Simple Summary
The emergence of antimicrobial resistance (AMR) has become an important consideration in animal health, including marine mammals, and several potential zoonotic AMR bacterial strains have been isolated from wild cetacean species. Although the emergence of AMR bacteria can be assumed to be much more plausible in captive than in free-ranging cetaceans owing to their frequent contact with humans and antibiotic treatments, the spread and its impacts of AMR bacteria in captive animals have not been adequately investigated yet. Here in this study, we present evidence on the presence of multidrug-resistant potential zoonotic bacteria which caused fatal infection in a captive dolphin bred at a dolphinarium in South Korea.
Abstract
The emergence of antimicrobial resistant (AMR) strains of Morganella morganii is increasingly being recognized. Recently, we reported a fatal M. morganii infection in a captive bottlenose dolphin (Tursiops truncatus) bred at a dolphinarium in South Korea. According to our subsequent investigations, the isolated M. morganii strain KC-Tt-01 exhibited extensive resistance to third-generation cephalosporins which have not been reported in animals. Therefore, in the present study, the genome of strain KC-Tt-01 was sequenced, and putative virulence and AMR genes were investigated. The strain had virulence and AMR genes similar to those of other M. morganii strains, including a strain that causes human sepsis. An amino-acid substitution detected at the 86th residue (Arg to Cys) of the protein encoded by ampR might explain the extended resistance to third-generation cephalosporins. These results indicate that the AMR M. morganii strain isolated from the captive dolphin has the potential to cause fatal zoonotic infections with antibiotic treatment failure due to extended drug resistance, and therefore, the management of antibiotic use and monitoring of the emergence of AMR bacteria are urgently needed in captive cetaceans for their health and conservation.
1. Introduction
The spread of antimicrobial resistance genes (ARGs) and their acquisition in potential zoonotic bacterial pathogens have been recognized as serious threats to human and animal health [1]. As in other animals, the emergence of antimicrobial resistant (AMR) bacteria in cetaceans has become an important consideration [2]. Several potential zoonotic bacterial species have been isolated from wild cetacean species inhabiting the United States [3] and Brazil [4]. Moreover, the emergence of AMR bacteria can be assumed to be much more plausible in captive than in free-ranging marine mammals owing to their frequent contact with humans and antibiotics used for prophylactic or curative treatment. Indeed, AMR bacteria, including those causing fatal infections, have been reported in captive dolphins bred in a dolphinarium [5,6,7,8]. However, the mechanisms of AMR in captive marine mammals have not been adequately investigated because of limited genetic (or genomic) information. Therefore, there is an urgent need to investigate the genomic characteristics of AMR bacteria isolated from captive cetaceans to evaluate the current status of AMR in dolphinariums and to clarify the potential health risks for marine mammals and the humans they encounter.
Morganella morganii is a motile flagellated, straight, rod-shaped, Gram-negative bacterium that is ubiquitous in the environment and intestinal tracts of humans and animals. M. morganii has clinical significance as a potential causal pathogen of nosocomial and animal infections [9]. Similar to other Enterobacteriaceae species, M. morganii shows intrinsic resistance to β-lactam antibiotics, including first- and second-generation cephalosporins. In addition, the rates of drug resistance and AMR genes in this bacterium have recently increased [9]. Moreover, this bacterium has been implicated in the notorious histamine fish poisoning (or scombroid poisoning) based on the sporadic presence of histidine decarboxylase (hdc) [10]. Recently, we reported the occurrence of a fatal M. morganii infection in a captive bottlenose dolphin (Tursiops truncatus) bred at a dolphinarium in South Korea [11]. In subsequent investigations of the isolated M. morganii strain KC-Tt-01, we found that it exhibited intrinsic resistance against several antibiotics and was extensively resistant to third-generation cephalosporins. Therefore, in the present study, we characterized the genome of M. morganii strain KC-Tt-01 and its relation to the phenotypic AMR profile.
2. Materials and Methods
2.1. Ethics Statement
The dolphins mentioned in this work were cared (or managed) in a captive environment and handled according to Korean law (Act on the management of zoos and aquariums, Act 14227/2016); all the samples obtained (in vivo diagnostic swabs and blood, and post-mortem samples) were collected according to the above and within Korean law (Wildlife protection and management act, Act 13882/2016), which establishes the management objectives and prescriptions to maintain the species under human care. The animal study was reviewed and approved by the ethics and welfare committee (Approved number: 2017-Animal Experiment-15) in the National Institute of Fisheries Science, Ministry of Oceans and Fisheries, Republic of Korea.
2.2. Origin of Clinical Isolate M. morganii Strain KC-Tt-01
The M. morganii strain KC-Tt-01, which caused fatal fibrino-hemorrhagic bronchopneumonia, was originally isolated from the pericardial fluid of a captive female bottlenose dolphin (T. truncatus) bred at a dolphinarium in South Korea [11]. Strain KC-Tt-01 was stored in tryptic soy broth (Difco, Detroit, MI, USA) with 10% glycerol at −80 °C until use.
2.3. Determination of Phenotypic Antibiotic Resistance of M. morganii Strain KC-Tt-01
Antimicrobial susceptibility of strain KC-Tt-01 was tested using the disk diffusion method according to the guidelines of the Clinical and Laboratory Standards Institute [12]. A total of 13 groups of antimicrobial agents were used: penicillins, β-lactam/β-lactamase inhibitor combinations, cephems (including first-, second-, third-, and fourth-generation cephalosporins), monobactams, carbapenems, aminoglycosides, tetracyclines, fluoroquinolones, quinolones, folate pathway inhibitors, macrolides, phenicols, and polymyxin (Table 1). The minimum inhibitory concentrations (MICs) of the selected antimicrobial agents were determined using MIC evaluator strips (Oxoid Ltd., Basingstoke, UK). For quality control, Escherichia coli ATCC25922 and ATCC35218 were used for the analysis.

Table 1.
Antibiotic resistance profile of Morganella morganii strain KC-Tt-01 †.
2.4. Sequencing and Analysis of the M. morganii Strain KC-Tt-01 Genome
M. morganii strain KC-Tt-01 was cultured overnight on 5% sheep blood agar (Hanil Komed, Seongnam, Korea) at 37 °C. Bacterial genomic DNA was isolated using a DNeasy blood and tissue kit (QIAGEN, Hilden, Germany) following the manufacturer’s protocol. Genome sequencing was carried out by Macrogen Inc. (Seoul, Korea) using a hybrid approach with the PacBio RS II system (Pacific Biosciences, USA) and the HiSeq 2000 system (Illumina, San Diego, CA, USA). The generated sequences (1,242,336,868 bp; 135,696 reads) were assembled using HGAP v.3.0 (https://github.com/PacificBiosciences/Bioinformatics-Training/wiki/HGAP), and the Illumina paired-end reads (965,969,557 bp, 9,572,268 reads) were mapped to the assembled contigs to improve the accuracy of the sequenced genome. Genome annotation was conducted using the National Center of Biotechnology Information Prokaryotic Genome Annotation Pipeline (http://www.ncbi.nlm.nih.gov/books/NBK174280/), and PHASTER (http://phaster.ca/) analysis was used to detect prophages. To assess the genomic relatedness to other Morganella species, the average nucleotide identity was analyzed using OrthoANI (http://www.ezbiocloud.net/tools/orthoani). Putative virulence and antimicrobial resistance genes were preliminarily screened by searching against the Virulence Factor (http://www.mgc.ac.cn/VFs/) and ARG-ANNOT (http://en.mediterranee-infection.com/article.php?laref=283&titre=arg-annot-) databases, respectively, and were ultimately identified by manual comparisons with those reported for other M. morganii strains in GenBank, including strain KT, which causes human sepsis [15].
2.5. Culture Deposition and Nucleotide Sequence Accession No.
M. morganii strain KC-Tt-01 was deposited in the Korean Culture Center of Microorganisms (KCCM) as KCCM 90280. The complete genome sequence of the strain has been deposited in GenBank under accession number CP025933.
3. Results and Discussion
The antimicrobial susceptibility profile of M. morganii strain KC-Tt-01 is shown in Table 1. The strain was resistant to ampicillin (MIC >256 μg/mL), amoxicillin-clavulanate (>256 μg/mL), ampicillin-sulbactam, cephalothin, cephazolin, cefoxitin, cefuroxime, cefotaxime (64 μg/mL), ceftazidime, aztreonam, erythromycin (>256 μg/mL), and polymyxin B. The intrinsic resistance of M. morganii against ampicillin, amoxicillin-clavulanate, first- and second-generation cephalosporins, tetracycline, erythromycin, and polymyxin B has been well documented [12,13,14], and similar results were obtained for strain KC-Tt-01 in this study. However, our isolate was also resistant to cefotaxime, a third-generation cephalosporin. Therefore, we sequenced the genome of strain KC-Tt-01 to determine its resistance mechanism and provide genomic insights into this potential zoonotic pathogen infecting marine mammal species.
The fully assembled complete genome of strain KC-Tt-01 was 3,824,890 bp long (G + C content, 51.1%), and plasmids were not detected. The annotated genome included 3,611 genes, 3,506 coding sequences, 22 rRNAs (5S, 16S, and 23S), 79 tRNAs, and four non-coding RNAs. In addition, five prophage regions (three intact and two incomplete) were detected (Supplementary Table S1). The KC-Tt-01 genome showed the highest similarity to M. morganii strain FDAARGOS_63 (CP026046, 98.94%) based on OrthoANI analysis, and clustered with other M. morganii strains in a phylogenetic tree (Figure 1). Most of the potential virulence genes in KC-Tt-01 were very similar to those in strain KT, including the histidine decarboxylase gene cluster (hdcT1, hdc, hdcT2, and hisRS); however, repeats-in-toxin (RTX) toxin hemolysin A was detected only in our isolate (Supplementary Table S2). Moreover, the genome included genes known to be involved in resistance to β-lactams (ampD, ampH, ampR, dha-4, and emrAB), phenicols (catA2), and polymyxin (arnA), which were very similar to those identified in other M. morganii strains included in GenBank (Supplementary Table S3). These results support the intrinsic resistance of M. morganii strain KC-Tt-01 to β-lactam antibiotics, including first- and second-generation cephalosporins and polymyxin B. Although the emergence of third-generation cephalosporin-resistant M. morganii has been reported, all previously reported strains originated from nosocomial infections, and TEM β-lactamase production was implicated in the resistance mechanism (e.g., TEM-10 and TEM-21) [16,17]. However, no such β-lactamase was found in strain KC-Tt-01, whereas we detected a substitution at the 86th amino acid (Arg to Cys) of the protein encoded by ampR only in strain KC-Tt-01. This alteration might be associated with the extended resistance to third-generation cephalosporins, as has been reported in other Enterobacteriaceae species [18].
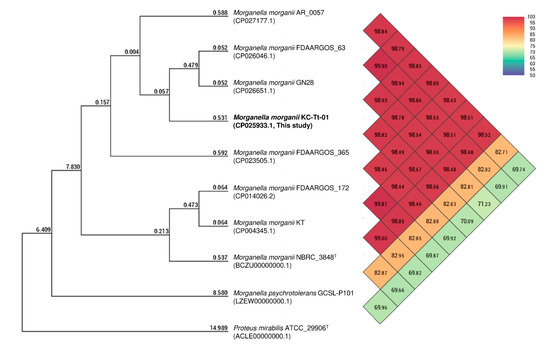
Figure 1.
Phylogenetic trees based on the orthologous average nucleotide identity (orthoANI) values calculated using available genomes of Morganella morganii, M. psychrotolerans, and Proteus mirabilis. Comparisons between two strains are given at the junction point of the diagonals departing from each strain; i.e., the OrthoANI value for M. morganii strain KC-Tt-01 (CP025933.1) and strain GN28 (CP026651.1) is 98.93%. (Two-column fitting image).
Although the origin and route of infection of strain KC-Tt-01 in a captive dolphin bred at the dolphinarium remain unclear, these findings strongly suggest that the captive dolphin-isolated M. morganii may have the potential to cause fatal zoonotic infections in humans considering the similarities in their virulence genes, along with failure in antibiotic treatment due to the prolonged cephalosporin resistance. Unfortunately, only a few studies on the AMR of bacteria in dolphinariums and resident dolphins have been conducted to date [5,6,7,8], making direct comparisons difficult. Nevertheless, the acquisition of AMR clearly poses a public health risk for humans who come in direct contact with dolphins (e.g., dolphin trainers and veterinarians). Moreover, captive dolphins harboring AMR bacteria could be a source of unintentional and unexpected spread of ARGs and AMR bacteria in wild dolphins and marine environments during their rehabilitation. Therefore, the management of antibiotic use and monitoring of the emergence of ARGs and AMR bacteria are urgently needed in captive cetaceans, at least during their rehabilitation programs, for their health and conservations. To the best of our knowledge, this is the first report of the occurrence of a third-generation cephalosporin-resistant M. morganii infection in animals including marine mammals and its associated genomic characteristics.
Supplementary Materials
The following are available online at https://www.mdpi.com/2076-2615/10/11/2052/s1, Table S1: The five prophage regions detected in the genome of Morganella morganii strain KC-Tt-01, Table S2: Potential virulence genes detected in Morganella morganii KC-Tt-01, Table S3: Potential antimicrobial resistance genes detected in Morganella morganii KC-Tt-01.
Author Contributions
Conceptualization, K.L. and J.H.K.; data curation, S.Y.P. and H.K.; formal analysis, S.Y.P. and H.K.; funding acquisition, J.H.K.; investigation, S.Y.P., S.R.L., J.E.H., K.L., and Y.C.; methodology, S.Y.P., J.E.H., and J.H.K.; project administration, J.H.K.; resources, K.L. and Y.C.; supervision, J.H.K.; writing—original draft, S.Y.P., K.L., and J.H.K.; and writing—review and editing, S.Y.P., J.E.H. and J.H.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the KRIBB Initiative programs, the National Research Foundation (NRF) of Korea (NRF-2020R1I1A2068827), and the National Institute of Fisheries Science (R2020024) of the Ministry of Oceans and Fisheries in Republic of Korea.
Conflicts of Interest
The authors declare no conflict of interest.
Ethics Statement
The animal study was reviewed and approved by ethics and welfare committee (Approved number: 2017-Animal Experiment-15) in National Institute of Fisheries Science, Ministry of Oceans and Fisheries, Republic of Korea.
References
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Bürgmann, H.; Sørum, H.; Norström, M.; Pons, M.N.; et al. Tackling antibiotic resistance: The environmental framework. Nat. Rev. Microbiol. 2015, 13, 310–317. [Google Scholar] [CrossRef]
- Dold, C. Cetacea (Whales, Dolphins, Porpoises). In Fowler’s Zoo and Wild Animal Medicine; Volume 8-E-Book; Elsevier Health Sciences: St. Louis, MO, USA, 2014; Volume 8, p. 422. [Google Scholar]
- Schaefer, A.M.; Goldstein, J.D.; Reif, J.S.; Fair, P.A.; Bossart, G.D. Antibiotic-resistant organisms cultured from Atlantic bottlenose dolphins (Tursiops truncatus) inhabiting estuarine waters of Charleston, SC and Indian River Lagoon, FL. Ecohealth 2009, 6, 33–41. [Google Scholar] [CrossRef]
- Prichula, J.; Pereira, R.I.; Wachholz, G.R.; Cardoso, L.A.; Tolfo, N.C.C.; Santestevan, N.A.; Medeiros, A.W.; Tavares, M.; Frazzon, J.; D’Azevedo, P.A.; et al. Resistance to antimicrobial agents among enterococci isolated from fecal samples of wild marine species in the southern coast of Brazil. Mar. Pollut. Bull. 2016, 105, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Wong, S. Ocean sentinels: Marine mammals and antimicrobial resistance. In Proceedings of the 42nd Interscience Conference on Antimicrobial Agents and Chemotherapy, San Diego, CA, USA, 27–30 September 2002. [Google Scholar]
- Rozanova, E.I.; Alekseev, A.Y.; Abramov, A.V.; Rassadkin, Y.N.; Shestopalov, A.M. Death of the killer whale Orsinus orca from bacterial pneumonia in 2003. Russ. J. Mar. Biol. 2007, 33, 321–323. [Google Scholar] [CrossRef]
- Gili, C.; Biancani, B.; Gulland, F.; Mazzariol, S. Meticillin-resistant Staphylococcus aureus (MRSA) associated dolphin mortality and the subsequent facility decolonisation protocol. Vet. Rec. Case Rep. 2017, e000444. [Google Scholar] [CrossRef]
- Mazzariol, S.; Corrò, M.; Tonon, E.; Centelleghe, C.; Biancani, B.; Gili, C. Death associated to methicillin resistant Staphylococcus aureus ST8 infection in two dolphins maintained under human care, Italy. Front. Immunol. 2018, 9, 2726. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhu, J.; Hu, Q.; Rao, X. Morganella morganii, a non-negligent opportunistic pathogen. Int. J. Infect Dis. 2016, 50, 10–17. [Google Scholar] [CrossRef]
- Hungerford, J.M. Scombroid poisoning: A review. Toxicon 2010, 56, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Elfadl, A.K.; Lee, S.W.; Kim, J.H.; Lee, K.L.; Ullah, H.A.; Chung, M.J.; Ghim, S.G.; Lee, E.J.; Kim, Y.D.; Kim, S.M.; et al. Fatal fibrino-hemorrhagic bronchopneumonia associated with Morganella morganii in a bottlenose dolphin: A case report. Dis. Aquat. Organ. 2017, 127, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing: 24th Informational Supplement M100-S24; CLSI: Wayne, PA, USA, 2014. [Google Scholar]
- Holt, J.G.; Kreig, N.R.; Sneath, P.H.A.; Staley, J.T.; Williams, S.T. Bergey’s Manual of Determinative Bacteriology, 9th ed.; Williams and Wilkins Co.: Baltimore, MD, USA, 1994. [Google Scholar]
- Stock, I.; Wiedemann, B. Identification and natural antibiotic susceptibility of Morganella morganii. Diagn. Microbiol. Infect. Dis. 1998, 30, 153–165. [Google Scholar] [CrossRef]
- Chen, Y.T.; Peng, H.L.; Shia, W.C.; Hsu, F.R.; Ken, C.F.; Tsao, Y.M.; Chen, C.H.; Liu, C.E.; Hsieh, M.F.; Chen, H.C.; et al. Whole-genome sequencing and identification of Morganella morganii KT pathogenicity-related genes. BMC Genom. 2012, 13, S4. [Google Scholar] [CrossRef] [PubMed]
- Tessier, F.; Arpin, C.; Allery, A.; Quentin, C. Molecular characterization of a TEM-21 β-lactamase in a clinical isolate of Morganella morganii. Antimicrob. Agents Chemother. 1998, 42, 2125–2127. [Google Scholar] [CrossRef] [PubMed]
- Barroso, H.; Freitas-Vieira, A.; Duarte, A. Molecular characterization of a ceftazidime-resistant Morganella morganii isolate producing a TEM-10 β-lactamase. Antimicrob. Agents Chemother. 1999, 43, 434–435. [Google Scholar] [CrossRef] [PubMed]
- Kuga, A.; Okamoto, R.; Inoue, M. ampR gene mutations that greatly increase class C β-lactamase activity in Enterobacter cloacae. Antimicrob. Agents Chemother. 2000, 44, 561–567. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).