First Report of aacC5-aadA7Δ4 Gene Cassette Array and Phage Tail Tape Measure Protein on Class 1 Integrons of Campylobacter Species Isolated from Animal and Human Sources in Egypt

 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Bacteriological Analysis and Molecular Identification
2.3. Antimicrobial Susceptibility Testing
2.4. PCR Amplification of Class 1 Integrons and Associated Gene Cassettes
2.5. Characterization of Gene Cassettes Arrays by DNA Sequencing
2.6. Bioinformatics and Statistical Analysis
2.7. Nucleotide Sequence Accession Numbers
3. Results
3.1. Prevalence of Campylobacter Species in Livestock Animals and Humans
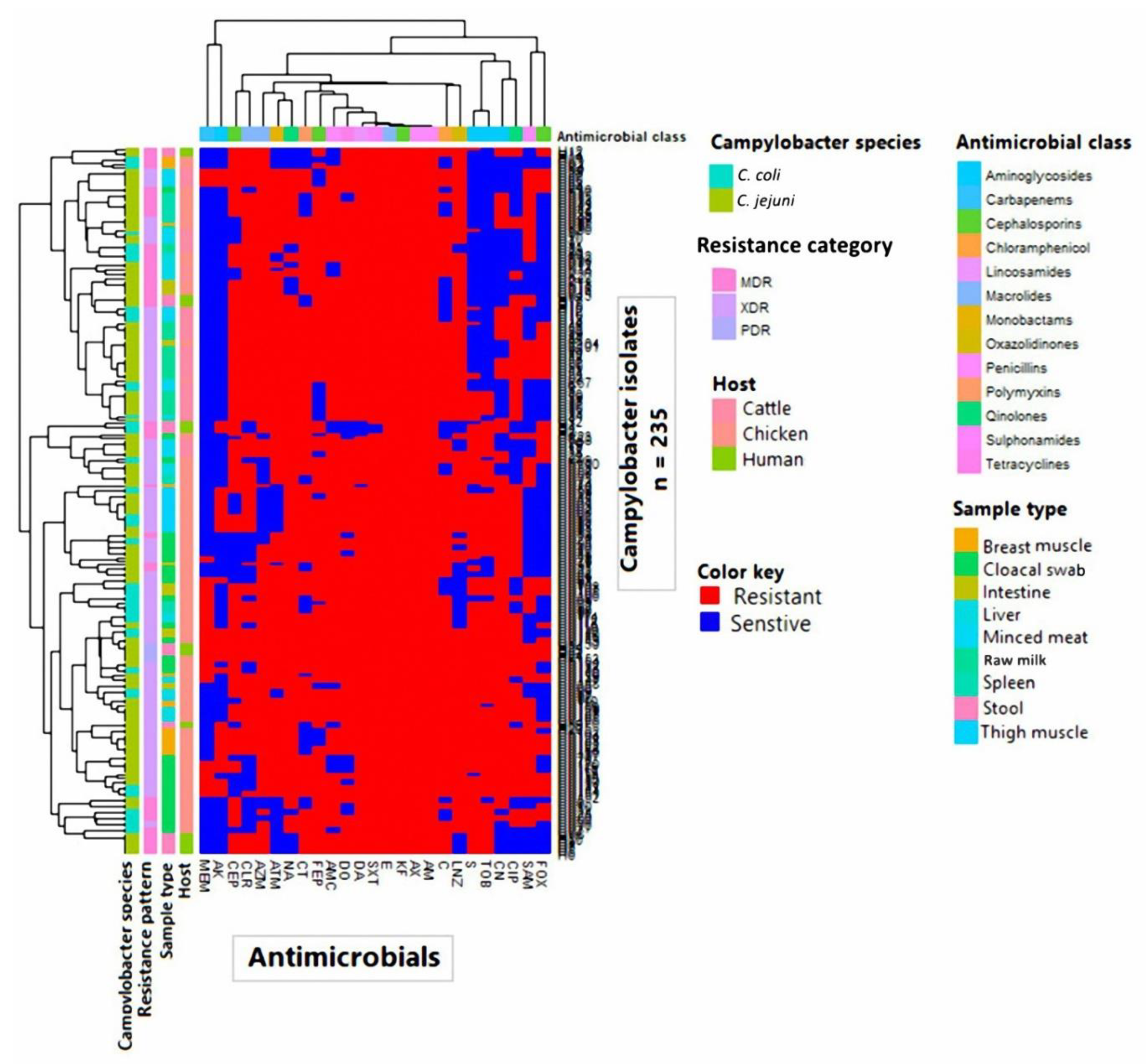
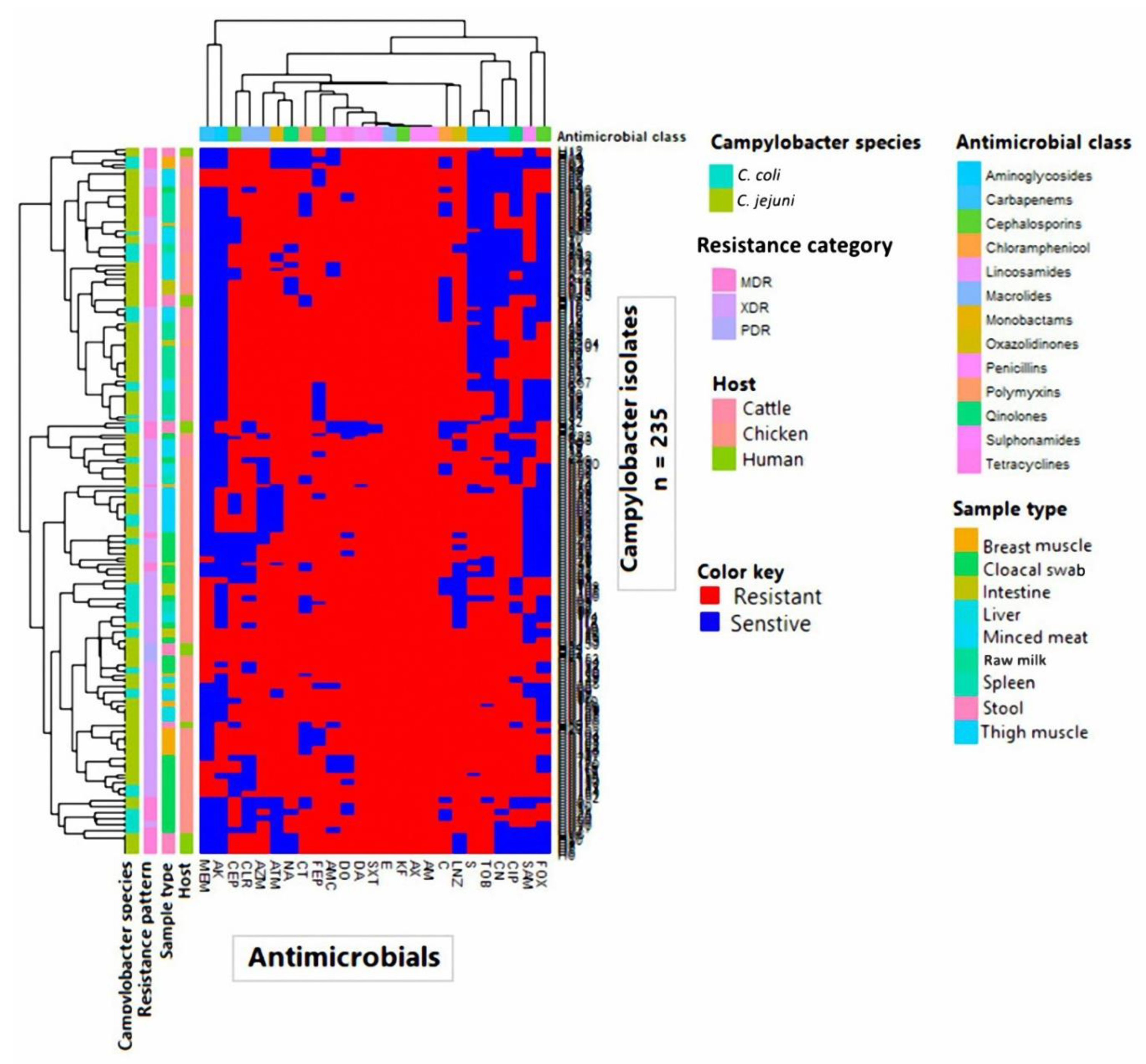
3.2. Antimicrobial Resistance Profiles
3.3. Screening for Class 1 Integrons and Characterization of Associated Gene Cassettes in Campylobacter Isolates
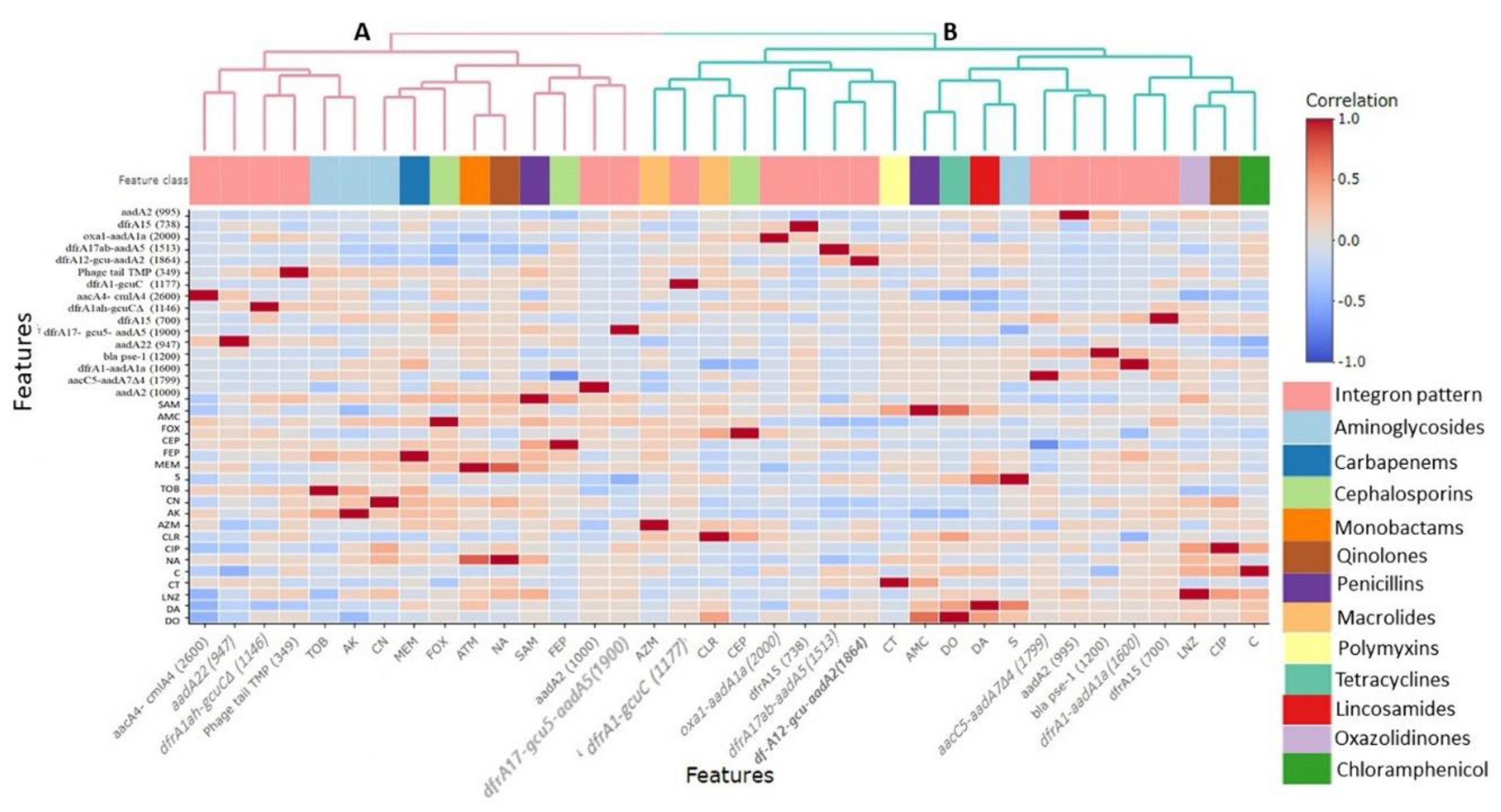
3.4. Correlation between Class 1 Integrons and Antimicrobial Resistance Phenotypes in Campylobacter Isolates
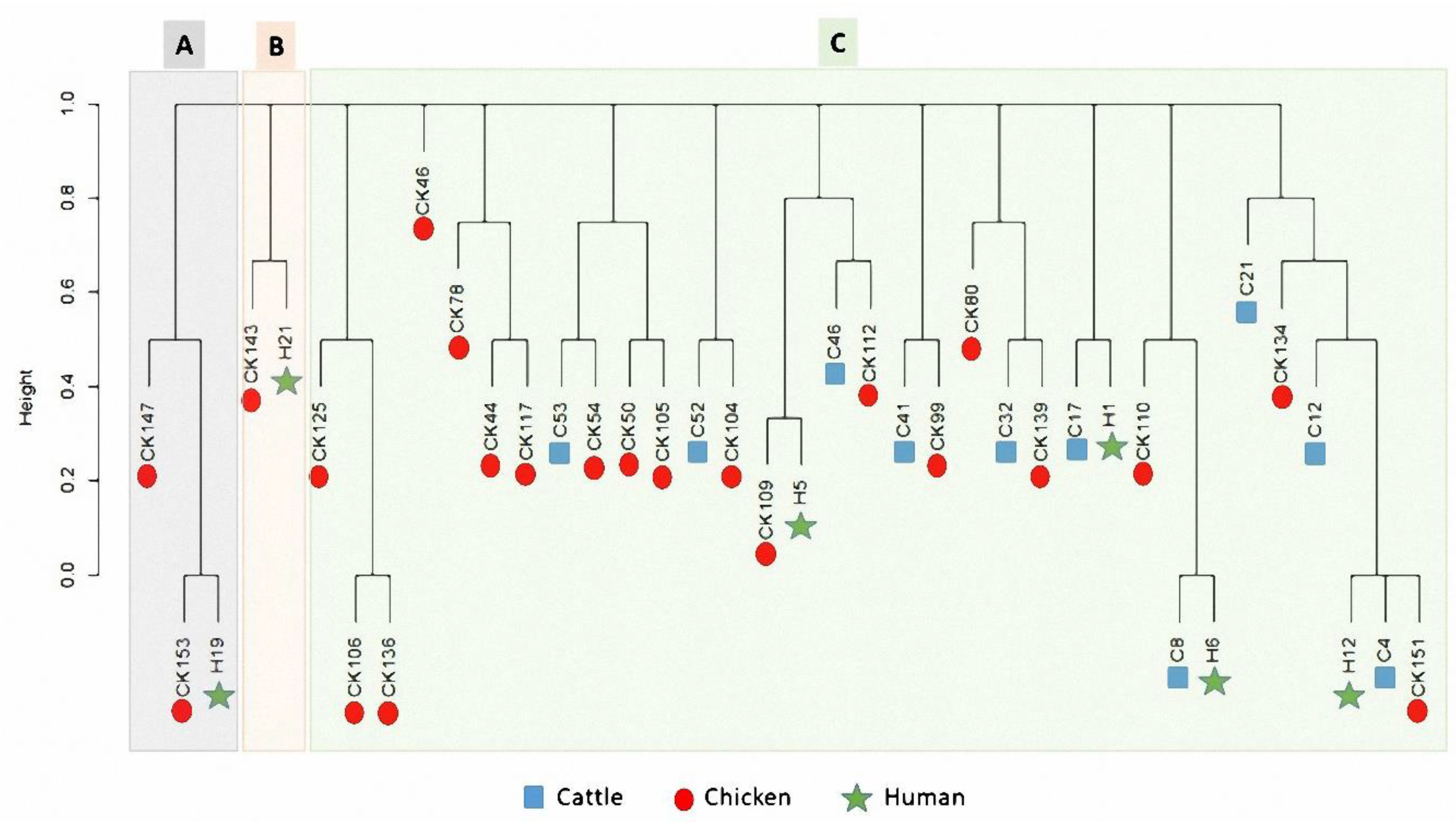
3.5. Cluster Analysis of Gene Cassette Arrays in Campylobacter Isolates from Human and Animal Populations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moore, J.E.; Corcoran, D.; Dooley, J.S.; Fanning, S.; Lucey, B.; Matsuda, M.; McDowell, D.A.; Mégraud, F.; Millar, B.C.; O’Mahony, R. Campylobacter. Vet. Res. 2005, 36, 351–382. [Google Scholar] [CrossRef] [Green Version]
- Friedman, C.R.; Hoekstra, R.M.; Samuel, M.; Marcus, R.; Bender, J.; Shiferaw, B.; Reddy, S.; Ahuja, S.D.; Helfrick, D.L.; Hardnett, F. Risk factors for sporadic Campylobacter infection in the United States: A case-control study in FoodNet sites. Clin. Infect. Dis. 2004, 38, S285–S296. [Google Scholar] [CrossRef] [Green Version]
- Neimann, J.; Engberg, J.; Mølbak, K.; Wegener, H.C. A case–control study of risk factors for sporadic Campylobacter infections in Denmark. Epidemiol. Infect. 2003, 130, 353–366. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. EU Summary Report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2013. EFSA J. 2015, 13, 4036. [Google Scholar]
- Luangtongkum, T.; Jeon, B.; Han, J.; Plummer, P.; Logue, C.M.; Zhang, Q. Antibiotic resistance in Campylobacter: Emergence, transmission and persistence. Future Microbiol. 2009, 4, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Laroche, E.; Pawlak, B.; Berthe, T.; Skurnik, D.; Petit, F. Occurrence of antibiotic resistance and class 1, 2 and 3 integrons in Escherichia coli isolated from a densely populated estuary (Seine, France). FEMS Microbiol. Ecol. 2009, 68, 118–130. [Google Scholar] [CrossRef]
- Brink, A.; Feldman, C.; Richards, G.; Moolman, J.; Senekal, M. Emergence of extensive drug resistance (XDR) among Gram-negative bacilli in South Africa looms nearer. S. Afr. Med. J. 2008, 98, 586–592. [Google Scholar]
- Park, Y.K.; Peck, K.R.; Cheong, H.S.; Chung, D.-R.; Song, J.-H.; Ko, K.S. Extreme drug resistance in Acinetobacter baumannii infections in intensive care units, South Korea. Emerg. Infect. Dis. 2009, 15, 1325. [Google Scholar] [CrossRef]
- Mazel, D. Integrons: Agents of bacterial evolution. Nat. Rev. Microbiol. 2006, 4, 608–620. [Google Scholar] [CrossRef]
- Labbate, M.; Case, R.J.; Stokes, H.W. The integron/gene cassette system: An active player in bacterial adaptation. In Horizontal Gene Transfer; Springer: Berlin/Heidelberg, Germany, 2009; pp. 103–125. [Google Scholar]
- Recchia, G.D.; Hall, R.M. Gene cassettes: A new class of mobile element. Microbiology 1995, 141, 3015–3027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.D.; Sanchez, S.; Zimmer, M.; Idris, U.; Berrang, M.E.; McDermott, P.F. Class 1 integron-associated tobramycin-gentamicin resistance in Campylobacter jejuni isolated from the broiler chicken house environment. Antimicrob. Agents Chemother. 2002, 46, 3660–3664. [Google Scholar] [CrossRef] [Green Version]
- Gibreel, A.; Sköld, O. An integron cassette carrying dfr1 with 90-bp repeat sequences located on the chromosome of trimethoprim-resistant isolates of Campylobacter jejuni. Microb. Drug Resist. 2000, 6, 91–98. [Google Scholar] [CrossRef] [PubMed]
- O’Halloran, F.; Lucey, B.; Cryan, B.; Buckley, T.; Fanning, S. Molecular characterization of class 1 integrons from Irish thermophilic Campylobacter spp. J. Antimicrob. Chemother. 2004, 53, 952–957. [Google Scholar] [CrossRef] [Green Version]
- Gibreel, A.; Sköld, O. High-level resistance to trimethoprim in clinical isolates of Campylobacter jejuni by acquisition of foreign genes (dfr1 and dfr9) expressing drug-insensitive dihydrofolate reductases. Antimicrob. Agents Chemother. 1998, 42, 3059–3064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.-C.; Tien, N.; Yang, J.-S.; Lu, C.-C.; Tsai, F.-J.; Huang, T.-J.; Wang, I.-K. Class 1 integrons and plasmid-mediated multiple resistance genes of the Campylobacter species from pediatric patient of a university hospital in Taiwan. Gut Pathog. 2017, 9, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, E.; Hong, H.; Oh, Y.; Lee, Y. First report and molecular characterization of a Campylobacter jejuni isolate with extensive drug resistance from a travel-associated human case. Antimicrob. Agents Chemother. 2015, 59, 6670–6672. [Google Scholar] [CrossRef] [Green Version]
- Vandepitte, J.; Verhaegen, J.; Engbaek, K.; Rohner, P.; Piot, P.; Heuck, C.; Heuck, C. Basic Laboratory Procedures in Clinical Bacteriology; World Health Organization: Geneva, Switzerland, 2003. [Google Scholar]
- Quinn, P.J.; Markey, B.; Carter, M.E. Campylobacter spp. In Clinical Veterinary Microbiology, 1st ed.; Wolfe Publishing: London, UK, 1994; pp. 268–272. [Google Scholar]
- Wang, G.; Clark, C.G.; Taylor, T.M.; Pucknell, C.; Barton, C.; Price, L.; Woodward, D.L.; Rodgers, F.G. Colony multiplex PCR assay for identification and differentiation of Campylobacter jejuni, C. coli, C. lari, C. upsaliensis, and C. fetus subsp. fetus. J. Clin. Microbiol. 2002, 40, 4744–4747. [Google Scholar] [CrossRef] [Green Version]
- Shin, E.; Lee, Y. Comparison of three different methods for Campylobacter isolation from porcine intestines. J. Microbiol. Biotechnol. 2009, 19, 647–650. [Google Scholar]
- Bauer, A.; Kirby, W.; Sherris, J.; Turck, M. Antibiotic susceptibility testing by a standardized single disc method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; Twenty-First Informational Supplement, CLSI document M100-S21; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2011. [Google Scholar]
- European Committee on Antimicrobial Susceptibility Testing. Breakpoint tables for interpretation of mics and zone diameters. Version 7.1, Valid from 2017-03-10. Available online: http://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_7.1_Breakpoint_Tables.pdf (accessed on 20 August 2019).
- Tambekar, D.; Dhanorkar, D.; Gulhane, S.; Khandelwal, V.; Dudhane, M. Antibacterial susceptibility of some urinary tract pathogens to commonly used antibiotics. Afr. J. Biotechnol. 2006, 5, 1562–1565. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Kashif, J.; Buriro, R.; Memon, J.; Yaqoob, M.; Soomro, J.; Dongxue, D.; Jinhu, H.; Liping, W. Detection of class 1 and 2 integrons, β-lactamase genes and molecular characterization of sulfonamide resistance in Escherichia coli isolates recovered from poultry in China. Pak. Vet. J. 2013, 33, 321–324. [Google Scholar]
- White, P.A.; McIver, C.J.; Deng, Y.-M.; Rawlinson, W.D. Characterisation of two new gene cassettes, aadA5 and dfrA17. FEMS Microbiol. Lett. 2000, 182, 265–269. [Google Scholar] [CrossRef] [Green Version]
- Sow, A.G.; Wane, A.A.; Diallo, M.H.; Boye, C.S.-B.; Aïdara-Kane, A. Genotypic characterization of antibiotic-resistant Salmonella Enteritidis isolates in Dakar, Senegal. J. Infect. Dev. Ctries. 2007, 1, 284–288. [Google Scholar]
- Kolde, R. Package ‘pheatmap’: Pretty Heat Map. 2018. Available online: https://rdrr.io/cran/pheatmap/ (accessed on 12 May 2020).
- Friendly, M. Corrgrams. Am. Stat. 2002, 56, 316–324. [Google Scholar] [CrossRef]
- Galili, T.; O’Callaghan, A.; Sidi, J.; Sievert, C. Heatmaply: An R package for creating interactive cluster heatmaps for online publishing. Bioinformatics 2018, 34, 1600–1602. [Google Scholar] [CrossRef]
- Harrell, F.E., Jr. Package ‘Hmisc’. Available online: https://cran.r-project.org/web/packages/Hmisc/Hmisc.pdf (accessed on 20 May 2020).
- Mshana, S.; Joloba, M.; Kakooza, A.; Kaddu-Mulindwa, D. Campylobacter spp among Children with acute diarrhea attending Mulago hospital in Kampala-Uganda. Afr. Health Sci. 2009, 9, 201. [Google Scholar]
- Abushahba, M.F.; Ahmed, S.O.; Ibrahim, A.A.; Mosa, H.A. Prevalence of zoonotic species of Campylobacter in broiler chicken and humans in Assiut governorate, Egypt. Approaches Poult. Dairy Vet. Sci 2018, 3, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Jouahri, M.; Asehraou, A.; Karib, H.; Hakkou, A.; Touhami, M. Prevalence and control of thermo-tolerant Campylobacter species in raw poultry meat in Morocco. MESO Prvi Hrvat. Časopis Mesu 2007, 9, 262–267. [Google Scholar]
- Leedom, J.M. Milk of nonhuman origin and infectious diseases in humans. Clin. Infect. Dis. 2006, 43, 610–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, W.; Guo, Y.C.; Ning, L. Prevalence and risk assessment of Campylobacter jejuni in chicken in China. Biomed. Environ. Sci. 2013, 26, 243–248. [Google Scholar]
- Wicker, C.; Giordano, M.; Rouger, S.; Sorin, M.-L.; Arbault, P. Campylobacter detection in food using an ELISA based method. Int. J. Med. Microbiol. 2001, 291, 1–12. [Google Scholar]
- Woldemariam, T.; Asrat, D.; Zewde, G. Prevalence of thermophilic Campylobacter species in carcasses from sheep and goats in an abattoir in Debre Zeit area, Ethiopia. Ethiop. J. Health Dev. 2009, 23. [Google Scholar] [CrossRef]
- Singh, H.; Rathore, R.S.; Singh, S.; Cheema, P.S. Comparative analysis of cultural isolation and PCR based assay for detection of Campylobacter jejuni in food and faecal samples. Braz. J. Microbiol. 2009, 42, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Hassanain, N.A. Antimicrobial resistant Campylobacter jejuni isolated from humans and animals in Egypt. Glob. Vet. 2011, 6, 195–200. [Google Scholar]
- El-Hamid, M.I.; El-Aziz, A.; Khairy, N.; Samir, M.; Remela, A.; Etab, M.; El-Naenaeey, E.-s.Y.; Bendary, M.M.; Mosbah, R.A. Genetic diversity of Campylobacter jejuni isolated from Avian and human sources in Egypt. Front. Microbial. 2019, 10, 2353. [Google Scholar] [CrossRef] [PubMed]
- Ghunaim, H.; Behnke, J.M.; Aigha, I.; Sharma, A.; Doiphode, S.H.; Deshmukh, A.; Abu-Madi, M.M. Analysis of resistance to antimicrobials and presence of virulence/stress response genes in Campylobacter isolates from patients with severe diarrhoea. PLoS ONE 2015, 10, e0119268. [Google Scholar] [CrossRef] [PubMed]
- De Vries, S.P.; Vurayai, M.; Holmes, M.; Gupta, S.; Bateman, M.; Goldfarb, D.; Maskell, D.J.; Matsheka, M.I.; Grant, A.J. Phylogenetic analyses and antimicrobial resistance profiles of Campylobacter spp. from diarrhoeal patients and chickens in Botswana. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Mdegela, R.; Nonga, H.; Ngowi, H.; Kazwala, R. Prevalence of thermophilic Campylobacter infections in humans, chickens and crows in Morogoro, Tanzania. J. Vet. Med. Ser. B 2006, 53, 116–121. [Google Scholar] [CrossRef]
- Bouhamed, R.; Bouayad, L.; Messad, S.; Zenia, S.; Naïm, M.; Hamdi, T.-M. Sources of contamination, prevalence, and antimicrobial resistance of thermophilic Campylobacter isolated from turkeys. Vet. World 2018, 11, 1074. [Google Scholar] [CrossRef]
- Marotta, F.; Garofolo, G.; di Marcantonio, L.; Di Serafino, G.; Neri, D.; Romantini, R.; Sacchini, L.; Alessiani, A.; Di Donato, G.; Nuvoloni, R. Antimicrobial resistance genotypes and phenotypes of Campylobacter jejuni isolated in Italy from humans, birds from wild and urban habitats, and poultry. PLoS ONE 2019, 14, e0223804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Giannatale, E.; Calistri, P.; Di Donato, G.; Decastelli, L.; Goffredo, E.; Adriano, D.; Mancini, M.E.; Galleggiante, A.; Neri, D.; Antoci, S. Thermotolerant Campylobacter spp. in chicken and bovine meat in Italy: Prevalence, level of contamination and molecular characterization of isolates. PLoS ONE 2019, 14, e0225957. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.A.L.-v.; Blok, H.E.M.; Donders, T.R.; Paauw, A.; Fluit, A.C.; Verhoef, J. Multidrug resistance among Enterobacteriaceae is strongly associated with the presence of integrons and is independent of species or isolate origin. J. Infec. Dis. 2003, 187, 251–259. [Google Scholar]
- Scott, A.E.; Timms, A.R.; Connerton, P.L.; Carrillo, C.L.; Radzum, K.A.; Connerton, I.F. Genome dynamics of Campylobacter jejuni in response to bacteriophage predation. PLoS Pathog. 2007, 3. [Google Scholar] [CrossRef] [Green Version]
- Scott, A.E.; Timms, A.R.; Connerton, P.L.; El-Shibiny, A.; Connerton, I.F. Bacteriophage influence Campylobacter jejuni types populating broiler chickens. Environ. Microbiol. 2007, 9, 2341–2353. [Google Scholar] [CrossRef]
- Yan, H.; Li, L.; Zong, M.; Alam, M.J.; Shinoda, S.; Shi, L. Occurrence and characteristics of Class 1 and 2 integrons in clinical bacterial isolates from patients in south China. J. Health Sci. 2010, 56, 442–450. [Google Scholar] [CrossRef] [Green Version]
- Lorestani, R.C.; Akya, A.; Elahi, A.; Hamzavi, Y. Gene cassettes of class 1 integron-associated with antimicrobial resistance in isolates of Citrobacter spp. with multidrug resistance. Iran. J. Microbiol. 2018, 10, 22. [Google Scholar]
- Zhao, X.; Yang, J.; Zhang, B.; Sun, S.; Chang, W. Characterization of integrons and resistance genes in Salmonella isolates from farm animals in Shandong Province, China. Front. Microbiol. 2017, 8, 1300. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Source (No.) | Sample Type (No.) | Overall Prevalence of Campylobacter Isolates No. (%) | Campylobacter Species No. (%) | p Value | |
|---|---|---|---|---|---|
| C. jejuni | C. coli | ||||
| Chickens and chicken products (265) | Cloacal swab (70) | 60 (85.71) | 40 (57.14) | 20 (28.57) | 0.001 |
| Breast muscle (25) | 18 (72.00) | 14 (56.00) | 4 (16.00) | 0.003 | |
| Thigh chicken muscle (25) | 16 (64.00) | 10 (40.00) | 6 (24.00) | 0.225 | |
| Liver (25) | 25 (100.00) | 16 (64.00) | 9 (36.00) | 0.048 | |
| Spleen (20) | 18 (90.00) | 14 (70.00) | 4 (20.00) | 0.001 | |
| Intestine (20) | 17 (85.00) | 10 (50.00) | 7 (35.00) | 0.337 | |
| Chicken beef (40) | 0 (0.00) | 0 (0.00) | 0 (0.00) | NE | |
| Chicken luncheon (40) | 0 (0.00) | 0 (0.00) | 0 (0.00) | NE | |
| Meat and meat products (160) | Minced meat (35) | 28 (80.00) | 18 (51.43) | 10 (28.57) | 0.05 |
| Smoked meat (45) | 0 (0.00) | 0 (0.00) | 0 (0.00) | NE | |
| Meat beef (40) | 0 (0.00) | 0 (0.00) | 0 (0.00) | NE | |
| Meat luncheon (40) | 0 (0.00) | 0 (0.00) | 0 (0.00) | NE | |
| Dairy products (95) | Raw milk (35) | 29 (82.86) | 26 (74.29) | 3 (8.57) | <0.0001 |
| Canned milk (20) | 0 (0.00) | 0 (0.00) | 0 (0.00) | NE | |
| Canned cheese (20) | 0 (0.00) | 0 (0.00) | 0 (0.00) | NE | |
| Raw cheese (20) | 0 (0.00) | 0 (0.00) | 0 (0.00) | NE | |
| Human (30) | Stool (30) | 24 (80.00) | 20 (66.67) | 4 (13.33) | <0.0001 |
| Total | 550 | 235 (42.73) | 168 (71.49) | 67 (28.51) | <0.0001 |
| AMA | Number of C. jejuni Isolates | Number of C. coli Isolates | p Value | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chickens (n = 104) | Raw Milk (26) | Minced Meat (18) | Human Stool (20) | Total (168) | Chickens (n = 50) | Raw Milk (3) | Minced Meat (10) | Human Stool (4) | Total (67) | |||||||
| Muscle (24) | Internal Organs (40) | Cloacal Swabs (40) | Muscle (10) | Internal Organs (20) | Cloacal Swabs (20) | Various Hosts | Campylobacter Species | |||||||||
| AX | 24 | 40 | 40 | 26 | 18 | 20 | 168 | 10 | 20 | 20 | 3 | 10 | 4 | 67 | NA | NA |
| (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | |||
| AM | 24 | 40 | 40 | 26 | 18 | 20 | 168 | 10 | 20 | 20 | 3 | 10 | 4 | 67 | NA | NA |
| (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | |||
| SAM | 15 | 27 | 20 | 11 | 10 | 10 | 93 | 0.00 | 11 | 16 | 3 | 1 | 0.00 | 31 | 0.1022 | 0.2 |
| (62.50) | (67.50) | (50.00) | (42.31) | (55.55) | (50.00) | (55.36) | (0.00) | (55.00) | (80.00) | (100.00) | (10.00) | (0.00) | (46.27) | |||
| AMC | 24 | 34 | 34 | 26 | 17 | 15 | 150 | 7 | 20 | 18 | 3 | 10 | 0.00 | 58 | <0.0001 | 0.55 |
| (100.00) | (85.00) | (85.00) | (100.00) | (94.44) | (75.00) | (89.29) | (70.00) | (100.00) | (90.00) | (100.00) | (100.00) | (0.00) | (86.57) | |||
| KF | 24 | 40 | 40 | 26 | 18 | 20 | 168 | 10 | 20 | 20 | 3 | 10 | 4 | 67 | NA | NA |
| (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | |||
| FOX | 8 | 4 | 18 | 17 | 12 | 9 | 68 | 0.00 | 4 | 10 | 0.00 | 0.00 | 4 | 18 | 0.002 | 0.05 |
| (33.33) | (10.00) | (45.00) | (65.38) | (66.67) | (45.00) | (40.48) | (0.00) | (20.00) | (50.00) | (0.00) | (0.00) | (100.00) | (26.87) | |||
| CEP | 14 | 34 | 19 | 26 | 12 | 18 | 123 | 10 | 20 | 12 | 3 | 9 | 4 | 58 | 0.0068 | 0.02 |
| (58.33) | (85.00) | (47.50) | (100.00) | (66.67) | (90.00) | (73.21) | (100.00) | (100.00) | (60.00) | (100.00) | (90.00) | (100.00) | (86.57) | |||
| FEP | 16 | 38 | 39 | 17 | 10 | 20 | 140 | 8 | 19 | 20 | 3 | 6 | 4 | 60 | <0.0001 | 0.2 |
| (66.67) | (95.00) | (97.50) | (65.38) | (55.56) | (100.00) | (83.33) | (80.00) | (95.00) | (100.00) | (100.00) | (60.00) | (100.00) | (89.55) | |||
| MEM | 1 | 2 | 20 | 0.00 | 6 | 4 | 33 | 0.00 | 14 | 8 | 0.00 | 0.00 | 0.00 | 22 | 0.0124 | 0.03 |
| (4.17) | (5.00) | (50.00) | (0.00) | (33.33) | (20.00) | (19.64) | (0.00) | (70.00) | (40.00) | (0.00) | (0.00) | (0.00) | (32.84) | |||
| ATM | 14 | 39 | 36 | 26 | 17 | 10 | 142 | 0.00 | 14 | 12 | 3 | 10 | 4 | 43 | <0.0001 | 0.0006 |
| (58.33) | (97.50) | (90.00) | (100.00) | (94.44) | (50.00) | (84.52) | (0.00) | (70.00) | (60.00) | (100.00) | (100.00) | (100.00) | (64.18) | |||
| S | 22 | 16 | 34 | 12 | 7 | 14 | 105 | 4 | 17 | 18 | 0.00 | 2 | 3 | 44 | <0.0001 | 0.6 |
| (91.67) | (40.00) | (85.00) | (46.15) | (38.89) | (70.00) | (62.50) | (40.00) | (85.00) | (90.00) | (0.00) | (20.00) | (75.00) | (65.67) | |||
| TOB | 22 | 16 | 30 | 8 | 6 | 13 | 95 | 4 | 17 | 17 | 0.00 | 0.00 | 4 | 42 | <0.0001 | 0.38 |
| (91.67) | (40.00) | (75.00) | (30.77) | (33.33) | (65.00) | (56.55) | (40.00) | (85.00) | (85.00) | (0.00) | (0.00) | (100.00) | (62.69) | |||
| CN | 24 | 24 | 36 | 14 | 2 | 9 | 109 | 6 | 17 | 14 | 0.00 | 5 | 4 | 46 | <0.0001 | 0.58 |
| (100.00) | (60.00) | (90.00) | (53.85) | (11.11) | (45.00) | (64.88) | (60.00) | (85.00) | (70.00) | (0.00) | (50.00) | (100.00) | (68.66) | |||
| AK | 18 | 0.00 | 14 | 0.00 | 6 | 6 | 44 | 6 | 3 | 6 | 0.00 | 00.00 | 4 | 19 | 0.0032 | 0.73 |
| (75.00) | (0.00) | (35.00) | (0.00) | (33.33) | (30.00) | (26.19) | (60.00) | (15.00) | (30.00) | (0.00) | (0.00) | (100.00) | (28.36) | |||
| E | 24 | 40 | 40 | 26 | 18 | 20 | 168 | 10 | 20 | 20 | 3 | 10 | 4 | 67 | NA | NA |
| (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | |||
| AZM | 15 | 33 | 32 | 26 | 18 | 20 | 144 | 4 | 20 | 14 | 3 | 10 | 4 | 55 | <0.0001 | 0.48 |
| (62.50) | (82.50) | (80.00) | (100.00) | (100.00) | (100.00) | (85.71) | (40.00) | (100.00) | (70.00) | (100.00) | (100.00) | (100.00) | (82.09) | |||
| CLR | 24 | 40 | 6 | 26 | 18 | 20 | 134 | 10 | 20 | 2 | 3 | 10 | 0.00 | 45 | <0.0001 | 0.04 |
| (100.00) | (100.00) | (15.00) | (100.00) | (100.00) | (100.00) | (79.76) | (100.00) | (100.00) | (10.00) | (100.00) | (100.00) | (0.00) | (67.16) | |||
| CIP | 24 | 16 | 32 | 26 | 1 | 11 | 110 | 6 | 9 | 16 | 0.00 | 9 | 0.00 | 40 | <0.0001 | 0.4 |
| (100.00) | (40.00) | (80.00) | (100.00) | (5.56) | (55.00) | (65.48) | (60.00) | (45.00) | (80.00) | (0.00) | (90.00) | (0.00) | (59.70) | |||
| NA | 24 | 33 | 40 | 26 | 17 | 10 | 150 | 6 | 20 | 18 | 0.00 | 10 | 4 | 58 | <0.0001 | 0.55 |
| (100.00) | (82.50) | (100.00) | (100.00) | (94.44) | (50.00) | (89.29) | (60.00) | (100.00) | (90.00) | (0.00) | (100.00) | (100.00) | (86.57) | |||
| SXT | 24 | 40 | 40 | 26 | 18 | 20 | 168 | 10 | 20 | 20 | 3 | 10 | 4 | 67 | <0.0001 | 0.005 |
| (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | |||
| C | 24 | 26 | 32 | 26 | 17 | 16 | 141 | 6 | 16 | 16 | 3 | 5 | 0.00 | 46 | 0.0461 | 0.0088 |
| (100.00) | (65.00) | (80.00) | (100.00) | (94.44) | (80.00) | (83.93) | (60.00) | (80.00) | (80.00) | (100.00) | (50.00) | (0.00) | (68.66) | |||
| CT | 14 | 28 | 34 | 26 | 18 | 11 | 131 | 6 | 16 | 18 | 3 | 10 | 4 | 57 | <0.0001 | 0.21 |
| (58.33) | (70.00) | (85.00) | (100.00) | (100.00) | (55.00) | (77.98) | (60.00) | (80.00) | (90.00) | (100.00) | (100.00) | (100.00) | (85.07) | |||
| LNZ | 24 | 39 | 28 | 26 | 14 | 13 | 144 | 10 | 9 | 18 | 0.00 | 10 | 0.00 | 47 | 0.0012 | 0.005 |
| (100.00) | (97.50) | (70.00) | (100.00) | (77.78) | (65.00) | (85.71) | (100.00) | (45.00) | (90.00) | (0.00) | (100.00) | (0.00) | (70.15) | |||
| DA | 23 | 40 | 39 | 26 | 18 | 20 | 166 | 10 | 20 | 20 | 3 | 10 | 0.00 | 63 | <0.0001 | 0.03 |
| (95.83) | (100.00) | (97.50) | (100.00) | (100.00) | (100.00) | (98.81) | (100.00) | (100.00) | (100.00) | (100.00) | (100.00) | (0.00) | (94.03) | |||
| DO | 24 | 37 | 24 | 26 | 18 | 20 | 149 | 10 | 20 | 18 | 3 | 10 | 0.00 | 61 | 0.0103 | 0.59 |
| (100.00) | (92.50) | (60.00) | (100.00) | (100.00) | (100.00) | (88.69) | (100.00) | (100.00) | (90.00) | (100.00) | (100.00) | (0.00) | (91.04) | |||
| Resistance Category | Resistance to Antimicrobial Class (n = 14) | Resistance to Antimicrobial Agent (n = 25) | No. of Resistant Campylobacter Isolates (Source) | |
|---|---|---|---|---|
| C. jejuni (n = 168) | C. coli (n = 67) | |||
| MDR (n = 67) | 7 | 11 | 0 | 2 (chicken muscle) |
| 12 | 0 | 1 (chicken muscle) | ||
| 14 | 0 | 1 (human stool) | ||
| 15 | 0 | 3 (human stool) | ||
| 8 | 13 | 0 | 1 (chicken muscle) | |
| 10 | 13 | 2 (cloacal swab) | 0 | |
| 14 | 1 (minced meat) | 0 | ||
| 15 | 8 (chicken organ) | 0 | ||
| 16 | 2 (cloacal swab), 9 (human stool) | 3 (raw milk) | ||
| 17 | 5 (human stool), 1 (cloacal swab) | 2 (cloacal swab) | ||
| 11 | 15 | 1 (minced meat) | 2 (cloacal swab) | |
| 16 | 4 (cloacal swab), 2 (chicken organ) | 0 | ||
| 17 | 8 (chicken organ), 2 (cloacal swab) | 3 (chicken organ) | ||
| 18 | 1 (cloacal swab) | 0 | ||
| 19 | 1 (chicken muscle) | 2 (cloacal swab) | ||
| XDR (n = 162) | 12 | 16 | 2 (cloacal swab) | 0 |
| 17 | 3 (minced meat) | 4 (minced meat) | ||
| 18 | 2 (chicken muscle), 5 (chicken organ), 4 (cloacal swab) | 4 (minced meat), 2 (chicken muscle), 2 (cloacal swab) | ||
| 19 | 5 (chicken muscle), 5 (chicken organ), 2 (cloacal swab), 4 (minced meat) | 1 (minced meat), 1 (chicken organ), 2 (cloacal swab) | ||
| 20 | 5 (raw milk), 2 (chicken muscle), 2 (cloacal swab), 2 (chicken organ), 2 (human stool) | 6 (chicken organ), 4 (chicken muscle) | ||
| 21 | 2 (chicken muscle) | 4 (chicken organ) | ||
| 22 | 4 (chicken muscle) | 0 | ||
| 23 | 3 (chicken muscle) | 0 | ||
| 13 | 18 | 1 (minced meat) | 1 (minced meat), 2 (cloacal swab) | |
| 19 | 9 (raw milk), 1 (chicken muscle), 3 (chicken organ) | 1 (cloacal swab) | ||
| 20 | 2 (minced meat), 5 (raw milk) | 1 (cloacal swab) | ||
| 21 | 6 (raw milk), 3 (chicken muscle), 6 (cloacal swab) | 3 (chicken organ) | ||
| 22 | 1 (raw milk), 5 (chicken organ), 2 (cloacal swab) | 0 | ||
| 23 | 1 (chicken muscle) | 0 | ||
| 14 | 18 | 1 (cloacal swab) | 0 | |
| 19 | 1 (cloacal swab) | 0 | ||
| 20 | 6 (minced meat) | 0 | ||
| 21 | 1 (cloacal swab) | 0 | ||
| 22 | 3 (cloacal swab) | 0 | ||
| 23 | 2 (cloacal organ) | 3 (chicken organ), 4 (cloacal swab) | ||
| 24 | 2 (cloacal swab) | 2 (cloacal swab) | ||
| PDR (n = 6) | 14 | 25 | 4 (human stool), 2 (cloacal swab) | 0 |
| Isolate No. | Code No. | Campylobacter Species | Source | Antimicrobial Resistance Pattern | Resistance to Antimicrobial Agents | Resistance to Antimicrobial Classes | Antimicrobial Resistance Type | intI1/Class 1 Integron | Resistance Gene(s) in Class 1 Integron (Size in bp) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | CK105 | C. coli | Chicken intestine | AX, AM, SAM, AMC, KF, FOX, CEP, FEP, MEM, ATM, S, TOB, CN, E, AZM, CLR, NA, SXT, CT, DA, DO | 21 | 12 | XDR | +/+ | aadA2 (995) |
| 2 | CK134 | C. jejuni | Chicken cloacal swab | AX, AM, SAM, AMC, KF, FOX, FEP, MEM, ATM, TOB, CN, E, AZM, CIP, NA, SXT, C, CT, LNZ, DA, DO | 21 | 14 | XDR | +/+ | dfrA15 (738) |
| 3 | CK110 | C. jejuni | Chicken thigh muscle | AX, AM, SAM, AMC, KF, FEP, ATM, S, TOB, CN, E, AZM, CLR, CIP, NA, SXT, C, CT, LNZ, DA, DO | 21 | 13 | XDR | +/+ | dfrA15 (738); oxa1-aadA1a (2000) |
| 4 | H12 | C. jejuni | Human stool | AX, AM, KF, FOX, CEP, FEP, S, CN, E, AZM, CLR, CIP, SXT, C, LNZ, DA, DO | 17 | 10 | MDR | +/+ | dfrA17ab-aadA5 (1513); dfrA12-gcu-aadA2 (1864) |
| 5 | H19 | C. jejuni | Human stool | AX, AM, SAM, KF, FOX, FEP, ATM, S, TOB, CN, AK, E, AZM, CLR, NA, SXT, C, LNZ, DA, DO | 20 | 12 | XDR | +/+ | dfrA15 (738); dfrA17ab-aadA5 (1513); oxa1-aadA1a (2000) |
| 6 | CK139 | C. jejuni | Chicken breast muscle | AX,AM, SAM, AMC, KF, FOX, CEP, FEP, MEM, ATM, S, CN, TOB, E, AZM, CLR, CIP, NA, SXT, C, LNZ, DA, DO | 23 | 13 | XDR | +/+ | dfrA12-gcu-aadA2 (1864) |
| 7 | CK104 | C. jejuni | Chicken intestine | AX, AM, SAM, AMC, KF, FOX, CEP, FEP, ATM, E, AZM, CLR, CIP, NA, SXT, C, CT, LNZ, DA, DO | 20 | 12 | XDR | +/+ | Phage tail TMP (349) |
| 8 | CK147 | C. jejuni | Chicken Liver | AX, AM, AMC, SAM, KF, FEP, CEP, FOX, MEM, ATM, S, TOB, CN, E, AZM, CLR, NA, SXT, C, CT, LNZ, DA, DO | 23 | 14 | XDR | +/+ | dfrA1-gcuC (1177) |
| 9 | C41 | C. jejuni | Raw milk | AX, AM, SAM, AMC, KF, FOX, CEP, FEP, ATM, CN, E, AZM, CLR, CIP, NA, SXT, C, CT, LNZ, DA, DO | 21 | 13 | XDR | +/+ | aacA4-cmlA4 (2600) |
| 10 | CK143 | C. coli | Chicken cloacal swab | AX, AM, SAM, AMC, KF, FOX, FEP, MEM, ATM, S, TOB, CN, AK, E, AZM, CIP, NA, SXT, C, CT, LNZ, DA, DO | 23 | 14 | XDR | +/+ | dfrA15 (738); dfrA17ab-aadA5 (1513) |
| 11 | H1 | C. coli | Human stool | AX, AM, KF, FOX, CEP, FEP, ATM, TOB, CN, AK, E, AZM, NA, SXT, CT | 15 | 8 | MDR | +/+ | dfrA15 (700); dfrA1ah-gcuCΔ (1146) |
| 12 | C52 | C. jejuni | Raw milk | AX, AM, SAM, AMC, KF, FOX, CEP, FEP, ATM, S, CN, E, AZM, CLR, CIP, NA, SXT, C, CT, LNZ, DA, DO | 22 | 13 | XDR | +/+ | dfrA17ab-aadA5 (1513); dfrA12-gcu-aadA2 (1864) |
| 13 | C8 | C. jejuni | Minced meat | AX, AM, AMC, SAM, KF, FEP, ATM, S, E, AZM, CLR, NA, SXT, C, CT, LNZ, DA, DO | 18 | 12 | XDR | +/+ | dfrA17-gcu5-aadA5 (1900) |
| 14 | H5 | C. jejuni | Human stool | AX, AM, AMC, KF, CEP, FEP, S, TOB, E, AZM, CLR, SXT, C, CT, DA, DO | 15 | 10 | MDR | +/+ | Phage tail TMP (349) |
| 15 | CK54 | C. coli | Chicken cloacal swab | AX, AM, AMC, KF, FEP, ATM, S, TOB, CN, E, CIP, NA, SXT, C, CT, LNZ, DA, DO | 18 | 13 | XDR | +/+ | aadA22 (947); dfrA17ab-aadA5 (1513) |
| 16 | CK80 | C. jejuni | Chicken breast muscle | AX, AM, SAM, AMC, KF, CEP, FEP, TOB, CN, E, AZM, CLR, CIP, NA, SXT, C, CT, LNZ, DO | 19 | 11 | MDR | -/- | ND |
| 17 | CK125 | C. jejuni | Chicken Liver | AX, AM, SAM, AMC, KF, FEP, CEP, ATM, S, TOB, CN, E, AZM, CLR, CIP, NA, SXT, C, CT, LNZ, DA, DO | 22 | 13 | XDR | +/+ | dfrA1-gcuC (1177); dfrA12-gcu-aadA2 (1864) |
| 18 | C12 | C. coli | Minced meat | AX, AM, AMC, KF, CEP, FEP, ATM, S, CN, E, AZM, CLR, CIP, NA, SXT, CT, LNZ, DA, DO | 19 | 12 | XDR | +/+ | dfrA15 (700); dfrA1-gcuC (1177); dfrA17ab-aadA5 (1513) |
| 19 | CK78 | C. jejuni | Chicken cloacal swab | AX, AM, AMC, KF, FEP, MEM, ATM, S, CN, E, AZM, CIP, NA, SXT, C, CT, LNZ, DA, DO | 19 | 14 | XDR | +/+ | dfrA15 (738); aadA22 (947); dfrA1ah-gcuCΔ (1146) |
| 20 | C46 | C. jejuni | Raw milk | AX, AM, AMC, KF, FOX, CEP, FEP, ATM, S, CN, E, AZM, CLR, CIP, NA, SXT, C, CT, LNZ, DA, DO | 21 | 13 | XDR | +/+ | aadA2 (995); bla pse-1 (1200) |
| 21 | CK109 | C. jejuni | Chicken breast muscle | AX, AM, SAM, AMC, KF, CEP, FEP, ATM, S, CN, E, AZM, CLR, CIP, NA, SXT, C, CT, LNZ, DA, DO | 21 | 13 | XDR | +/+ | dfrA15 (700); dfrA1-aadA1a (1600) |
| 22 | CK153 | C. jejuni | Chicken cloacal swab | AX, AM, SAM, AMC, KF, FOX, CEP, FEP, MEM, ATM, S, TOB, CN, AK, E, AZM, CLR, CIP, NA, SXT, C, CT, LNZ, DA, DO | 25 | 14 | PDR | +/+ | dfrA15 (738); dfrA17-gcu5-aadA5 (1900) |
| 23 | CK50 | C. jejuni | Chicken liver | AX, AM, SAM, AMC, KF, FEP, CEP, ATM, S, TOB, E, CLR, NA, SXT, CT, LNZ, DA, DO | 18 | 12 | XDR | +/+ | dfrA15 (738); bla pse-1 (1200); dfrA1-aadA1a (1600) |
| 24 | CK117 | C. jejuni | Chicken cloacal swab | AX, AM, SAM, KF, CEP, FEP, MEM, ATM, S, TOB, CN, AK, E, AZM, CIP, NA, SXT, C, CT, LNZ, DA | 21 | 13 | XDR | +/+ | aadA2 (995); dfrA17-gcu5-aadA5 (1900) |
| 25 | H21 | C. jejuni | Human stool | AX, AM, SAM, AMC, KF, FOX, CEP, FEP, MEM, ATM, S, TOB, CN, AK, E, AZM, CLR, CIP, NA, SXT, C, CT, LNZ, DA, DO | 25 | 14 | PDR | +/+ | aadA22 (947); aacA4-cmlA4 (2600) |
| 26 | C32 | C. jejuni | Raw milk | AX, AM, AMC, KF, CEP, ATM, S, CN, E, AZM, CLR, CIP, NA, SXT, C, CT, LNZ, DA, DO | 19 | 13 | XDR | +/+ | dfrA15 (700); aadA2 (995); bla pse-1 (1200); aacC5-aadA7Δ4 (1799) |
| 27 | CK44 | C. jejuni | Chicken thigh muscle | AX, AM, AMC, KF, S, TOB, CN, AK, E, CLR, CIP, NA, SXT, C, CT, LNZ, DA, DO | 18 | 12 | XDR | +/+ | aadA2 (1000) |
| 28 | H6 | C. jejuni | Human stool | AX, AM, AMC, KF,CEP, FEP, S, TOB, E, AZM, CLR, SXT, C, CT, DA, DO | 16 | 10 | MDR | +/+ | aadA2 (995) |
| 29 | CK112 | C. coli | Chicken Liver | AX, AM, AMC, KF, FEP, CEP, MEM, ATM, S, TOB, CN, E, AZM, CLR, CIP, NA, SXT, C, CT, DA, DO | 21 | 13 | XDR | +/+ | dfrA1-gcuC (1177) |
| 30 | C17 | C. jejuni | Minced meat | AX, AM, AMC, KF, FOX, CEP, FEP, ATM, S, TOB, E, AZM, CLR, NA, SXT, C, CT, DA, DO | 19 | 12 | XDR | +/+ | dfrA15 (738); aadA2 (1000) |
| 31 | CK106 | C. coli | Chicken intestine | AX, AM, SAM, AMC, KF, FOX, CEP, FEP, MEM, ATM, S, TOB, CN, E, AZM, CLR, NA, SXT, CT, DA, DO | 21 | 12 | XDR | +/+ | Phage tail TMP (349); aadA22 (947) |
| 32 | CK99 | C. coli | Chicken cloacal swab | AX, AM, SAM, AMC, KF, FOX, CEP, FEP, ATM, S, CN, E, NA, SXT, C, CT, LNZ, DA, DO | 19 | 13 | XDR | +/+ | dfrA15 (738); aacC5-aadA7Δ4 (1799) |
| 33 | C53 | C. jejuni | Raw milk | AX, AM, SAM, AMC, KF, FOX, CEP, FEP, ATM, S, CN, E, AZM, CLR, CIP, NA, SXT, C, CT, LNZ, DA, DO | 22 | 13 | XDR | +/+ | aadA2 (995) |
| 34 | C4 | C. coli | Minced meat | AX, AM, AMC, KF, CEP, ATM, S, E, AZM, CLR, CIP, NA, SXT, C, CT, LNZ, DA, DO | 18 | 13 | XDR | +/+ | aadA22 (947) |
| 35 | C21 | C. jejuni | Minced meat | AX, AM, AMC, KF, FOX, CEP, ATM, S, TOB, CN, E, AZM, CLR, NA, SXT, C, CT, LNZ, DA, DO | 20 | 13 | XDR | +/+ | oxa1-aadA1a (2000) |
| 36 | CK46 | C. jejuni | Chicken cloacal swab | AX, AM, AMC, KF, FEP, ATM, S, TOB, CN, E, AZM, CIP, NA, SXT, C, CT, DA, DO | 18 | 12 | XDR | +/+ | Phage tail TMP (349); dfrA1ah-gcuCΔ (1146); oxa1-aadA1a (2000) |
| 37 | CK151 | C. coli | Chicken cloacal swab | AX, AM, SAM, AMC, KF, FOX, FEP, MEM, ATM, S, TOB, CN, AK, E, AZM, CLR, CIP, NA, SXT, C, CT, LNZ, DA, DO | 24 | 14 | XDR | +/+ | dfrA15 (738); aadA22 (947); bla pse-1 (1200) |
| 38 | CK136 | C. jejuni | Chicken breast muscle | AX, AM, SAM, AMC, KF, FOX, CEP, FEP, ATM, S, CN, TOB, AK, E, AZM, CLR, CIP, NA, SXT, C, LNZ, DA, DO | 23 | 12 | XDR | +/+ | dfrA17ab-aadA5 (1513) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd El-Aziz, N.K.; Ammar, A.M.; Hamdy, M.M.; Gobouri, A.A.; Azab, E.; Sewid, A.H. First Report of aacC5-aadA7Δ4 Gene Cassette Array and Phage Tail Tape Measure Protein on Class 1 Integrons of Campylobacter Species Isolated from Animal and Human Sources in Egypt. Animals 2020, 10, 2067. https://doi.org/10.3390/ani10112067
Abd El-Aziz NK, Ammar AM, Hamdy MM, Gobouri AA, Azab E, Sewid AH. First Report of aacC5-aadA7Δ4 Gene Cassette Array and Phage Tail Tape Measure Protein on Class 1 Integrons of Campylobacter Species Isolated from Animal and Human Sources in Egypt. Animals. 2020; 10(11):2067. https://doi.org/10.3390/ani10112067
Chicago/Turabian StyleAbd El-Aziz, Norhan K., Ahmed M. Ammar, Mona M. Hamdy, Adil A. Gobouri, Ehab Azab, and Alaa H. Sewid. 2020. "First Report of aacC5-aadA7Δ4 Gene Cassette Array and Phage Tail Tape Measure Protein on Class 1 Integrons of Campylobacter Species Isolated from Animal and Human Sources in Egypt" Animals 10, no. 11: 2067. https://doi.org/10.3390/ani10112067
APA StyleAbd El-Aziz, N. K., Ammar, A. M., Hamdy, M. M., Gobouri, A. A., Azab, E., & Sewid, A. H. (2020). First Report of aacC5-aadA7Δ4 Gene Cassette Array and Phage Tail Tape Measure Protein on Class 1 Integrons of Campylobacter Species Isolated from Animal and Human Sources in Egypt. Animals, 10(11), 2067. https://doi.org/10.3390/ani10112067