Scale bar: 500 µm).
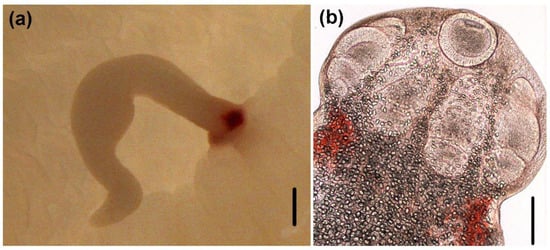
Figure 2.
Light micrographs of larvae of Acanthobothrium. (a) attached to the intestinal lumen of O. vulgaris (Scale bar: 500 µm; (b) scolex (Scale bar: 100 µm).
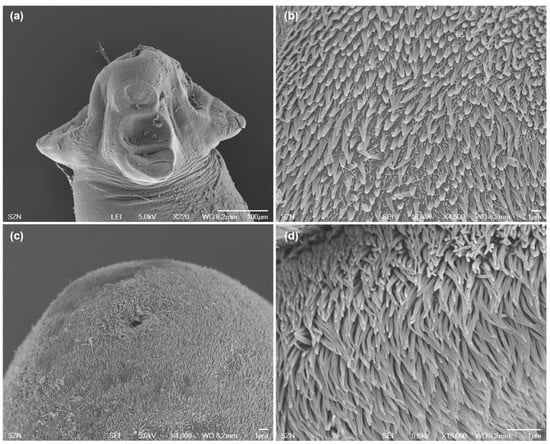
Figure 3.
Scanning electron micrographs of larvae of Acanthobothrium from O. vulgaris. (a) scolex, lateral view; (b) detail of bothridial surface with capilliform filitriches and gladiate spinitriches; (c) caudal end, sub-apical view; (d) detail of surface of the caudal end with capilliform filitriches.
3.1.2. Anthobothrium sp. (“Tetraphyllidea”: Clade 2) plerocercoids
(Measurements based on two specimens). Body elongated, brownish, 1029 (1007–1051) total length × 344 (315–373) maximum width, tapering posteriorly (Figure 1b). Scolex 196 (181–211) long × 404 (392–416) wide, with apical sucker and four sessile acetabula (Figure 4a). Apical sucker 130 (112–148) long × 110 (102–118) wide, acetabula in the form of suckers 157 (151–163) long × 131 (127–135) wide. Larval body and scolex covered with capilliform filitriches (Figure 4b–d).
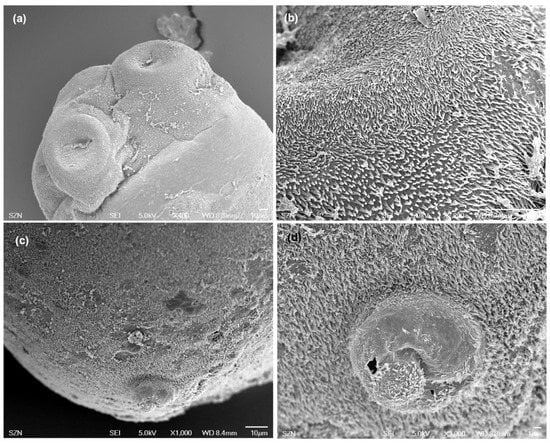
Figure 4.
Scanning electron micrographs of larvae of Anthobothrium. (a) scolex, lateral view; (b) detail of surface of acetabula showing capilliform filitriches on distal acetabular surface and gladiate spinitriches on proximal bothridial surfaces; (c) caudal end, sub-apical view; (d) detail of caudal end.
3.1.3. Nybelinia sp. (Trypanorhyncha: Tentaculariidae) plerocercoid
(Measurements based on a single specimen). Body pyriform (Figure 1c), 2269 μm total length × 1000 μm maximum width. Scolex 1220 μm wide, with two bothria; bothria 1370 μm long × 806 μm wide (Figure 5a). Prebulbar organs absent, tentacular bulbs 484 μm long. Appendix 622 μm long. Velum 556 μm long. Presence of four slender cylindrical tentacles 237 μm long (everted part) × 39 μm maximum width, slightly attenuated at the ends. Armature homeoacanthous, homeomorphous (Figure 5b,c). Metabasal hooks uncinate 15 μm long × 12 μm wide, arranged in diagonal spiral rows. Surface of bothria (Figure 5d) covered with filitriches and lineate spinitriches (Figure 5e), bothrial border covered with longer lineate spinitriches (Figure 5f).

Figure 5.
Scanning electron micrographs of larvae of Nybelinia. (a) scolex with partially everted tentacles; (b) detail of the tentacles; (c) metabasal armature of tentacle with uncinate hooks arranged in diagonal spiral rows; (d) detail of distal bothrial surface; (e) detail of capilliform filitriches and digitiform spinitriches on distal bothrial surface; (f) detail of bothrial border with longer digitiform spinitriches.
3.2. Molecular Results
The 28S rDNA of 8 specimens, 7 Acanthobothrium sp. and 1 Anthobothrium sp. were successfully sequenced and were 1763–1799 bp in length, with a mean divergence 0–0.12% among them. The BLAST analysis of the 7 specimens identified morphologically as Acanthobothrium sp. indicated the highest identity (98%) with Acanthobothrium santarosaliense Caira & Zahner, 2001 (KF685751) adults infecting the Mexican hornshark Heterodontus mexicanus Taylor & Castro-Aguirre, 1972 fished off Santa Rosalia (Gulf of California) and A. wedli Robinson, 1959 (MH913270) adults infecting the rough skate Zearaja nasuta (Müller & Henle, 1841) off New Zealand, while the single Anthobothrium sp. analyzed was 100% (69% coverage) identical with Anthobothrium sp. (GQ470168) larva infecting the largehead hairtail Trichiurus lepturus Linnaeus, 1758 from the Gulf of Mexico. The multiple alignment of the 7 Acanthobothrium showed they were all identical to each other except for one specimen showing some transition and transversion. The Maximum-Likelihood tree (Figure 6) generated aligning our sequences with species of Acanthobothrium spp. and Anthobothrium spp. 28S rDNA retrieved from GenBank showed our Acanthobothrium specimens forming a monophyletic group separated but closely related and well supported with the species A. santarosaliense, A. wedli and Acanthobothrium sp. Anthobothrium from the present study is included in the cluster of the Anthobothrium sp. (mean distance 0–0.07%).
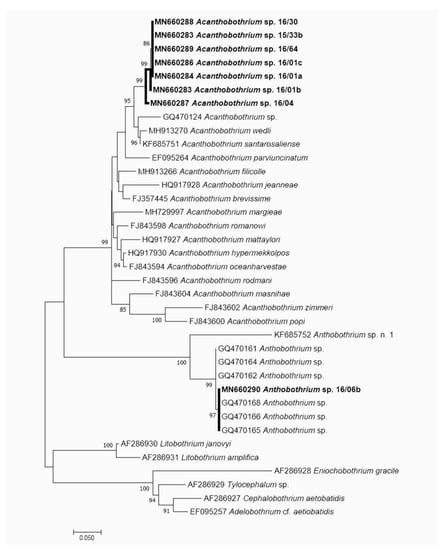
Figure 6.
Maximum Likelihood tree (GTR + G + I) showing the relationship with Acanthobothrium sp. and Anthobothrium sp. described in the present research. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. The details of the sequences used to build the maximum likelihood (ML) tree are reported in the Supplementary Table S1.
4. Discussion
Understanding the diversity and life cycle of elasmobranch tapeworms is often problematic due to difficulties in the morphological identification of larval forms [9] and in performing experimental life-cycle studies, insufficient information available in the literature and the frequent reporting of currently unaccepted larval names (e.g., Scolex pleuronectis) in the intermediate hosts.
For several orders of elasmobranch tapeworms (e.g., “tetraphyllideans”), the identification of larvae is particularly troublesome, since their scoleces remain less differentiated until they develop in the definitive host, therefore the use of molecular data is often required in order to clarify their taxonomy [8].
In a study aimed at generating a molecular sequence library for a wide range of larval cestodes, in order to elucidate morphological features useful for their identification [8], Acanthobothrium and Anthobothrium larvae infecting teleosts from the Gulf of Mexico have been characterized with morphological and morphometric analyses and through amplification and sequencing of partial 28S rDNA gene. Their size range and general morphology are in accordance with our description of larval Acanthobothrium and Anthobothrium from Mediterranean O. vulgaris.
Plerocercoids tentatively assigned to the genus Acanthobothrium based on the morphology of the bothridia were recovered from the intestine of O. vulgaris collected in Roscoff (Northeastern Atlantic, France) [26]. Similar larvae, reported as Scolex polymorphus, were found infecting the digestive tract of Sepia officinalis Linnaeus, 1758 and O. vulgaris collected in the Bay of Arcachon (Northeastern Atlantic, France) [27]. Larvae described by Dollfus [26,27] are similar to the larvae of Acanthobothrium described in the present study with respect to their general morphology and the presence of bright red regions in the anterior part of the body, posteriorly to the scolex. However, their bothridia differ in the number of septa dividing the loculus posterior to the accessory sucker: bothridia of larvae described by Dollfus are provided with an anterior accessory sucker and a posterior loculus divided in two parts by one transversal septum, while in our Acanthobothrium larvae the posterior loculus is divided in three parts by two transversal septa. In larvae of Acanthobothrium identified with molecular methods the number of visible septa in the posterior loculus of bothridia can vary from one to two [8]; based on this evidence we speculate that our Acanthobothrium larvae and those described by Dollfus [26,27] in O. vulgaris from northeastern Atlantic may belong to the same species of Acanthobothrium. Possible routes of infection with Acanthobothrium in O. vulgaris may be represented by bivalve and gastropod mollusks and decapod crustaceans, that are part of the diet of this octopus species and in which different Acanthobothrium spp. have been repeatedly reported [28,29,30].
With 201 Acanthobothrium species described so far [31], this genus is the most speciose in the order Onchoproteocephalidea and among tapeworms parasitizing elasmobranchs [32] and includes a majority of species that parasitize batoids and some that parasitize sharks [31,33]. Unfortunately, sequence data are currently available for only 16 Acanthobothrium species, therefore the identification to the species level of larvae belonging to this genus remains problematic. Our samples apparently belong either to not yet molecularly characterized species of Acanthobothrium or to a new species.
Several Acanthobothrium species have been reported in elasmobranchs from the Mediterranean Sea [34,35,36]. Particularly, for the Tyrrhenian Sea most of the records are quite old—A. benedeni Lönnberg, 1889 in the pelagic stingray Pteroplatytrygon violacea (Bonaparte, 1832); A. coronatum (Rudolphi, 1819) Blanchard, 1848 in the dogfish Scyliorhinus stellaris (Linnaeus, 1758) and S. canicula (Linnaeus, 1758), in the common smooth-hound Mustelus mustelus (Linnaeus, 1758), in the picked dogfish Squalus acanthias Linnaeus, 1758, the angelshark Squatina squatina (Linnaeus, 1758) and the common torpedo Torpedo torpedo (Linnaeus, 1758); A. crassicolle Wedl, 1855 in an unidentified species of Raja Linnaeus, 1758; A. filicolle (Zschokke, 1887) Yamaguti, 1959 in P. violacea, in the Marbled electric ray Torpedo marmorata Risso, 1810 and in T. torpedo; A. musculosum (Baer, 1948) Yamaguti, 1959 in P. violacea and A. zschokkei Baer, 1948 in T. torpedo.
The genus Anthobothrium is currently classified in the non-monophyletic order “Tetraphyllidea” and has not been assigned to a family; the most recent classification scheme assigns this genus to the Clade 2 of the “Tetraphyllidea” [33]. The taxonomy of the “Tetraphyllidea” have more recently been revised in the light of new molecular data, with the creation of new eucestode orders—namely Rhinebothriidea [37], Onchoproteocephalidea and Phyllobothriidea [38]—while other species remain in the non-monophyletic “Tetraphyllidea” until additional molecular data become available [33]. With respect to previous work carried out in cephalopods, the description of our Anthobothrium sp. larva is consistent with the description of Scolex sp. infecting Loligo vulgaris Lamarck, 1798 from Brest, France [26] and from the Bay of Arcachon [27]. Nevertheless, morphological data alone do not allow to assign these larval stages to a genus, since larvae with unilocular bothridia could be tentatively assigned to genera belonging to different cestode orders [8,39]. To the authors’ knowledge, no confirmed report of Anthobothrium in cephalopod hosts was so far available in the literature.
Similarly to Acanthobothrium, the identification of our Anthobothrium larvae to the species level is hampered by the scarcity of molecular information: this genus currently includes 16 valid species [33], however sequence data of only two species (A. laciniatum, A. caseyi) are available.
In the Tyrrhenian Sea, two species of Anthobothrium are reported in different elasmobranchs: A. auriculatum (Rudolphi, 1819) in T. torpedo [40], in T. marmorata and in S. squatina [41,42,43] and A. cornucopia Van Beneden, 1850 in S. squatina [43]. The diet of these host species consists of a variety of vertebrate and invertebrate preys, including cephalopods [44,45]. A few additional Anthobothrium species are reported from different parts of the Mediterranean Sea (Supplementary Table S1).
Among Trypanorhyncha, the species Nybelinia lingualis and Tentacularia coryphaenae Bosc, 1802 are the most commonly encountered in cephalopods from Mediterranean and extra-Mediterranean areas. Particularly, N. lingualis has been reported in O. vulgaris from the Mediterranean Sea and English Channel [1,18,19,20,46,47,48]. Scolex features, body size and general morphology of the Nybelinia specimen characterized in the present study are consistent with the description of N. lingualis reported in O. vulgaris and Todaropsis eblanae (Ball, 1841) from NE Atlantic [1,27,49]; particularly, detailed observations by SEM allowed to observe the form and arrangement of hooks over the tentacles and the appearance and distribution of microtriches, adding further detailed information on the microthrix pattern over bothrial surface. Nevertheless, due to the scarcity of material available (only one specimen) and to the lack of molecular data it is not possible to confirm the identity of the collected larva to the species level.
5. Conclusions
Our results suggest a possible important role of O. vulgaris in the transmission of elasmobranch tapeworms, particularly Onchoproteocephalideans. With respect to tripanorhynchs, this role is arguably less important as compared to ommastrephid squids, where prevalence can be higher than 90% and more than 200 larvae can be found in a single squid [14].
Furthermore, the present work provides, for the first time, detailed morphological (SEM) and molecular support to confirm the occurrence of Anthobothrium sp. in cephalopod hosts and the presence of Acanthobothrium sp. infection in O. vulgaris from the Mediterranean Sea.
Supplementary Materials
The following are available online at https://www.mdpi.com/2076-2615/10/11/2038/s1, Table S1: List of the sequences used for build the ML tree, with details of the developmental stage, host, locality and specific references.
Author Contributions
Conceptualization, P.T. and G.F.; Methodology, P.T., M.C. and A.G.; Investigation, P.T.; Resources, G.F.; Data Curation, P.T.; Writing–Original Draft Preparation, P.T.; Writing–Review & Editing, P.T., M.C., A.G., M.L.F.; Supervision, M.L.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors would like to thank F. Iamunno and R. Graziano (Electron Microscopy Service, Stazione Zoologica Anton Dohrn, Naples, Italy) for their technical assistance. This study was supported by RITMARE (Italian research for the Sea) Flagship project funded by the Italian Ministry of University and Research (MIUR) and by Stazione Zoologica Anton Dohrn.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gestal, C.; Abollo, E.; Arias, C.; Pascual, S.S.E.M. study of the plerocercoid larval Phyllobothrium sp. (Tetraphyllidea, Phyllobothriidae) and Nybelinia lingualis (Trypanothyncha, Tentaculariidae), cestode parasite in Octopus vulgaris (Mollusca, Cephalopoda) off Vigo estuary. Iberus 1998, 16, 125–132. [Google Scholar]
- Brickle, P.; Olson, P.D.; Littlewood, D.T.J.; Bishop, A.; Arkhipkin, A.I. Parasites of Loligo gahi from waters off the Falkland Islands, with a phylogenetically based identification of their cestode larvae. Can J. Zool. 2001, 79, 2289–2296. [Google Scholar] [CrossRef]
- Shukhgalter, O.A.; Nigmatullin, C.M. Parasitic helminths of jumbo squid Dosidicus gigas (Cephalopoda: Ommastrephidae) in open waters of the central east Pacific. Fish Res. 2001, 54, 95–110. [Google Scholar] [CrossRef]
- Pardo-Gandarillas, M.C.; Lohrmann, K.B.; Valdivia, A.L.; Ibáñez, C.M. First record of parasites of Dosidicus gigas (d’Orbigny, 1835) (Cephalopoda: Ommastrephidae) from the Humboldt Current system off Chile. Rev. Biol. Mar. Oceanogr 2009, 44, 397–408. [Google Scholar]
- Naidenova, N.N.; Nigmatullin, C.M.; Gaevskaya, A.V. The helminthofauna and parasite-host relationships of the squid Sthenoteuthis oualaniensis in the Indian Ocean and Red Sea. In Proceedings of the Symposium on Parasitology and Pathology of Marine Organisms, Leningrad, Russia, 13–16 October 1981; pp. 69–74. (In Russian). [Google Scholar]
- Naidenova, N.N.; Nigmatullin, C.M.; Gaevskaya, A.V. The helminth fauna and host-parasite relations of squids Sthenoteuthis oualaniensis (Lesson) (Cephalopoda, Ommastrephidae) in the Indian Ocean and the Red Sea. In Parasitology and Pathology of Marine Organisms of the World Ocean; Hargis, W.J., Ed.; NOAA Scientific Publications Office: Seattle, WA, USA, 1985; Volume 25, pp. 113–116. [Google Scholar]
- Williams, H.; Jones, A. Parasitic Worms of Fish; Taylor & Francis: London, UK, 1994. [Google Scholar]
- Jensen, K.; Bullard, S.A. Characterization of a diversity of tetraphyllidean and rhinebothriidean cestode larval types, with comments on host associations and life-cycles. Int. J. Parasitol. 2010, 40, 889–910. [Google Scholar] [CrossRef] [PubMed]
- Caira, J.N.; Reyda, F.B. Eucestoda (true tapeworms). In Marine Parasitology; Rhode, K., Ed.; CABI Publishing: New York, NY, USA, 2005; pp. 92–104. [Google Scholar]
- Palm, H.W. The Trypanorhyncha Diesing, 1863; PKSPL-IPB Press: Bogor, Indonesia, 2004. [Google Scholar]
- Tedesco, P.; Bevilacqua, S.; Fiorito, G.; Terlizzi, A. Global patterns of parasite diversity in cephalopods. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pascual, S.; Abollo, E.; Mladineo, I.; Gestal, C. Metazoa and Related Diseases. In Handbook of Pathogens and Diseases in Cephalopods; Gestal, C., Pascual, S., Guerra, A., Fiorito, G., Vieites, J.M., Eds.; Springer: Cham, Switzerland, 2019; pp. 169–179. [Google Scholar]
- Ruhnke, T.R. Tapeworms of elasmobranchs (Part III) A Monograph on the Phyllobothriidae (Platyhelminthes, Cestoda). Bull Univ. Nebr. State Mus. 2011, 25, 1–208. [Google Scholar]
- Hochberg, F.G. Diseases of Mollusca Cephalopoda-Diseases caused by Protistans and Metazoans. In Diseases of Marine Animals; Kinne, O., Ed.; Biologische Anstalt Helgoland: Hamburg, Germany, 1990; Volume 3, pp. 47–227. [Google Scholar]
- Riser, N.W. Studies on the Tetraphyllidea. Ph.D. Thesis, Stanford University, Palo Alto, CA, USA, 1949. [Google Scholar]
- Cake, E.W. A key to larval cestodes of shallow-water, benthic mollusks of the northern Gulf of Mexico. Proc. Helminthol. Soc. Wash. 1976, 43, 160–171. [Google Scholar]
- González, A.F.; Pascual, S.; Gestal, C.; Abollo, E.; Guerra, A. What makes a cephalopod a suitable host for a parasite? The case of Galician waters. Fish. Res. 2003, 60, 177–183. [Google Scholar] [CrossRef]
- Dollfus, R.P. Cestodes des invertébrés marins et thalassoides. In Faune de France: Cestodes; Joyeux, C., Baer, J.G., Eds.; Paul Lechevalier et Fils: Paris, France, 1936; Volume 30, pp. 509–539. [Google Scholar]
- Dollfus, R.P. Copépodes, Isopodes et Helminths parasites de Cephalopodes de la Méditerranée et de l’Atlantique Européen. Faune Mar Pyrénées-Orientales 1958, 1, 61–72. [Google Scholar]
- Dollfus, R.P. Enumération des cestodes du plancton et des invertébrés marins. (6econtribution). Ann. Parasit. Hum. Comp. 1964, 39, 329–379. [Google Scholar] [CrossRef]
- Chervy, L. The terminology of larval cestodes or metacestodes. Syst. Parasitol. 2002, 52, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Chervy, L. Unified terminology for cestode microtriches: A proposal from the International Workshops on Cestode Systematics in 2002–2008. Folia. Parasit. 2009, 56, 199–230. [Google Scholar] [CrossRef] [PubMed]
- Lockyer, A.E.; Olson, P.D.; Littlewood, D.T.J. Utility of complete large and small subunit rRNA genes in resolving the phylogeny of the Neodermata (Platyhelminthes): Implications and a review of the cercomer theory. Biol. J. Linn. Soc. 2003, 78, 155–171. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT; Information Retrieval Ltd.: London, UK, 1999; Volume 41, pp. 5–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 547–1549. [Google Scholar] [CrossRef]
- Dollfus, R.P. Énumération des cestodes du plancton et des invertébrés marins. II. Mollusques céphalopodes et Crustacés. Ann. Parasit. Hum. Comp. 1923, 1, 363–394. [Google Scholar] [CrossRef]
- Dollfus, R.P. Addendum à mon «Énumération des cestodes du plancton et des invertébrés marins». Ann. Parasit. Hum. Comp. 1929, 7, 325–347. [Google Scholar] [CrossRef]
- Young, R.T. Cestodes of sharks and rays in Southern California. Proc. Helminthol. Soc. Wash. 1954, 21, 106–112. [Google Scholar]
- Goldstein, R.J. The genus Acanthobothrium van Beneden, 1849 (Cestoda: Tetraphyllidea). J. Parasitol. 1967, 53, 455–483. [Google Scholar] [CrossRef]
- Holland, N.D.; Wilson, N.G. Molecular identification of larvae of a tetraphyllidean tapeworm (Platyhelminthes: Eucestoda) in a razor clam as an alternative intermediate host in the life cycle of Acanthobothrium brevissime. J. Parasitol. 2009, 95, 1215–1218. [Google Scholar] [CrossRef]
- Zaragoza-Tapia, F.; Pulido-Flores, G.; Gardner, S.L.; Monks, S. Host relationships and geographic distribution of species of Acanthobothrium Blanchard, 1848 (Onchoproteocephalidea, Onchobothriidae) in elasmobranchs: A metadata analysis. ZooKeys 2020, 940, 1–49. [Google Scholar] [CrossRef] [PubMed]
- Randhawa, H.S.; Poulin, R. Evolution of interspecific variation in size of attachment structures in the large tapeworm genus Acanthobothrium (Tetraphyllidea: Onchobothriidae). Parasitology 2010, 137, 1707–1720. [Google Scholar] [CrossRef] [PubMed]
- Caira, J.N.; Jensen, K. Planetary Biodiversity Inventory (2008–2017): Tapeworms from Vertebrate Bowels of the Earth; Natural History Museum, The University of Kansas: Lawrence, KS, USA, 2017. [Google Scholar]
- Zschokke, F. Recherches sur la structure anatomique des cestodes. Mem. Inst. Nat. Genevois. 1888, 17, 1–396. [Google Scholar]
- Baer, J.G. Contributions à l’étude des cestodes de sélaciens. I-IV. Bull. Soc. Neuchâtel Sci. Nat. 1948, 71, 63–122. [Google Scholar]
- Yamaguti, S. Systema Helminthum. vol. II. The Cestodes of Vertebrates; Interscience Publishers: New York, NY, USA, 1959. [Google Scholar]
- Healy, C.J.; Caira, J.N.; Jensen, K.; Webster, B.L.; Littlewood, D.T.J. Proposal for a new tapeworm order, Rhinebothriidea. Int. J. Parasitol. 2009, 39, 497–511. [Google Scholar] [CrossRef]
- Caira, J.N.; Jensen, K.; Waeschenbach, A.; Olson, P.D.; Littlewood, D.T.J. Orders out of chaos–molecular phylogenetics reveals the complexity of shark and stingray tapeworm relationships. Int. J. Parasitol. 2014, 44, 55–73. [Google Scholar] [CrossRef] [PubMed]
- Avdeeva, N.V.; Avdeev, V.V. Peculiarities of morphogenesis of adhesive organs of some plerocercoids of the collective genus Scolex (Tetraphyllidea) and their identification. Parazitologiya 1980, 14, 242–250. [Google Scholar]
- Di Cave, D.; Orecchia, P.; Ortis, M.; Paggi, L. Metazoi parassiti di alcuni elasmobranchi del mar Tirreno. Biol. Mar. Mediterr. 2003, 10, 249–252. [Google Scholar]
- Orecchia, P.; Paggi, L.; Minervini, R.; Di Cave, D. La parassitofauna delle specie ittiche strascicabili presenti alla foce del fiume Tevere. Oebalia 1985, 11, 623–632. [Google Scholar]
- Bona, F.; Buriola, E.; Cerioni, S.; di Cave, D.; Orecchia, P.; Paggi, L. Cestoda, Monogenea. In Checklist Delle Specie Della Fauna Italiana; Minelli, A., Ruffo, S., La Posta, S., Eds.; Calderini: Bologna, Italy, 1995. [Google Scholar]
- Paggi, L. Cestoda. Biol. Mar. Mediterr. 2008, 15 (Suppl. 1), 150–154. [Google Scholar]
- Capapé, C.; Crouzet, S.; Clement, C.; Vergne, Y.; Guelorget, O. Diet of the marbled electric ray Torpedo marmorata (Chondrichthyes: Torpedinidae) off the Languedocian coast (Southern France, Northern Mediterranean). Ann. Ser. Hist. Nat. 2007, 17, 17–22. [Google Scholar]
- Miller, M.H. Status Review Report of 3 Species of Angelsharks: Squatina aculeata, S. oculata and S. squatina. Report to National Marine Fisheries Service, Office of Protected Resources; 2016; pp. 6–74. Available online: repository.library.noaa.gov/view/noaa/16283 (accessed on 1 September 2020).
- Vaullegeard, A.C. Recherches sur les tétrarhynqués. Mém. Soc. Linn. Normandie 1899, 19, 187–376. [Google Scholar]
- Vaullegeard, A.C. Sur les Tétrarhynqués de la collection helminthologique du Professeur Corrado Parona, de Gênes. Atti. Soc. Ligust. Sci. Nat. Geogr. 1901, 12, 43–49. [Google Scholar]
- Pintner, T. Wenig Bekanntes und Unbekanntes von Rüssel bandwürmer. Sitzungsber Akademie d Wissenschaften Wien Math–Naturw Classe 1 1930, 139, 445–537. [Google Scholar]
- Pascual, S.; Arias, C.; Guerra, Á. First record of the trypanorhynchidean parasite Nybelinia lingualis Cuvier, 1817 in the squid Todaropsis eblanae (Cephalopoda: Ommastrephidae). Sci. Mar. 1996, 60, 553–555. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).